Photosynthetic and Agronomic Traits of Winter Barley (Hordeum vulgare L.) Varieties




Abstract
:1. Introduction
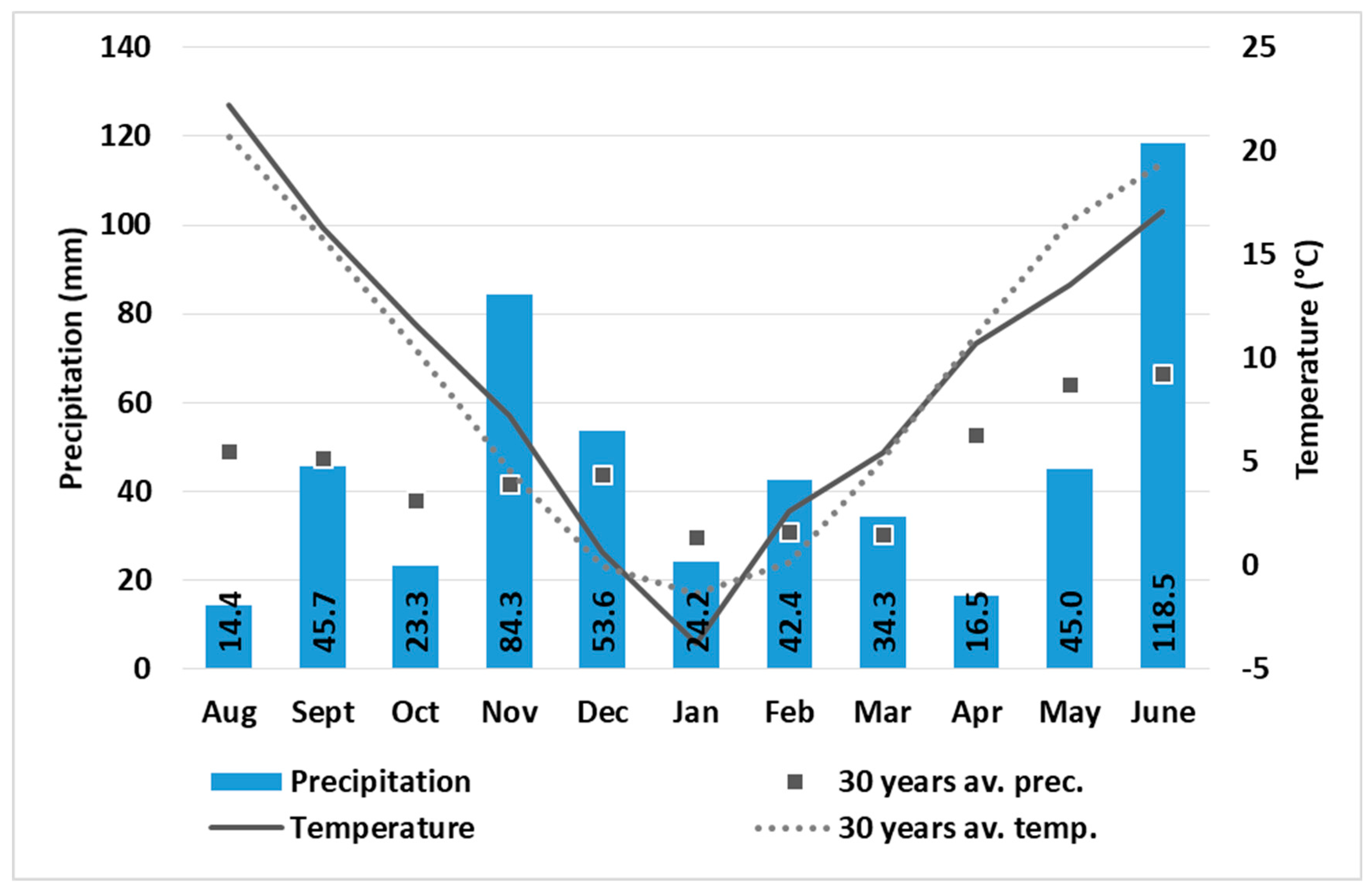
2. Materials and Methods
3. Results
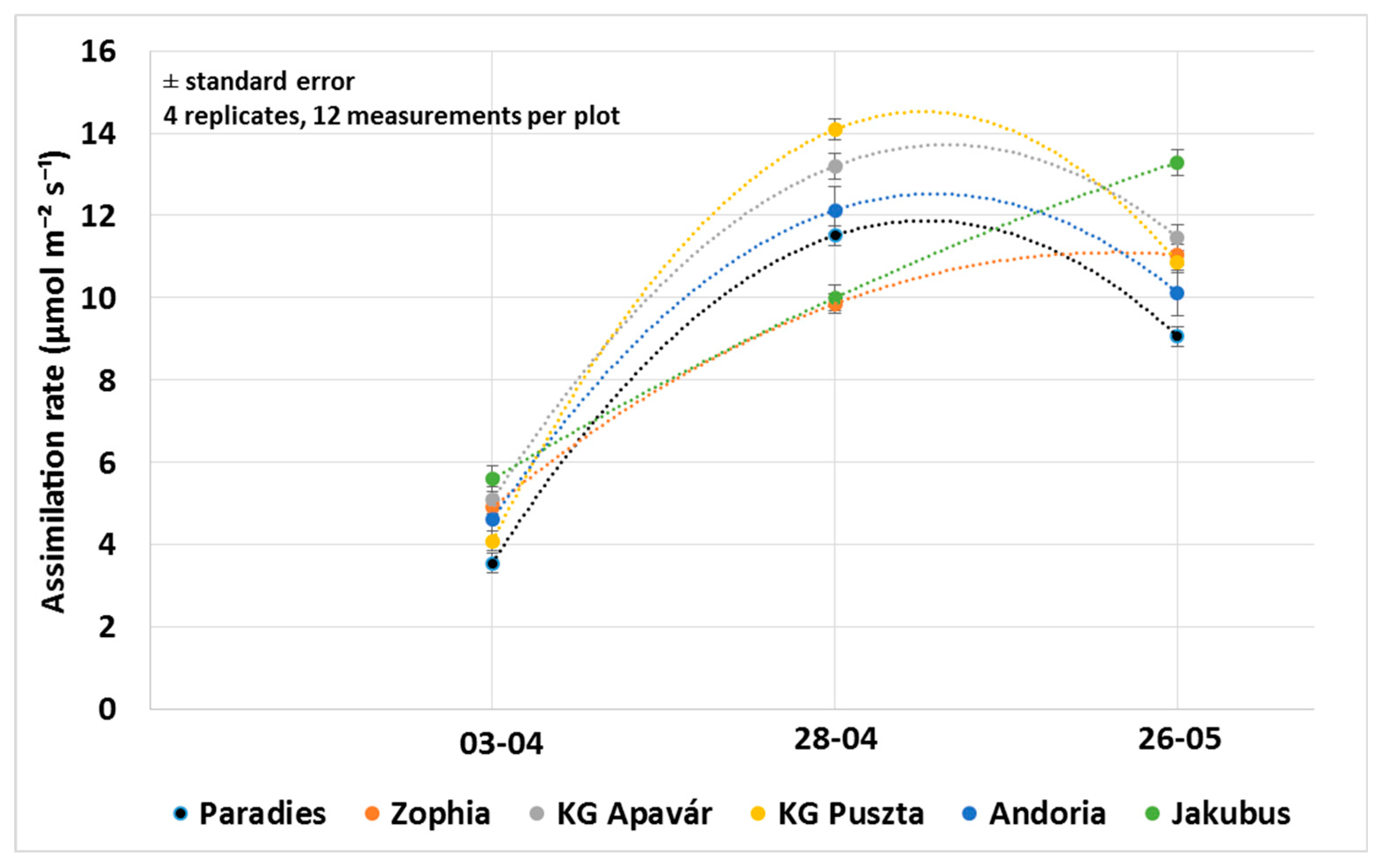
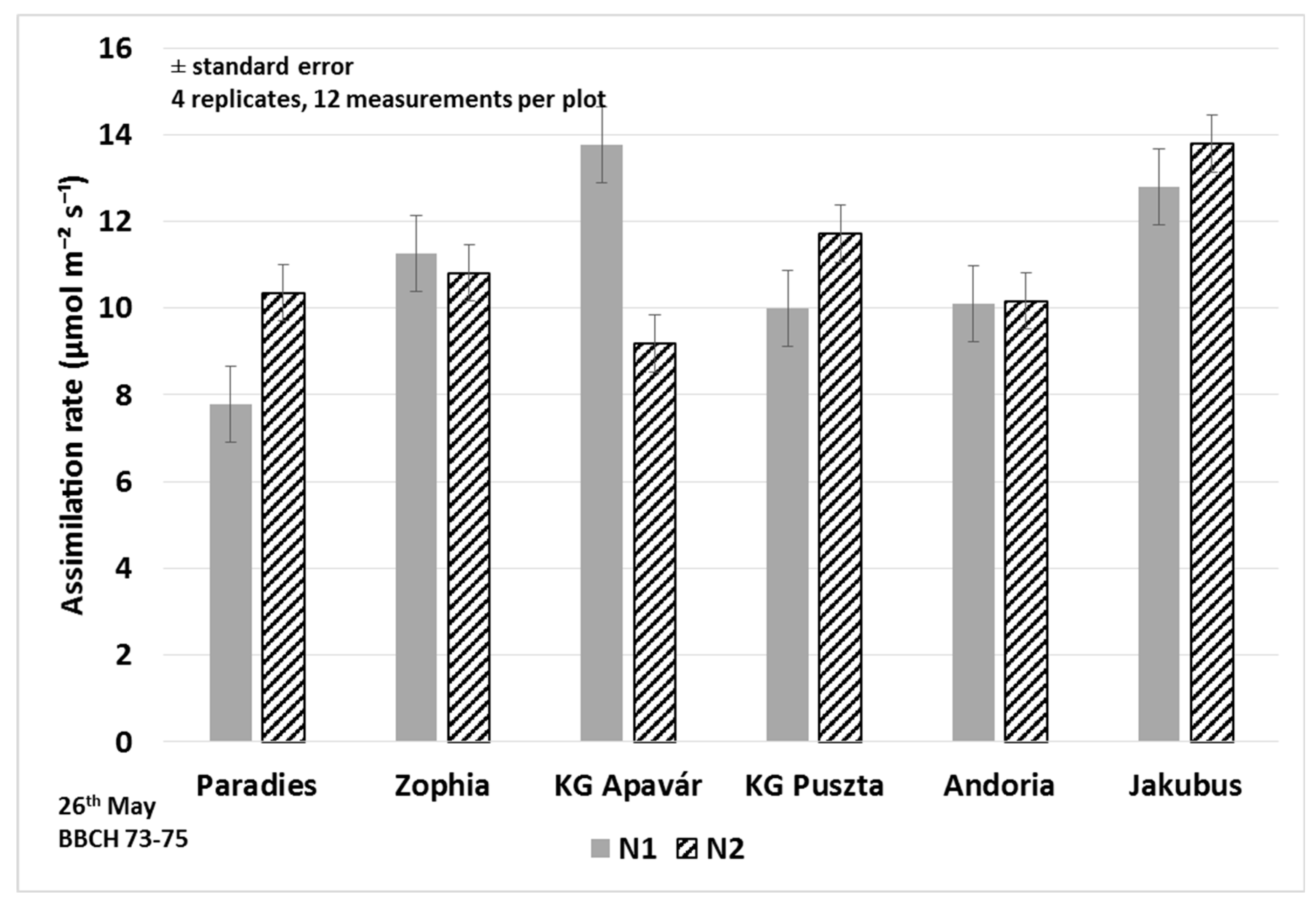
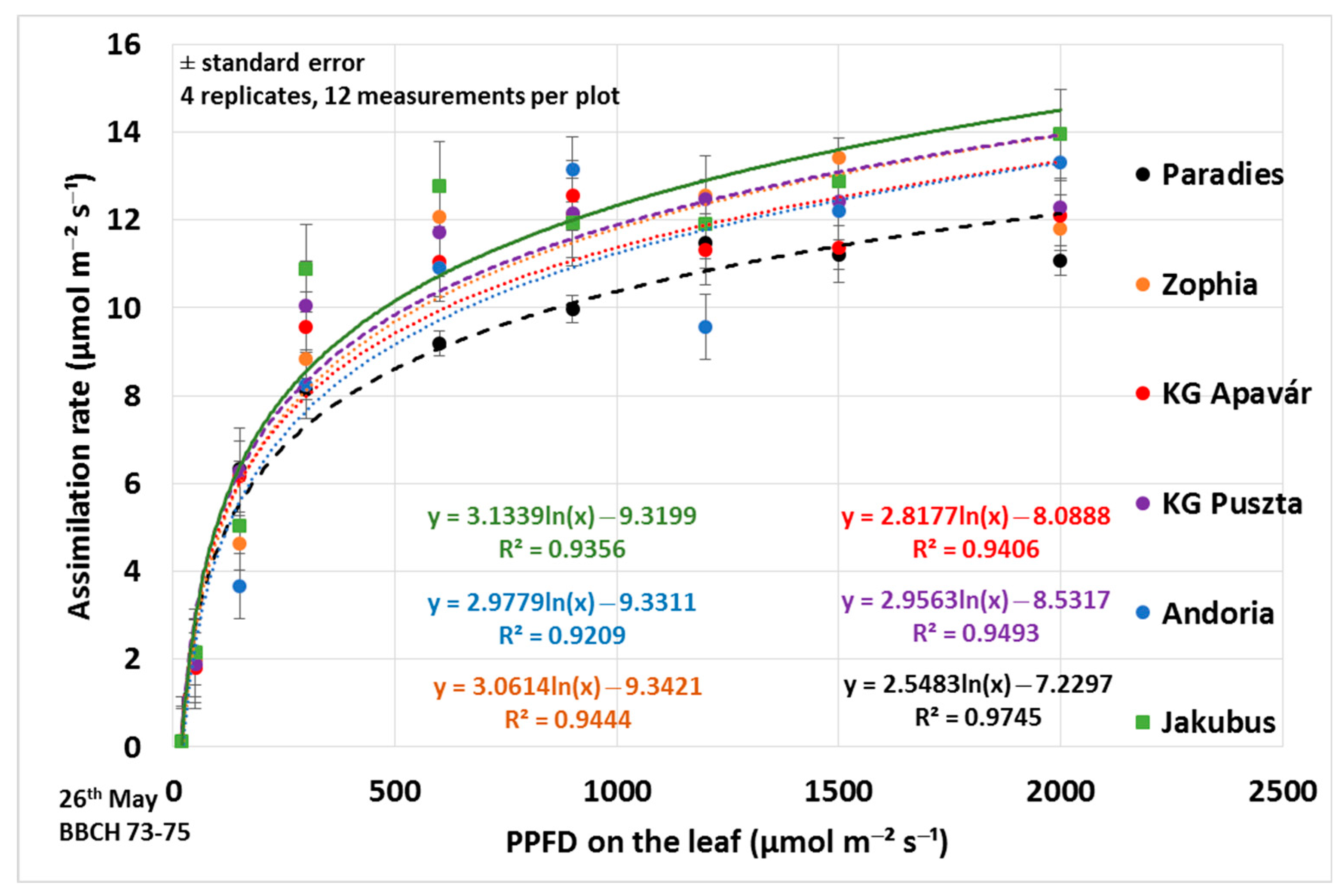
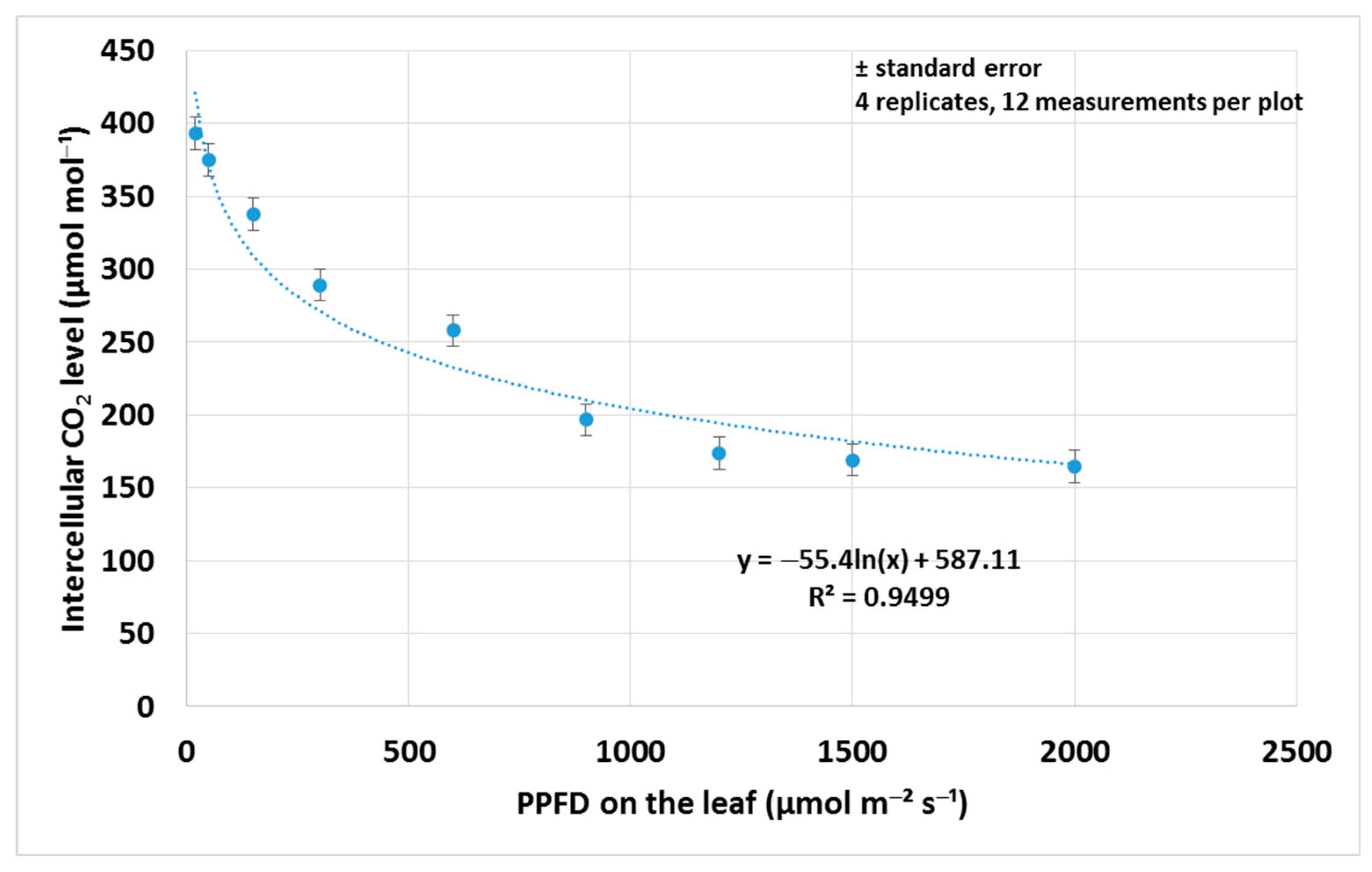
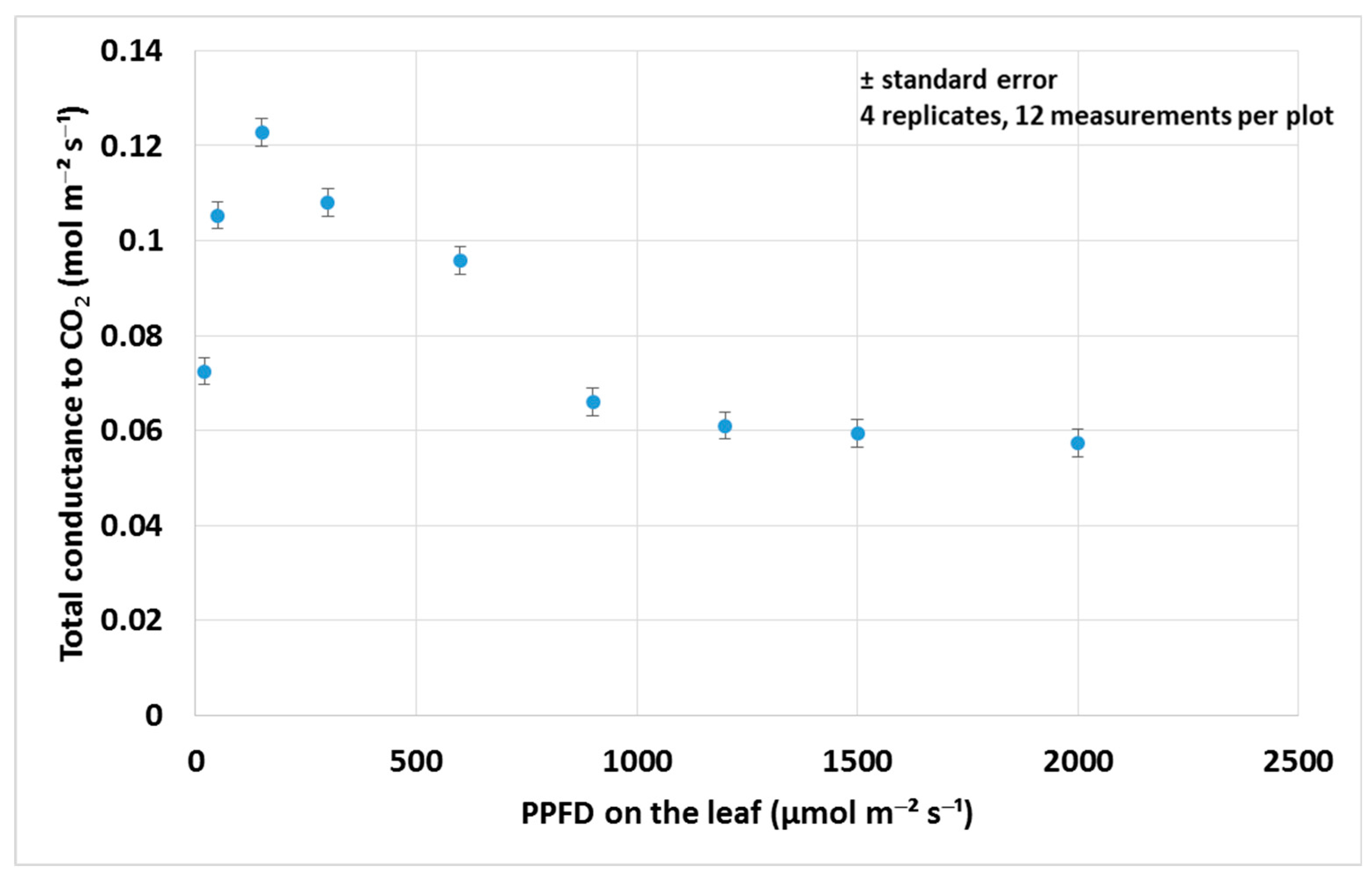
3.1. Assimilation Parameters
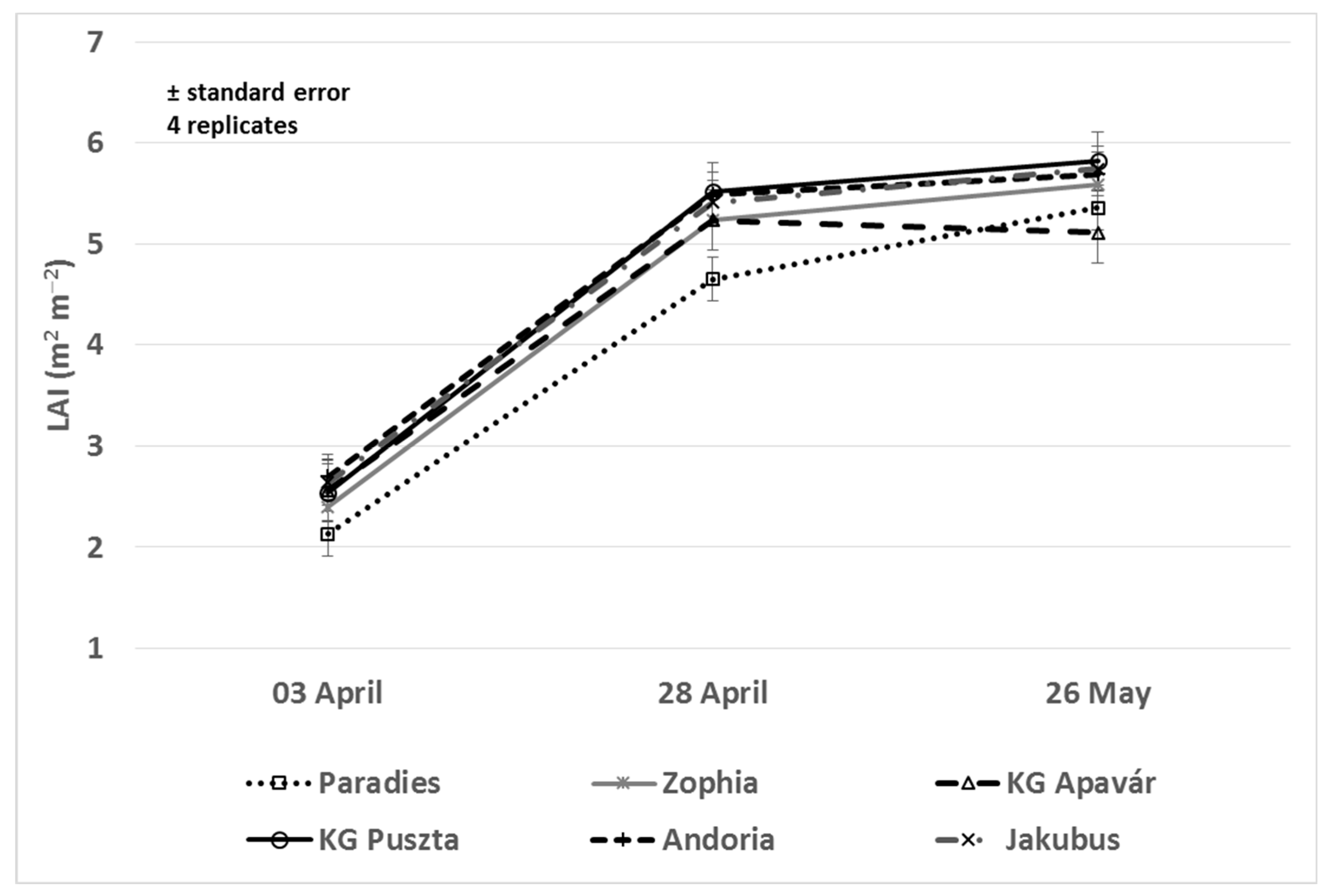
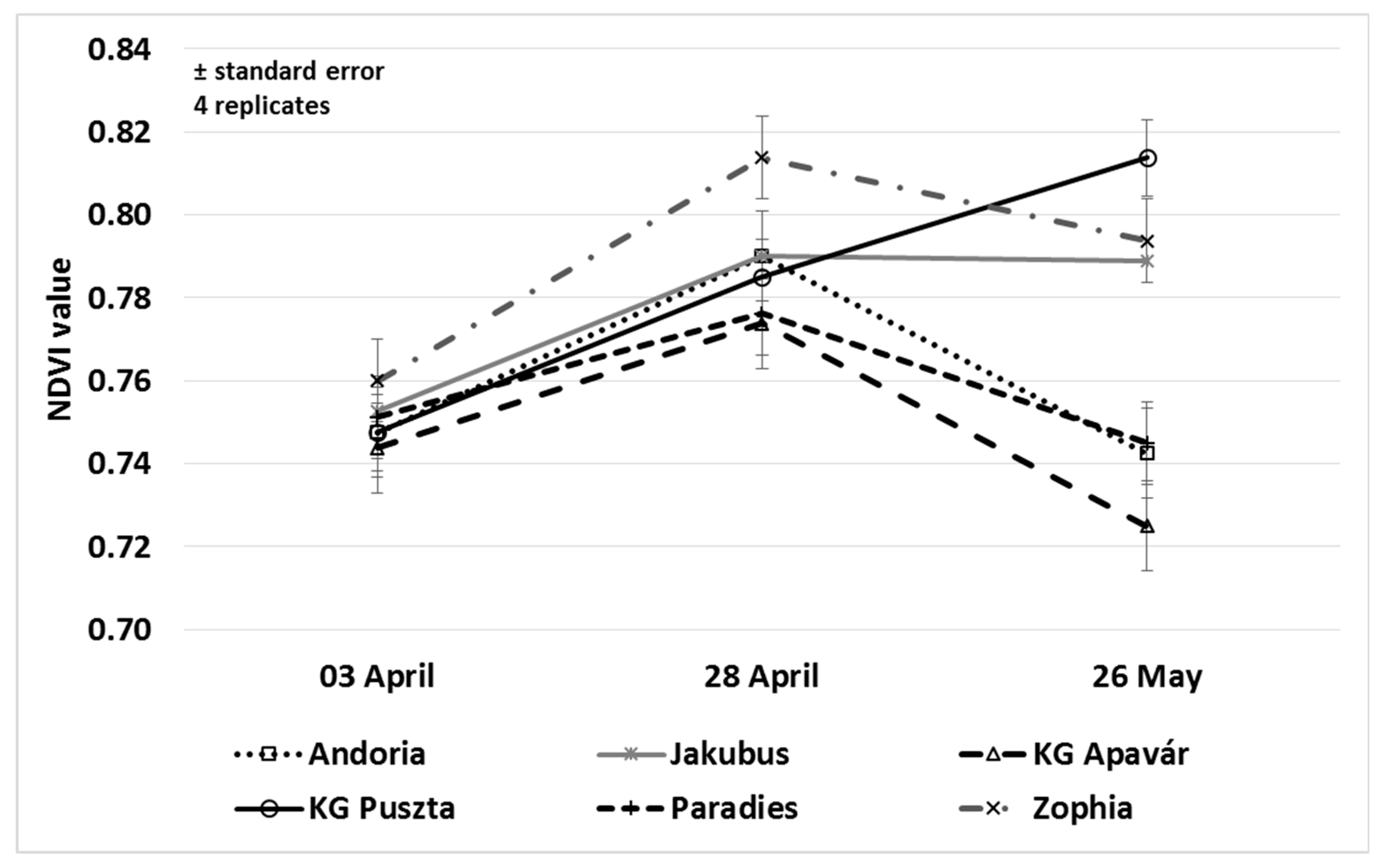
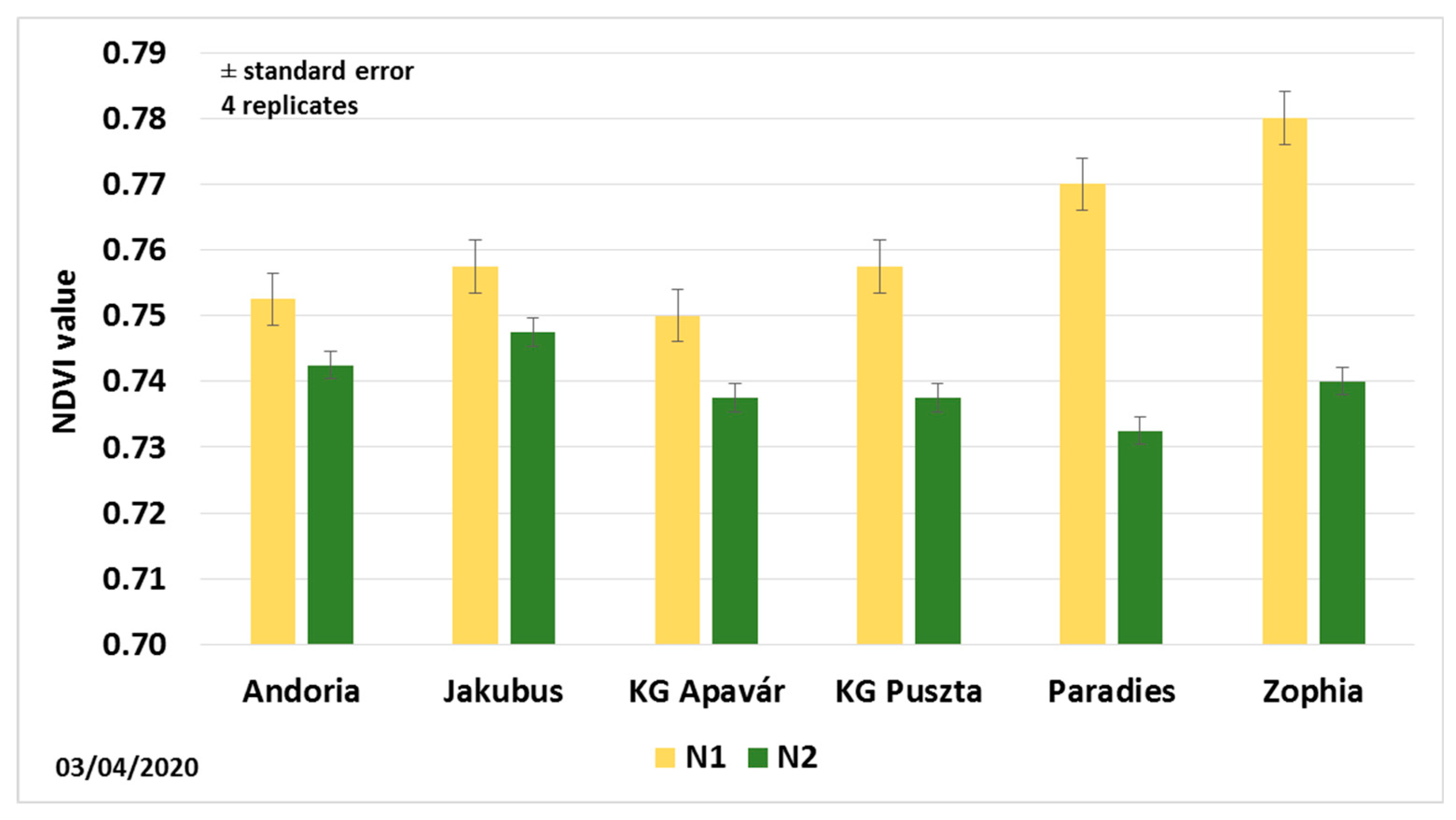
3.2. Leaf Area and NDVI Values
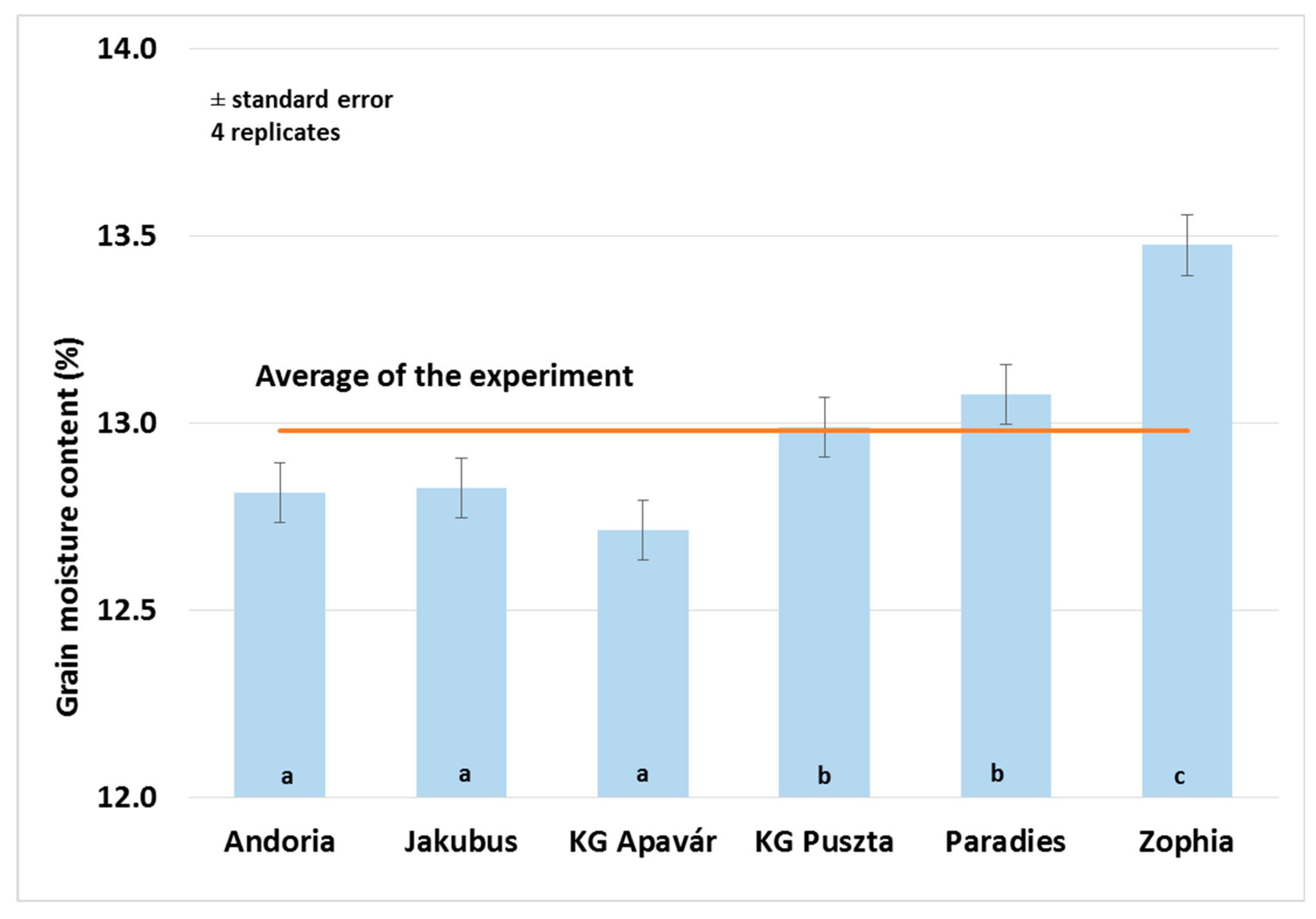
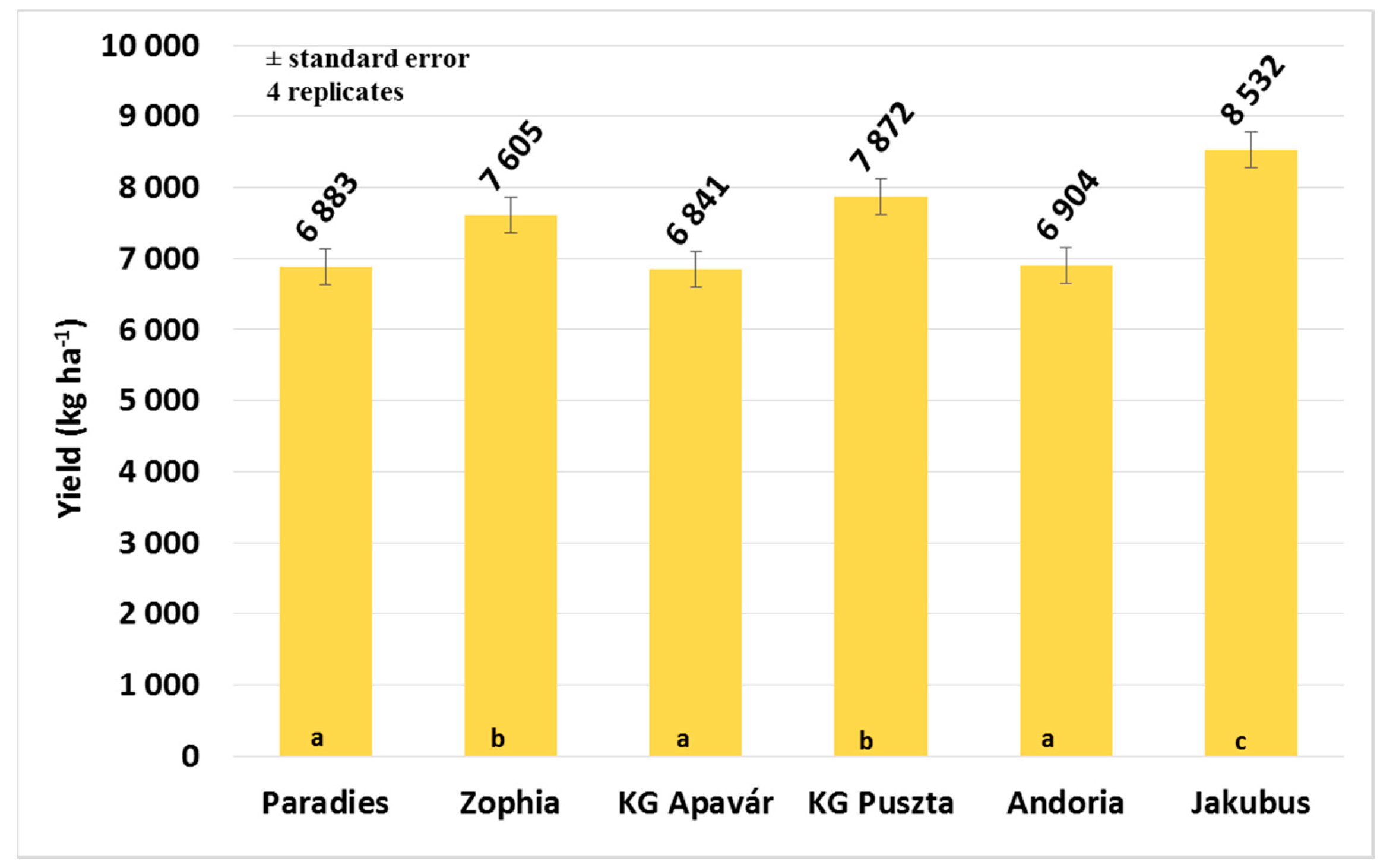
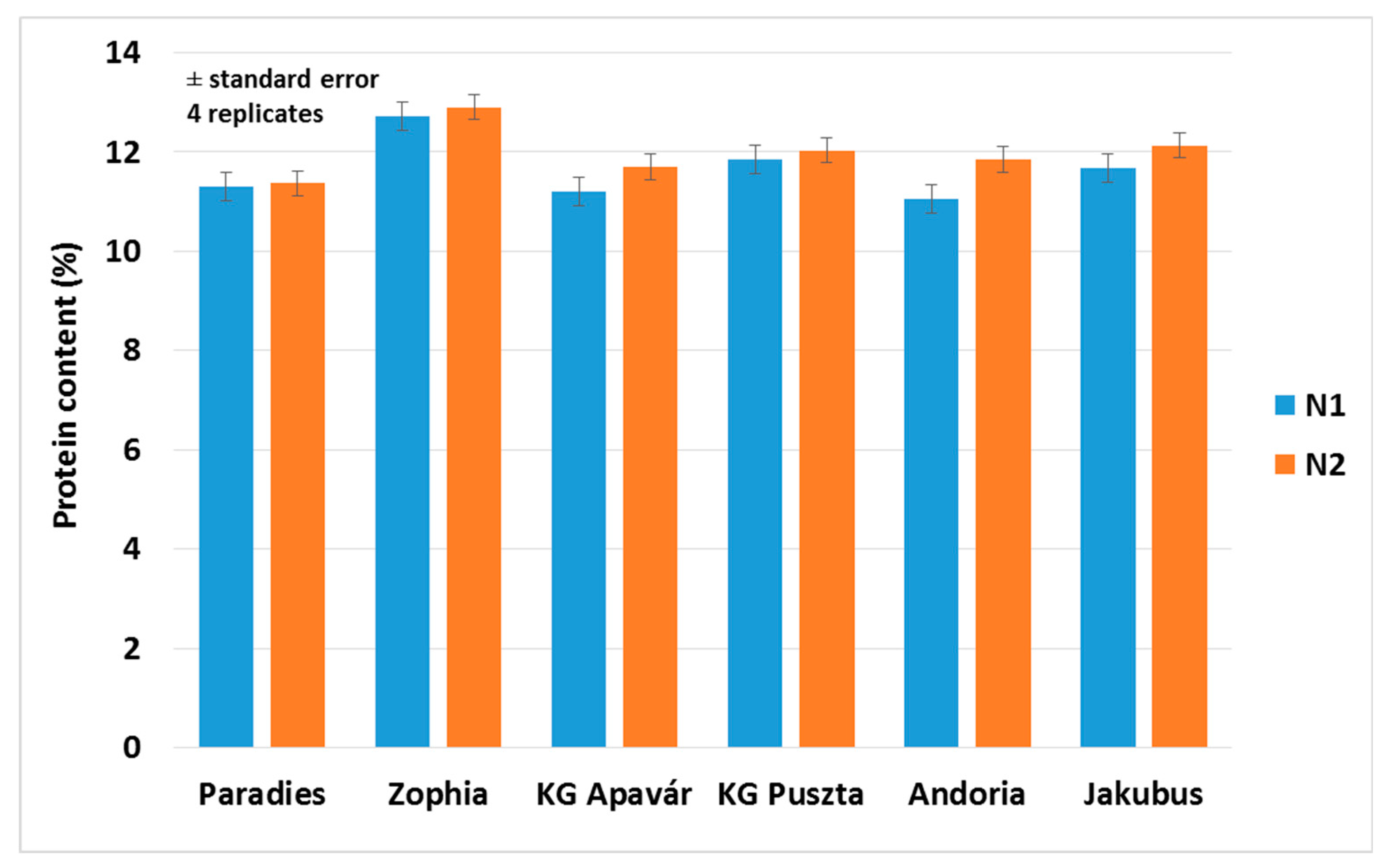
3.3. Yield and Quality
3.4. Connections between Assimilation Parameters and Yield and Quality
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lopes, M.S.; Nogués, S.; Araus, J.L. Nitrogen source and water regime effects on barley photosynthesis and isotope signature. Funct. Plant Biol. 2004, 31, 995. [Google Scholar] [CrossRef] [PubMed]
- Robredo, A.; Pérez-López, U.; De La Maza, H.S.; González-Moro, B.; Lacuesta, M.; Mena-Petite, A.; Muñoz-Rueda, A. Elevated CO2 alleviates the impact of drought on barley improving water status by lowering stomatal conductance and delaying its effects on photosynthesis. Environ. Exp. Bot. 2007, 59, 252–263. [Google Scholar] [CrossRef]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food Security: The Challenge of Feeding 9 Billion People. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newton, A.; Flavell, A.J.; George, T.S.; Leat, P.; Mullholland, B.; Ramsay, L.; Revoredo-Giha, C.; Russell, J.; Steffenson, B.J.; Swanston, J.S.; et al. Crops that feed the world 4. Barley: A resilient crop? Strengths and weaknesses in the context of food security. Food Secur. 2011, 3, 141–178. [Google Scholar] [CrossRef]
- Giraldo, P.; Benavente, E.; Manzano-Agugliaro, F.; Gimenez, E. Worldwide Research Trends on Wheat and Barley: A Bibliometric Comparative Analysis. Agronomy 2019, 9, 352. [Google Scholar] [CrossRef] [Green Version]
- Bertholdsson, N.-O. Screening for Barley Waterlogging Tolerance in Nordic Barley Cultivars (Hordeum vulgare L.) Using Chlorophyll Fluorescence on Hydroponically-Grown Plants. Agronomy 2013, 3, 376–390. [Google Scholar] [CrossRef] [Green Version]
- Al-Ajlouni, Z.I.; Al-Abdallat, A.M.; Al-Ghzawi, A.L.A.; Ayad, J.Y.; Abu Elenein, J.M.; Al-Quraan, N.A.; Baenziger, P.S. Impact of Pre-Anthesis Water Deficit on Yield and Yield Components in Barley (Hordeum vulgare L.) Plants Grown under Controlled Conditions. Agronomy 2016, 6, 33. [Google Scholar] [CrossRef] [Green Version]
- Carter, A.Y.; Hawes, M.C.; Ottman, M.J. Drought-Tolerant Barley: I. Field Observations of Growth and Development. Agronomy 2019, 9, 221. [Google Scholar] [CrossRef] [Green Version]
- Degl’Innocenti, E.; Hafsi, C.; Guidi, L.; Navari-Izzo, F. The effect of salinity on photosynthetic activity in potassium-deficient barley species. J. Plant Physiol. 2009, 166, 1968–1981. [Google Scholar] [CrossRef]
- Ahmed, I.M.; Nadira, U.A.; Zhang, G.; Wu, F. Exploration and Utilization of Drought-Tolerant Barley Germplasm; Academic Press: Cambridge, MA, USA, 2016; pp. 115–152. [Google Scholar]
- Sallam, A.; Alqudah, A.M.; Dawood, M.F.; Baenziger, P.S.; Börner, A. Drought Stress Tolerance in Wheat and Barley: Advances in Physiology, Breeding and Genetics Research. Int. J. Mol. Sci. 2019, 20, 3137. [Google Scholar] [CrossRef] [Green Version]
- Doré, T.; Makowski, D.; Malézieux, E.; Munier-Jolain, N.; Tchamitchian, M.; Tittonell, P. Facing up to the paradigm of ecological intensification in agronomy: Revisiting methods, concepts and knowledge. Eur. J. Agron. 2011, 34, 197–210. [Google Scholar] [CrossRef]
- Tittonell, P. Ecological intensification of agriculture—Sustainable by nature. Curr. Opin. Environ. Sustain. 2014, 8, 53–61. [Google Scholar] [CrossRef]
- Kovacevic, J.; Mazur, M.; Lalić, A.; Josipovic, M.; Josipovic, A.; Kocar, M.M.; Markovic, M.; Antunović, J.; Cesar, V. Photosynthetic performance index in early stage of growth, water use efficiency, and grain yield of winter barley cultivars. Chil. J. Agric. Res. 2015, 75, 275–283. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Mai, B.; Wu, R.; Feng, Y.; Sofo, A.; Ni, Y.; Sun, J.; Li, J.; Xu, J. Acclimation of winter wheat (Triticum aestivum, cv. Yangmai 13) to low levels of solar irradiance. Photosynthetica 2011, 49, 426. [Google Scholar] [CrossRef]
- Kadar, I.; Csathó, P. N-Cu interactions in a field experiment on spring barley. Agrokém. Talajt. 2013, 62, 345–358. [Google Scholar] [CrossRef]
- Drew, M.C. Comparison of the effects of a localized supply of phosphate, nitrate, ammonium and potassium on the growth of the seminal root system, and the shoot, in barley. New Phytol. 1975, 75, 479–490. [Google Scholar] [CrossRef]
- Siebrecht, S.; Tischner, R.; Mäck, G. Function and contribution of the root tip in the induction of NO3− uptake along the barley root axis. J. Exp. Bot. 1995, 46, 1669–1676. [Google Scholar] [CrossRef]
- Perry, E.M.; Fitzgerald, G.; Nuttall, J.G.; O’Leary, G.J.; Schulthess, U.; Whitlock, A. Rapid estimation of canopy nitrogen of cereal crops at paddock scale using a Canopy Chlorophyll Content Index. Field Crop. Res. 2012, 134, 158–164. [Google Scholar] [CrossRef]
- Csajbók, J.; Kutasy, E. Results of N Supply and Plant Protection Testing Experiment on Winter Barley Varieties. In Proceedings of the 17th Alps-Adria Scientific Workshop, Hnanice, Czech Republic, 9 April 2018; pp. 134–135. [Google Scholar]
- Oukarroum, A.; El Madidi, S.; Schansker, G.; Strasser, R.J. Probing the responses of barley cultivars (Hordeum vulgare L.) by chlorophyll a fluorescence OLKJIP under drought stress and re-watering. Environ. Exp. Bot. 2007, 60, 438–446. [Google Scholar] [CrossRef]
- Hejnak, V.; Zamecnikova, B.; Neckarova, J. The influence of high natrium and chlorine ion concentration on physiological responses of various spring barley varieties. Turk. J. Field Crops 2011, 16, 93–98. [Google Scholar]
- Hein, J.A.; Sherrard, M.E.; Manfredi, K.P.; Abebe, T. The fifth leaf and spike organs of barley (Hordeum vulgare L.) display different physiological and metabolic responses to drought stress. BMC Plant Biol. 2016, 16, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, T.; Yang, F.; Das, D.; Chen, M.; Hu, Q.; Tian, Y.; Cheng, C.; Liu, Y.; Zhang, J. Transcriptomic analysis of photosynthesis-related genes regulated by alternate wetting and drying irrigation in flag leaves of rice. Food Energy Secur. 2020, 9. [Google Scholar] [CrossRef]
- Blum, A. Selection for Sustained Production in Water Deficit Environments. In International Crop Science I; Crop Science Society of America (CSSA): Madison, WI, USA, 1993; pp. 343–347. [Google Scholar]
- Arenas-Corraliza, M.G.; Rolo, V.; López-Díaz, M.L.; Moreno, G. Wheat and barley can increase grain yield in shade through acclimation of physiological and morphological traits in Mediterranean conditions. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hack, H.; Bleiholder, H.; Buhr, L.; Meier, U.; Schnock-Fricke, U.; Weber, E.; Witzenberger, A. Einheitliche Codierung der Phänologischen Entwicklungsstadien mono- und dikotyler Pflanzen Erweiterte BBCH-Skala, Allgemein. Nachrichtenbl. Deut. Pflanzenschutzd. 1992, 44, 265–270. [Google Scholar]
- Von Caemmerer, S.; Farquhar, G.D. Some relationships between the biochemistry of photosynthesis and the gas exchange of leaves. Planta 1981, 153, 376–387. [Google Scholar] [CrossRef]
- LI-COR, Inc. Li-6800 Portable Photosynthesis System, Software Version 1.2; LI-COR, Inc.: Lincoln, NE, USA, 2017. [Google Scholar]
- Rouse, W.J.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring Vegetation Systems in the Great Plains with ERTS. In Proceedings of the Third ERTS Symposium, Washington DC, USA, 10–14 December 1973; pp. 309–317. [Google Scholar]
- Blum, A. Photosynthesis and Transpiration in Leaves and Ears of Wheat and Barley Varieties. J. Exp. Bot. 1985, 36, 432–440. [Google Scholar] [CrossRef]
- Pang, G.; Xu, Z.; Wang, T.; Cong, X.; Wang, H. Photosynthetic light response characteristics of winter wheat at heading and flowering stages under saline water irrigation. IOP Conf. Ser. Earth Environ. Sci. 2018, 170. [Google Scholar] [CrossRef] [Green Version]
- Vos, J.; Groenwold, J. Characteristics of photosynthesis and conductance of potato canopies and the effects of cultivars and transient drought. Field Crop. Res. 1989, 20, 237–250. [Google Scholar] [CrossRef]
- Flexas, J.; Medrano, H. Drought inhibition of photosynthesis in C3 plants: Stomatal limitations revisited. Ann. Bot. 2002, 89, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Kutasy, E.; Csajbók, J.; Borbély, É.H. Relations between yield and photosynthetic activity of winter wheat varieties. Cereal Res. Commun. 2005, 33, 173–176. [Google Scholar] [CrossRef]
- Mejri, M.; Siddique, K.H.M.; Saif, T.; Abdelly, C.; Hessini, K. Comparative effect of drought duration on growth, photosynthesis, water relations, and solute accumulation in wild and cultivated barley species. J. Plant Nutr. Soil Sci. 2016, 179, 327–335. [Google Scholar] [CrossRef]
- Küpper, P.M.; Perkons, U.; Pries, C.; Antar, P.M.; Kautz, T.; Köpke, U. Rates of photosynthesis and transpiration of wheat and barley as influenced by fodder precrops and their cropping period. In Proceedings of the 4th ISOFAR Scientific Conference—‘Building Organic Bridges’, Istanbul, Turkey, 13–15 October 2014. [Google Scholar]
- Allel, D.; Ben-Amar, A.; Abdelly, C. Leaf photosynthesis, chlorophyll fluorescence and ion content of barley (Hordeum vulgare) in response to salinity. J. Plant Nutr. 2017, 41, 497–508. [Google Scholar] [CrossRef]
- Panfilova, A.; Korkhova, M.; Gamayunova, V.; Fedorchuk, M.; Drobitko, A.; Nikonchuk, N.; Kovalenko, O. Formation of photosynthetic and grain yield of spring barley (Hordeum vulgare L.) depend on varietal characteristics and plant growth regulators. Agron. Res. 2019, 17, 608–620. [Google Scholar] [CrossRef]
- Chapin, F.S.; Walter, C.H.S.; Clarkson, D.T. Growth response of barley and tomato to nitrogen stress and its control by abscisic acid, water relations and photosynthesis. Planta 1988, 173, 352–366. [Google Scholar] [CrossRef]
- Cai, J.; Jiang, D.; Wollenweber, B.; Dai, T.; Cao, W. Effects of nitrogen application rate on dry matter redistribution, grain yield, nitrogen use efficiency and photosynthesis in malting barley. Acta Agric. Scand. Sect. B Plant Soil Sci. 2012, 62, 1–10. [Google Scholar] [CrossRef]
- Dubey, S.; Tiwari, A.; Pandey, V.; Singh, V.; Singh, G. Effect of nitrogen levels and its time of application on growth parameters of barley (Hordeum vulgare L.). J. Pharmacogn. Phytochem. 2018, 7, 333–338. [Google Scholar] [CrossRef]
- Paul, T.E.M.; Madan, G. Sensor-based algorithms to improve barley nitrogen efficiency in Queensland. Afr. J. Agric. Res. 2018, 13, 1476–1486. [Google Scholar] [CrossRef]
- Fetch, T.G.; Steffenson, B.J., Jr.; Pederson, V.D. Predicting agronomic performance of barley using canopy reflectance data. Can. J. Plant Sci. 2004, 84, 1–9. [Google Scholar] [CrossRef]
- Lalic, A.; Kovacevic, J.; Novoselovic, D.; Simic, G.; Abicic, I.; Guberac, V. Agronomic and quantitative traits of winter barley varieties (Hordeum vulgare L.) under growing conditions in Croatia. Agric. Conspec. Sci. 2009, 74, 283–289. [Google Scholar]
- Ram, H.; Dhaliwal, S.S. Effect of varieties and integrated nutrient management techniques on growth, productivity, quality and economics of barley (Hordeum vulgare L.). Int. J. Agric. Sci. 2012, 8, 91–97. [Google Scholar]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Layer 0–25 cm | Layer 25–50 cm | Layer 50–75 cm | Layer 75–100 cm | Layer 100–130 cm | |
|---|---|---|---|---|---|
| pH (KCl) | 6.46 | 6.36 | 6.58 | 7.27 | 7.36 |
| KA | 43 | 44.6 | 47.6 | 46.6 | 45.4 |
| CaCO3 (%) | 0 | 0 | 0 | 10.25 | 12.75 |
| Humus (%) | 2.76 | 2.16 | 1.52 | 0.9 | 0.59 |
| Organic C (%) | 1.60 | 1.25 | 0.88 | 0.52 | 0.34 |
| Total N (%) | 0.15 | 0.12 | 0.086 | 0.083 | 0.078 |
| NO3 + NO2 (mg kg−1) | 6.2 | 1.74 | 0.6 | 1.92 | 1.78 |
| P2O5 (AL) (mg kg−1) | 133.4 | 48 | 40.4 | 39.8 | 31.6 |
| K2O (AL) (mg kg−1) | 239.8 | 173.6 | 123 | 93.6 | 78 |
| Mg (mg kg−1) | 332.4 | 405.4 | 366.6 | 249 | 286.6 |
| Na (mg kg−1) | 38 | 66.2 | 55.4 | 67.8 | 62.6 |
| Zn (mg kg−1) | 2.8 | 0.8 | 0.58 | 0.48 | 0.84 |
| Cu (mg kg−1) | 5.86 | 4.54 | 3.64 | 2.24 | 1.64 |
| Mn (mg kg−1) | 438 | 406 | 339 | 74 | 4 |
| SO4 (mg kg−1) | 9.25 | 9.13 | 10.8 | 7.95 | 22.98 |
| Soil Layer | Bulk Density (g cm−3) | Porosity % | Field Capacity w w−1 % | Permanent Wilting Point w w−1 % | hy |
|---|---|---|---|---|---|
| 5–25 cm | 1.46 | 45.0 | 33.3 | 15.55 | 2.715 |
| 27–33 cm | 1.40 | 47.3 | 37.3 | 15.70 | 2.783 |
| 47–53 cm | 1.23 | 53.6 | 38.3 | 14.75 | 2.755 |
| 72–78 cm | 1.24 | 53.0 | 38.9 | 14.20 | 2.563 |
| 97–103 cm | 1.26 | 52.6 | 40.6 | 11.13 | 2.168 |
| 122–128 cm | 1.28 | 51.8 | 42.3 | 9.38 | 1.853 |
| 147–153 cm | 1.25 | 52.8 | 44.6 | 9.03 | 1.778 |
| 197–203 cm | 1.23 | 53.7 | 46.0 | 8.50 | 1.690 |
| Ass | Transp | GTW | GBW | GSW | |
|---|---|---|---|---|---|
| Correlation Coefficients (3 April, BBCH 27−29) | |||||
| Transp | 0.077 | 1 | 0.927 ** | 0.028 | 0.931 ** |
| GTW | 0.027 | 0.927 ** | 1 | −0.066 | 0.999 ** |
| GBW | −0.210 | 0.028 | −0.066 | 1 | −0.062 |
| GSW | 0.032 | 0.931 ** | 0.999 ** | −0.062 | 1 |
| Correlation Coefficients (28 April, BBCH 47−49) | |||||
| Transp | 0.297 * | 1 | 0.932 ** | 0.029 | 0.935 ** |
| GTW | 0.255 | 0.932 ** | 1 | −0.066 | 0.998 ** |
| GBW | 0.067 | 0.029 | −0.066 | 1 | −0.063 |
| GSW | 0.252 | 0.935 ** | 0.998 ** | −0.063 | 1 |
| Correlation Coefficients (26 May, BBCH 73−75) | |||||
| Transp | 0.662 ** | 1 | 0.992 ** | 0.047 | 0.991 ** |
| GTW | 0.667 ** | 0.992 ** | 1 | 0.041 | 0.999 ** |
| GBW | 0.012 | 0.047 | 0.041 | 1 | 0.038 |
| GSW | 0.667 ** | 0.991 ** | 0.999 ** | 0.038 | 1 |
| Ass | Ci | Phi CO2 | Transp | Tair-Tleaf | Phi PS2 | ETR | GTC | |
|---|---|---|---|---|---|---|---|---|
| PAR | 0.764 * | −0.921 ** | −0.667 * | −0.133 | −0.996 ** | 0.357 | −0.970 ** | −0.765 * |
| Ci | −0.931 ** | 1 | 0.578 | 0.094 | 0.885 ** | −0.452 | 0.903 ** | 0.688 * |
| Phi CO2 | −0.356 | 0.578 | 1 | 0.788 * | 0.699 * | −0.463 | 0.641 | 0.949 ** |
| Transp | 0.119 | 0.094 | 0.788 * | 1 | 0.174 | −0.127 | 0.155 | 0.727 * |
| Tair−Tleaf | −0.703 * | 0.885 ** | 0.699 * | 0.174 | 1 | −0.337 | 0.969 ** | 0.790 * |
| Phi PS2 | 0.534 | −0.452 | −0.463 | −0.127 | −0.337 | 1 | −0.204 | −0.308 |
| ETR | −0.715 * | 0.903 ** | 0.641 | 0.155 | 0.969 ** | −0.204 | 1 | 0.776 * |
| GTC | −0.424 | 0.688 * | 0.949 ** | 0.727 * | 0.790 * | −0.308 | 0.776 | 1 |
| Nitrogen Dose (kg ha−1) | Paradies | Zophia | KG Apavár | KG Puszta | Andoria | Jakubus |
|---|---|---|---|---|---|---|
| NDVI values (3 April) | ||||||
| N1:N90P45K50 | 0.770 a | 0.780 a | 0.750 a | 0.758 a | 0.753 a | 0.758 a |
| N2:N150P45K50 | 0.733 a | 0.740 a | 0.738 a | 0.738 a | 0.743 a | 0.748 a |
| NDVI values (28 April) | ||||||
| N1:N90P45K50 | 0.795 a | 0.828 b | 0.780 c | 0.788 c | 0.788 c | 0.788 c |
| N2:N150P45K50 | 0.758 a | 0.800 b | 0.768 a | 0.783 b | 0.793 c | 0.793 c |
| NDVI values (26 May) | ||||||
| N1:N90P45K50 | 0.753 a | 0.793 b | 0.715 c | 0.820 d | 0.740 a | 0.775 b |
| N2:N150P45K50 | 0.738 a | 0.795 b | 0.735 a | 0.808 b | 0.745 a | 0.803 b |
| Nitrogen Dose (kg ha−1) | Protein Yield (kg ha−1) | |||||
|---|---|---|---|---|---|---|
| Paradies | Zophia | KG Apavár | KG Puszta | Andoria | Jakubus | |
| N1: N90P45K50 | 795.0 a | 916.3 b | 762.5 c | 850.2 d | 737.7 c | 984.0 b |
| N2: N150P45K50 | 763.6 a | 1020.7 b | 797.3 a | 1020.1 b | 844.0 c | 1049.9 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Csajbók, J.; Pepó, P.; Kutasy, E. Photosynthetic and Agronomic Traits of Winter Barley (Hordeum vulgare L.) Varieties. Agronomy 2020, 10, 1999. https://doi.org/10.3390/agronomy10121999
Csajbók J, Pepó P, Kutasy E. Photosynthetic and Agronomic Traits of Winter Barley (Hordeum vulgare L.) Varieties. Agronomy. 2020; 10(12):1999. https://doi.org/10.3390/agronomy10121999
Chicago/Turabian StyleCsajbók, József, Péter Pepó, and Erika Kutasy. 2020. "Photosynthetic and Agronomic Traits of Winter Barley (Hordeum vulgare L.) Varieties" Agronomy 10, no. 12: 1999. https://doi.org/10.3390/agronomy10121999