Nitrate Transport Rate in the Xylem of Tomato Plants Grafted onto a Vigorous Rootstock

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Measurements
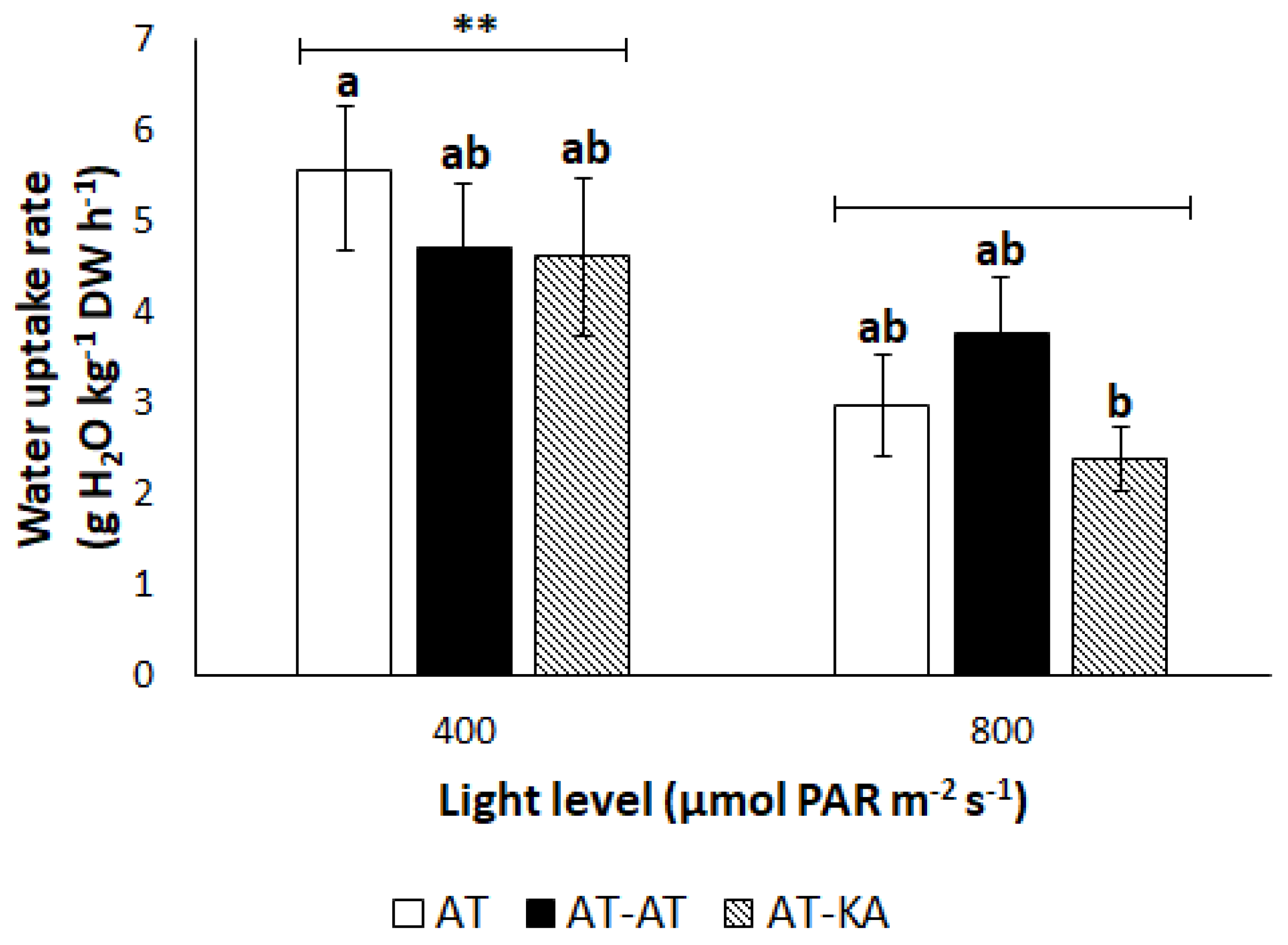
2.2.1. Plant Water Uptake
2.2.2. Plant Growth Rate and RGR Calculations
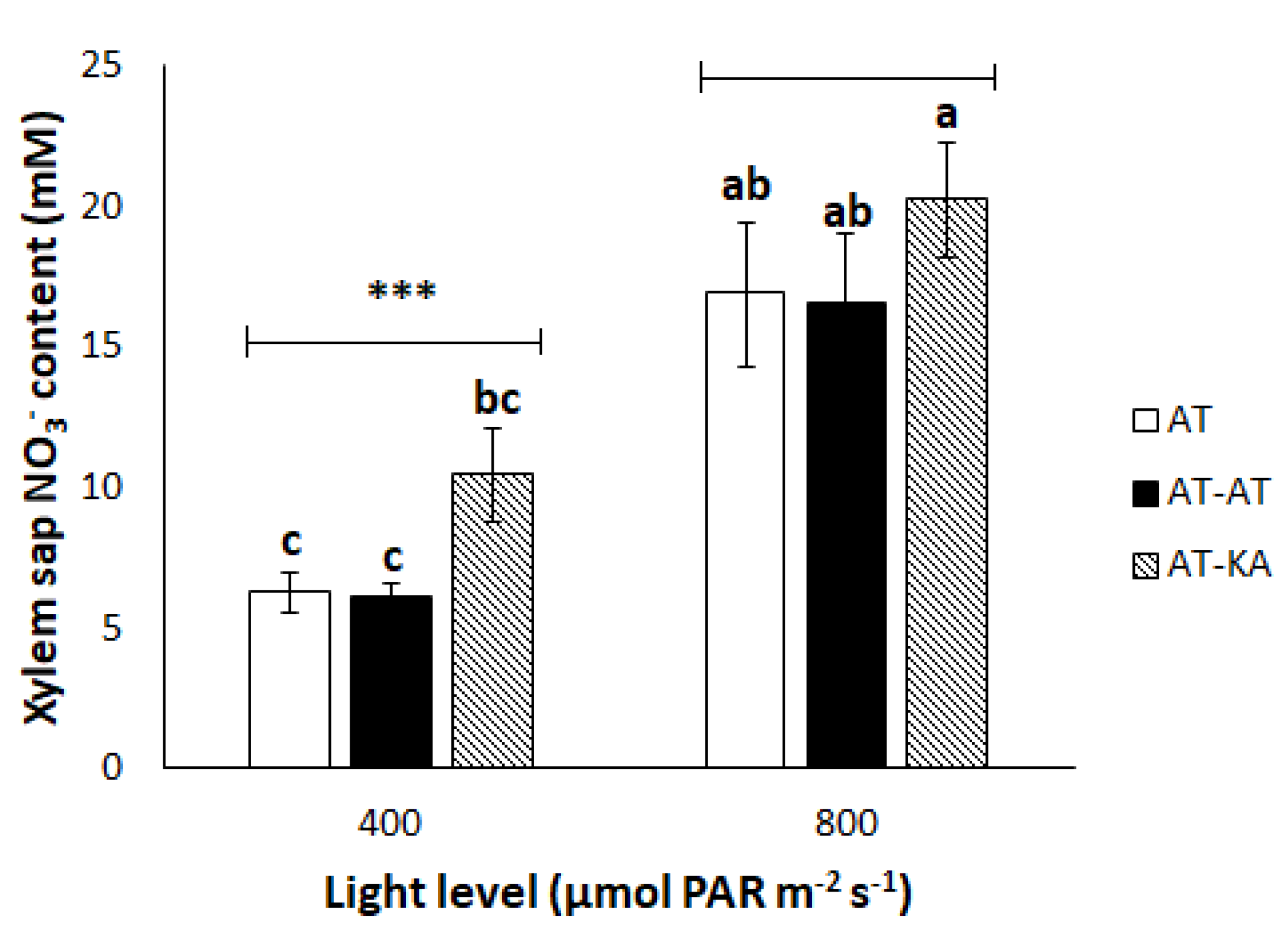
2.2.3. Xylem Sap NO3− Content
2.2.4. Stem Hydraulic Conductance (ks)
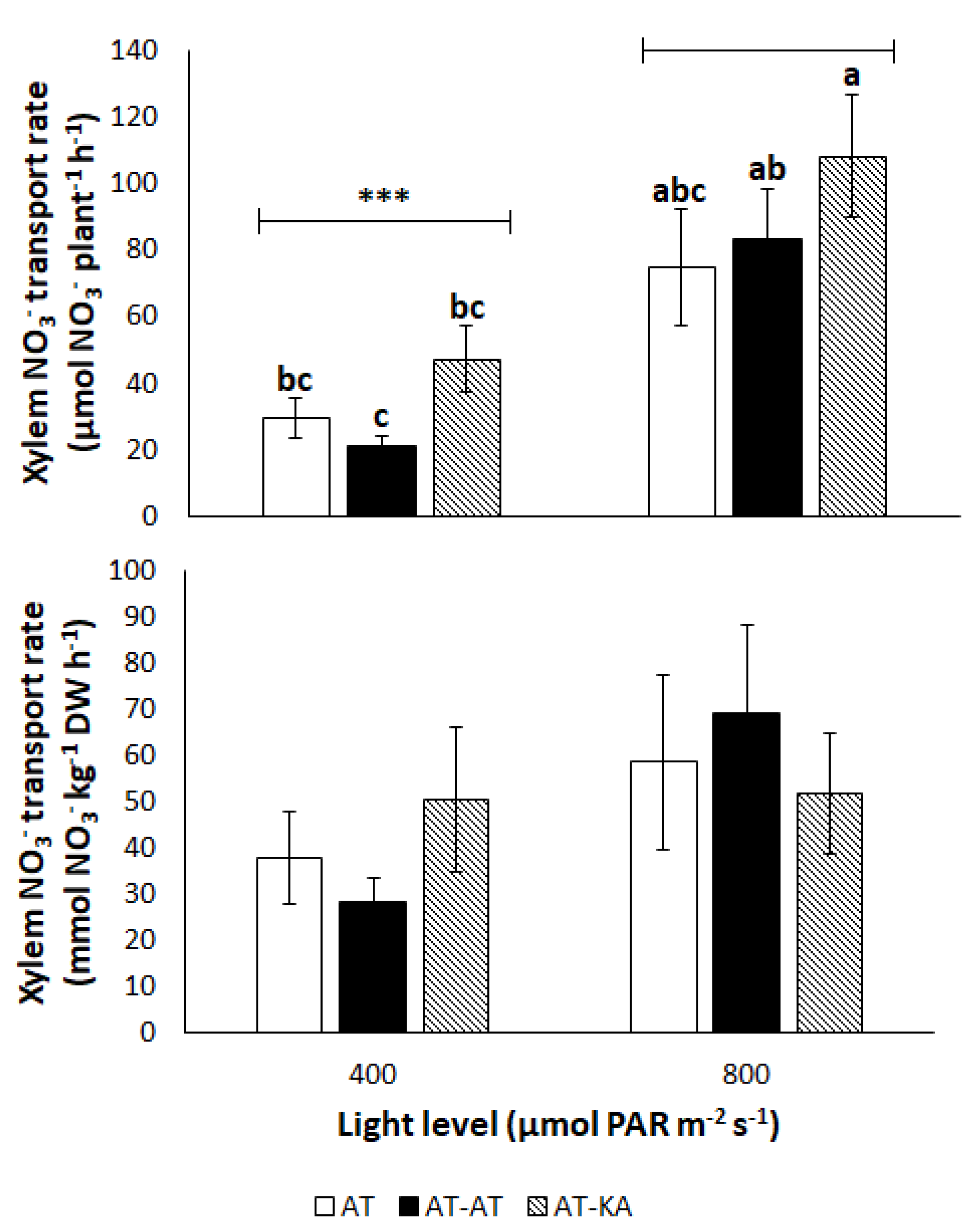
2.2.5. Nitrate Transport Rate
2.2.6. Theoretical Hydraulic Conductance (kh) Calculation
2.3. Statistical Analysis
3. Results
4. Discussion
4.1. Nitrate Content in the Xylem Sap
4.2. Water Transport Rate
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- King, S.R.; Davis, A.R.; Zhang, X.; Crosby, K. Genetic breeding and selection of rootstocks for Solanaceae and Cucurbitaceae. Sci. Hortic. 2010, 127, 106–111. [Google Scholar] [CrossRef]
- Albornoz, F.; Gebauer, M.; Ponce, C.; Cabeza, R. LeNRT1.1 improves nitrate uptake in grafted tomato plants under high nitrogen demand. Int. J. Mol. Sci. 2018, 19, 3921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djidonou, D.; Zhao, X.; Brecht, J.K.; Cordasco, K.M. Influence of interspecific hybrid rootstocks on tomato growth, nutrient accumulation, yield, and fruit composition under greenhouse conditions. HortTechonology 2017, 27, 868–877. [Google Scholar] [CrossRef] [Green Version]
- Marschner, P. Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: San Diego, CA, USA, 2012; pp. 49–70. [Google Scholar]
- Grassi, G.; Millard, P.; Wendler, R.; Minotta, G.; Tagliavini, M. Measurements of xylem sap amino acid concentrations in conjunction with whole tree transpiration estimates spring N remobilization by cherry (Prunus avium L.) trees. Plant Cell Environ. 2002, 25, 1689–1699. [Google Scholar] [CrossRef]
- Djidonou, D.; Lopiano, K.; Zhao, X.; Simonne, E.H.; Erickson, J.E.; Koch, K.E. Estimating nitrogen nutritional crop requirements of grafted tomatoes under field conditions. Sci. Hortic. 2015, 182, 18–26. [Google Scholar] [CrossRef]
- Chen, G.; Fu, X.; Lips, H.; Sagi, M. Control of plant growth resides in the shoot, and not in the root, in reciprocal grafts of flacca and wild-type tomato (Lycopersicon esculentum), in the presence and absence of salinity stress. Plant Soil 2003, 256, 205–215. [Google Scholar] [CrossRef]
- Sánchez-Rodríguez, E.; Leyva, R.; Constán-Aguilar, C.; Romero, L.; Ruíz, J.M. How does grafting affect the ionome of cherry tomato plants under water stress? Soil Sci. Plant Nutr. 2014, 60, 145–155. [Google Scholar] [CrossRef]
- Asins, M.J.; Albacete, A.; Martínez-Andujar, C.; Pérez-Alfocea, F.; Dodd, I.C.; Carbonell, E.A.; Dieleman, J.A. Genetic analysis of rootstock-mediated nitrogen (N) uptake and root-to-shoot signaling at contrasting N availabilities in tomato. Plant Sci. 2017, 263, 94–106. [Google Scholar] [CrossRef] [Green Version]
- Choi, K.J.; Chung, G.C.; Ahn, S.J. Effect of root zone temperature on the mineral composition of xylem sap and plasma membrane K+-Mg++-ATPase activity of grafted-cucumber and –figleaf gourd root systems. Plant Cell Physiol. 1995, 36, 639–643. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, A.; Yamashita, M.; Furuya, S. Mineral concentrations and cytokinin activity in the xylem exudate of grafted watermelons as affected by rootstocks and crop load. J. Japan. Soc. Hort. Sci. 1994, 62, 817–826. [Google Scholar] [CrossRef]
- Salehi, R.; Kashi, A.; Lee, J.; Babalar, M.; Delshad, M.; Lee, S.; Huh, Y. Leaf gas exchange and mineral ion composition in xylem sap of Iranian melon affected by rootstocks and training methods. HortScience 2010, 45, 766–770. [Google Scholar] [CrossRef] [Green Version]
- Albacete, A.; Martínez-Andújar, C.; Ghanem, M.E.; Acosta, M.; Sánchez-Bravo, J.; Asins, M.J.; Cuartero, J.; Lutts, S.; Dodd, I.C.; Pérez-Alfocea, F. Rootstock-mediated changes in xylem ionic and hormonal status are correlated with delayed leaf senescence, and increased leaf area and crop productivity in salinized tomato. Plant Cell Environ. 2009, 32, 928–938. [Google Scholar] [CrossRef]
- Weibel, A.; Johnson, R.S.; DeJong, T.M. Comparative vegetative growth responses of two peach cultivars grown on size-controlling versus standard rootstocks. J. Am. Soc. Hortic. Sci. 2003, 128, 463–471. [Google Scholar] [CrossRef] [Green Version]
- Sellin, A.; Ounapuu, E.; Karusion, A.; Oren, R. Experimental evidence supporting the concept of light-mediated modulation of stem hydraulic conductance. Tree Physiol. 2010, 30, 1528–1535. [Google Scholar] [CrossRef] [Green Version]
- Ragel, P.; Raddatz, N.; Leidi, E.O.; Quintero, F.J.; Pardo, J.M. Regulation of K+ nutrition in plants. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Melcher, P.J.; Holbrook, N.M.; Burns, M.J.; Zwieniecki, M.A.; Cobb, A.R.; Brodribb, T.J.; Choat, B.; Sack, L. Measurements of stem xylem hydraulic conductivity in the laboratory and field. Methods Ecol. Evol. 2012, 3, 685–694. [Google Scholar] [CrossRef]
- Tombesi, S.; Johnson, R.S.; Day, K.R.; DeJong, T.M. Relationship between xylem vessel characteristics, calculated axial hydraulic conductance and size-controlling capacity of peach rootstocks. Ann. Bot. 2010, 105, 327–331. [Google Scholar] [CrossRef] [Green Version]
- Sory, A.; Nieto-Angel, R.; Rodríguez-Pérez, J.E.; Barrientos-Priego, A.F.; Ibañez-Castillo, L.A.; Romabchik, E.; Núñez-Colín, C.A. Variación anatómica del xilema en tallo de cultivares de tomate injertados en un tipo criollo. Rev. Chapingo Ser. Hortic. 2010, 16, 67–76. [Google Scholar]
- Edelstein, M.; Burger, Y.; Horev, C.; Porat, A.; Meir, A.; Cohen, R. Assessing the effect of genetic and anatomic variation of Cucurbita rootstocks on vigor, survival and yield of grafted melons. J. Hortic. Sci. Biotechnol. 2004, 79, 370–374. [Google Scholar] [CrossRef]
- Belda, R.M.; Fenlon, J.S.; Ho, L.C. Salinity effects on the xylem vessels in tomato fruit among cultivars with different susceptibilities to blossom-end rot. J. Hort. Sci. 1996, 71, 173–179. [Google Scholar] [CrossRef]
- Paul, S.; Das, M.K.; Baishya, P.; Ramteke, A.; Farooq, M.; Baroowa, B.; Sunkar, R.; Gogoi, N. Effect of high temperature on yield associated parameters and vascular bundle development in five potato cultivars. Sci. Hortic. 2017, 225, 134–140. [Google Scholar] [CrossRef]
- Pérez-Donoso, A.; Greve, L.C.; Walton, J.H.; Shackel, K.A.; Labavitch, J.M. Xylella fastidiosa infection and ethylene exposure result in xylem and water movement disruption in grapevine shoots. Plant Physiol. 2007, 143, 1024–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansen, D.A. Plant Microtechnique, 1st ed.; McGraw-Hill: New York, NY, USA, 1940; pp. 65–94. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2008. [Google Scholar]
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Robledo, C.W. InfoStat Version 2014; Grupo InfoStat, Universidad Nacional de Córdoba: Córdoba, Argentina, 2014. [Google Scholar]
- Miller, A.J.; Shen, Q.; Xu, G. Freeways in the plant: Transporters for N, P and S and their regulation. Curr. Opin. Plant Biol. 2009, 12, 284–290. [Google Scholar] [CrossRef]
- Orieux, C.; Demarest, G.; Decau, M.; Beauclair, P.; Bataille, M.; Le Deunff, E. Changes in 15NO3− availability and transpiration rate are associated with a rapid diurnal adjustment of anion contents as well as 15N and water fluxes between the roots and shoots. Front. Plant Sci. 2018, 9, 1751. [Google Scholar] [CrossRef]
- Galleguillos-Cedillo, V.M.; Urrestarazu, M.; Álvaro, J.E. Influence of salinity on transport of nitrates and potassium by means of the xylem sap content between roots and shoots in young tomato plants. J. Soil Sci. Plant Nutr. 2016, 16, 991–998. [Google Scholar]
- Rivas-Ubach, A.; Sardans, J.; Pérez-Trujillo, M.; Estiarte, M.; Peñuelas, J. Strong relationship between elemental stoichiometry and metabolome in plants. Proc. Natl. Acad. Sci. USA 2012, 109, 4181–4186. [Google Scholar] [CrossRef] [Green Version]
- Zgallaï, H.; Steppe, K.; Lemeur, R. Photosynthetic, physiological and biochemical responses of tomato plants to polyethylene glycol-induced water deficit. J. Integr. Plant Biol. 2005, 47, 1470–1478. [Google Scholar] [CrossRef]
- Leonardi, C.; Giuffrida, F. Variation of plant growth and macronutrient uptake in grafted tomatoes and eggplants on three different rootstocks. Europ. J. Hort. Sci. 2006, 71, 97–101. [Google Scholar]
- Savvas, D.; Öztekin, G.B.; Tepecik, M.; Ropokis, A.; Tüzel, Y.; Ntatsi, G.; Schwarz, D. Impact of grafting and rootstock on nutrient-to-water uptake ratios during the first month after planting of hydroponically grown tomato. J. Hort. Sci. Biotechnol. 2017, 92, 294–302. [Google Scholar] [CrossRef]
- Djidonou, D.; Zhao, X.; Koch, K.E.; Zotarelli, L. Nitrogen accumulation and root distribution of grafted tomato plants as affected by nitrogen fertilization. HortScience 2019, 54, 1907–1914. [Google Scholar] [CrossRef] [Green Version]
- Borgognone, D.; Colla, G.; Rouphael, Y.; Cardarelli, M.; Rea, E.; Schwarz, D. Effect of nitrogen form and nutrient solution pH on growth and mineral composition of self-grafted and grafted tomatoes. Sci. Hortic. 2013, 149, 61–69. [Google Scholar] [CrossRef]
- De Groot, C.C.; Marcelis, L.F.M.; Van Den Boogaard, R.; Lambers, H. Growth and dry-mass partitioning in tomato as affected by phosphorus nutrition and light. Plant Cell Environ. 2001, 24, 1309–1317. [Google Scholar] [CrossRef]
- Xiaoying, L.; Shirong, G.; Taotao, C.; Zhigang, X.; Tezuka, T. Regulation of the growth and photosynthesis of cherry tomato seedlings by different light irradiations of light emitting diodes (LED). African J. Biotechnol. 2012, 11, 6169–6177. [Google Scholar] [CrossRef]
- Melnyk, C.W.; Schuster, C.; Leyser, O.; Meyerowitz, E.M. A developmental framework for graft formation and vascular reconnection in Arabidopsis thaliana. Curr. Biol. 2015, 25, 1306–1318. [Google Scholar] [CrossRef] [Green Version]
- Melnyk, C.W. Plant grafting: Insights into tissue regeneration. Regeneration 2017, 4, 3–14. [Google Scholar] [CrossRef]
- Fernandez-Garcia, N.; Carvajal, M.; Olmos, E. Graft union formation in tomato plants: Peroxidase and catalase involvement. Ann. Bot. 2004, 93, 53–60. [Google Scholar] [CrossRef]
- Zwieniecki, M.; Melcher, P.J.; Holbrook, N.M. Hydrogel control of xylem hydraulic resistance in plants. Science 2001, 291, 1059–1062. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | 400 µmol m−2 s−1 | 800 µmol m−2 s−1 | ||
|---|---|---|---|---|
| Plant Dry Weight (g) | N Content (mg N g−1 DW) | Plant Dry Weight (g) | N Content (mg N g−1 DW) | |
| AT | 0.82 ± 0.05 b | 46.8 ± 1.7 a | 1.69 ± 0.24 ab | 39.6 ± 3.9 a |
| AT-AT | 0.77 ± 0.05 b | 42.5 ± 2.2 a | 1.56 ± 0.22 b | 36.3 ± 1.3 a |
| AT-KA | 1.11 ± 0.13 a | 45.6 ± 2.1 a | 2.32 ± 0.18 a | 33.7 ± 2.1 a |
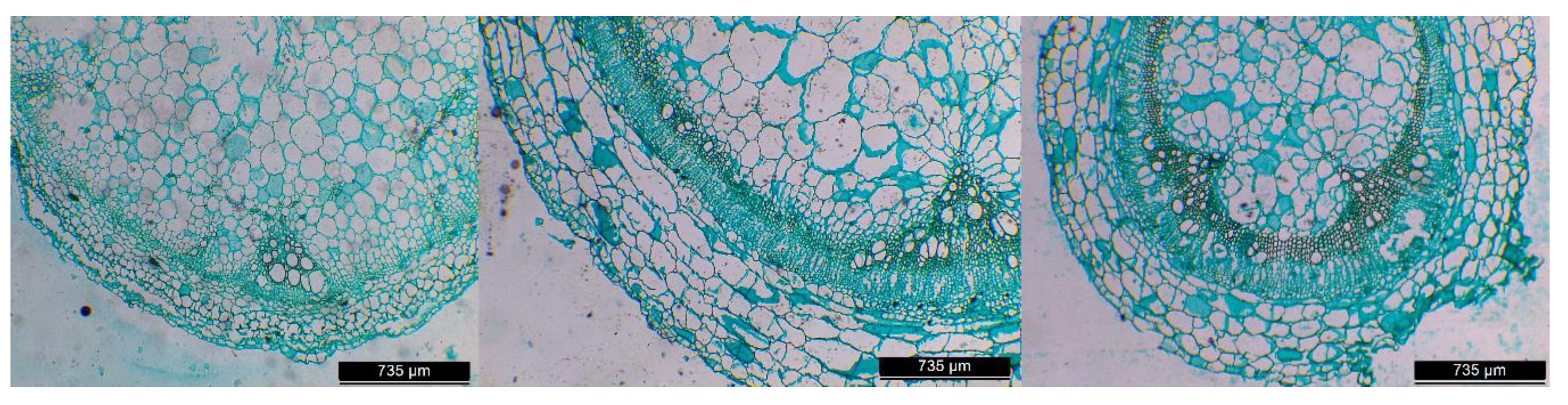
| Treatment | Vessel Number (N° mm−2) | Mean Vessel Diameter (µm) | kh (kg m MPa−1 s−1) | ks (kg MPa−1 m−1 s−1) |
|---|---|---|---|---|
| AT | 23.5 ± 1.65 c | 62.04 ± 10.14 ab | 5.8 x 10−7 b | 0.560 ± 0.213 a |
| AT-AT | 45.5 ± 0.86 b | 54.90 ± 7.46 b | 6.8 x 10−7 b | 0.207 ± 0.048 b |
| AT-KA | 76.2 ± 4.78 a | 79.31 ± 14.44 a | 6.9 x 10−6 a | 0.090 ± 0.025 c |
| WU | N_T | N_Xyl | Ves_D | Ves_N | RGR | |
|---|---|---|---|---|---|---|
| WU | - | 0.53 *** | −0.10 n.s. | −0.03 n.s. | −0.14 n.s. | −0.40 ** |
| N_T | 0.53 *** | - | 0.72 *** | 0.16 n.s. | 0.03 n.s. | 0.11 n.s. |
| N_Xyl | −0.10 n.s. | 0.72 *** | - | 0.19 n.s. | 0.20 n.s. | 0.46 ** |
| Ves_D | −0.03 n.s. | 0.16 n.s. | 0.19 n.s. | - | 0.33 * | 0.25 n.s. |
| Ves_N | −0.14 n.s. | 0.03 n.s. | 0.20 n.s. | 0.33 * | - | 0.26 n.s. |
| RGR | −0.40 ** | 0.11 n.s. | 0.46 ** | 0.25 n.s. | 0.26 n.s. | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Albornoz, F.; Pérez-Donoso, A.G.; Leigh Urbina, J.; Monasterio, M.; Gómez, M.; Steinfort, Ú. Nitrate Transport Rate in the Xylem of Tomato Plants Grafted onto a Vigorous Rootstock. Agronomy 2020, 10, 182. https://doi.org/10.3390/agronomy10020182
Albornoz F, Pérez-Donoso AG, Leigh Urbina J, Monasterio M, Gómez M, Steinfort Ú. Nitrate Transport Rate in the Xylem of Tomato Plants Grafted onto a Vigorous Rootstock. Agronomy. 2020; 10(2):182. https://doi.org/10.3390/agronomy10020182
Chicago/Turabian StyleAlbornoz, Francisco, Alonso G. Pérez-Donoso, Jorge Leigh Urbina, Matías Monasterio, Miguel Gómez, and Úrsula Steinfort. 2020. "Nitrate Transport Rate in the Xylem of Tomato Plants Grafted onto a Vigorous Rootstock" Agronomy 10, no. 2: 182. https://doi.org/10.3390/agronomy10020182