Shelf-Life of Bunched Carrots as Affected by Nitrogen Fertilization and Leaf Presence



Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Field, Plant Material, and Crop Management
2.2. Plant Harvest, Post-Harvest Handling and Sampling
2.3. Determinations and Calculations
2.3.1. Physicochemical Traits
2.3.2. Compositional Traits
2.4. Statistical Procedure
2.5. Meteorological Conditions during the Experiment
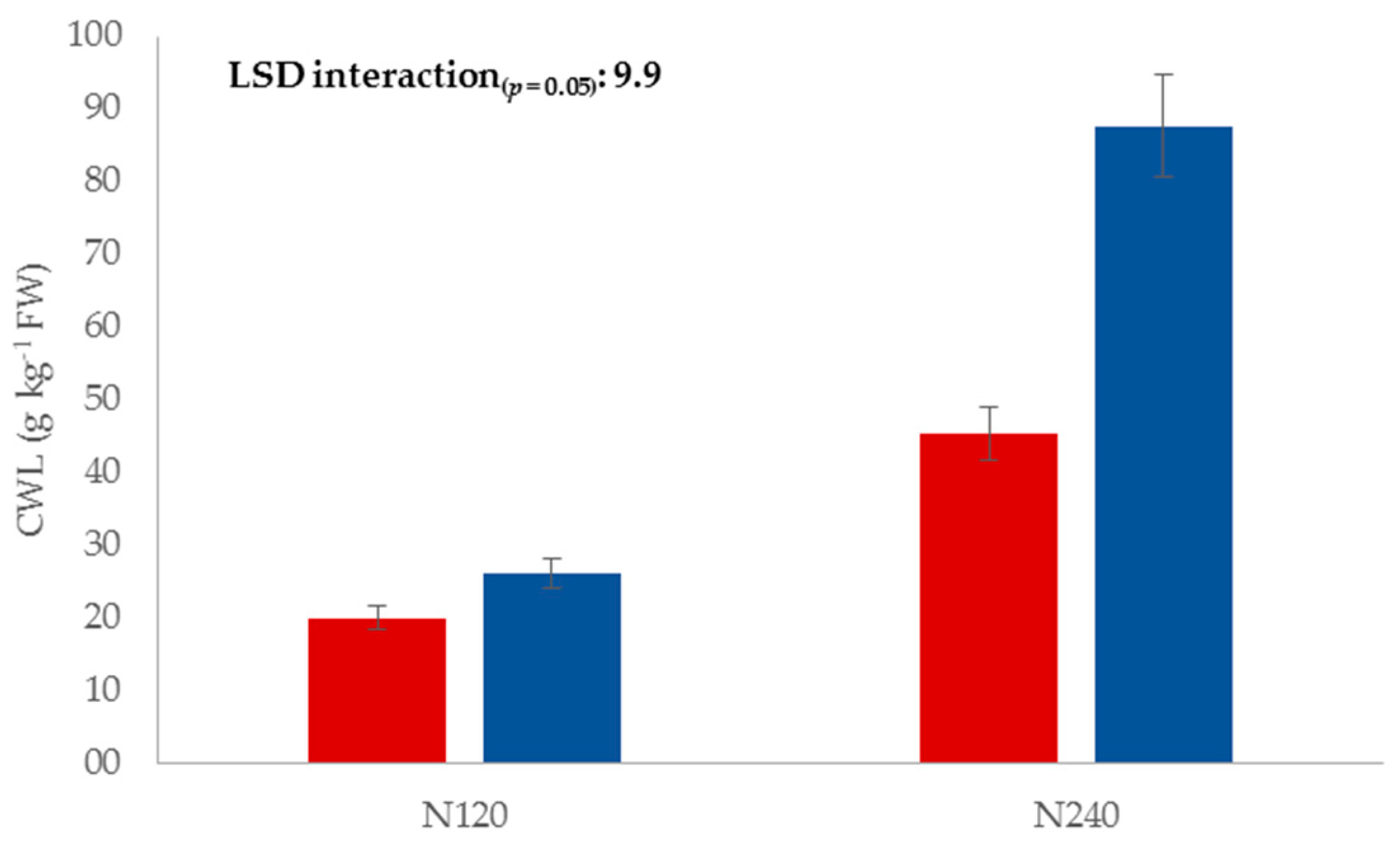
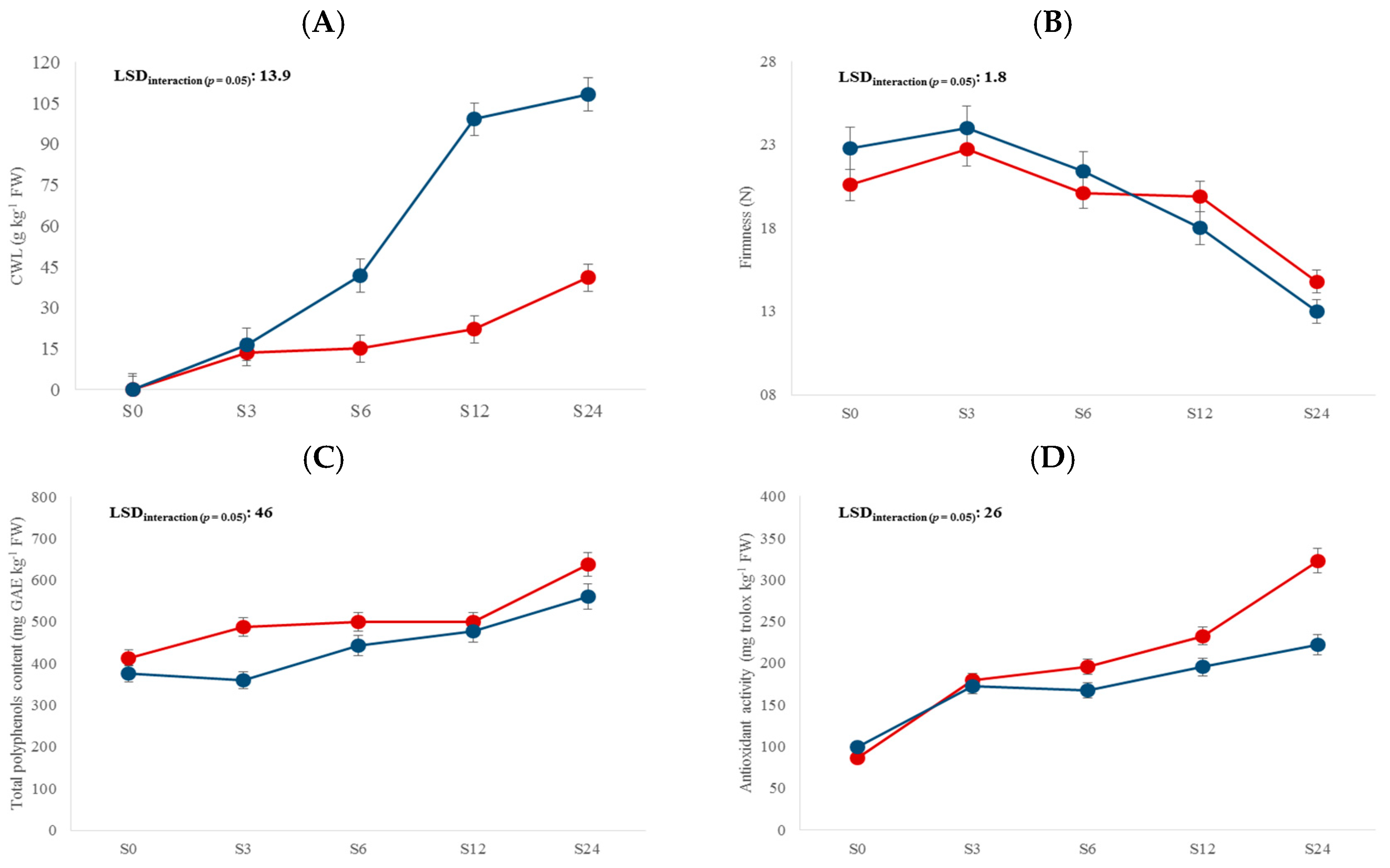
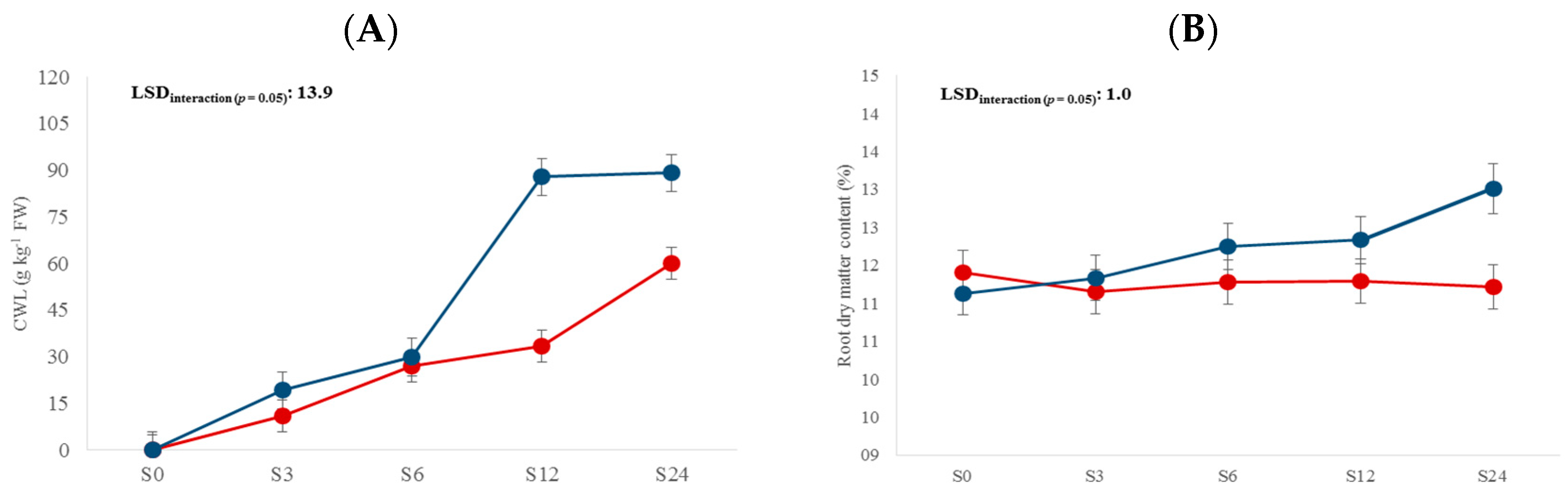
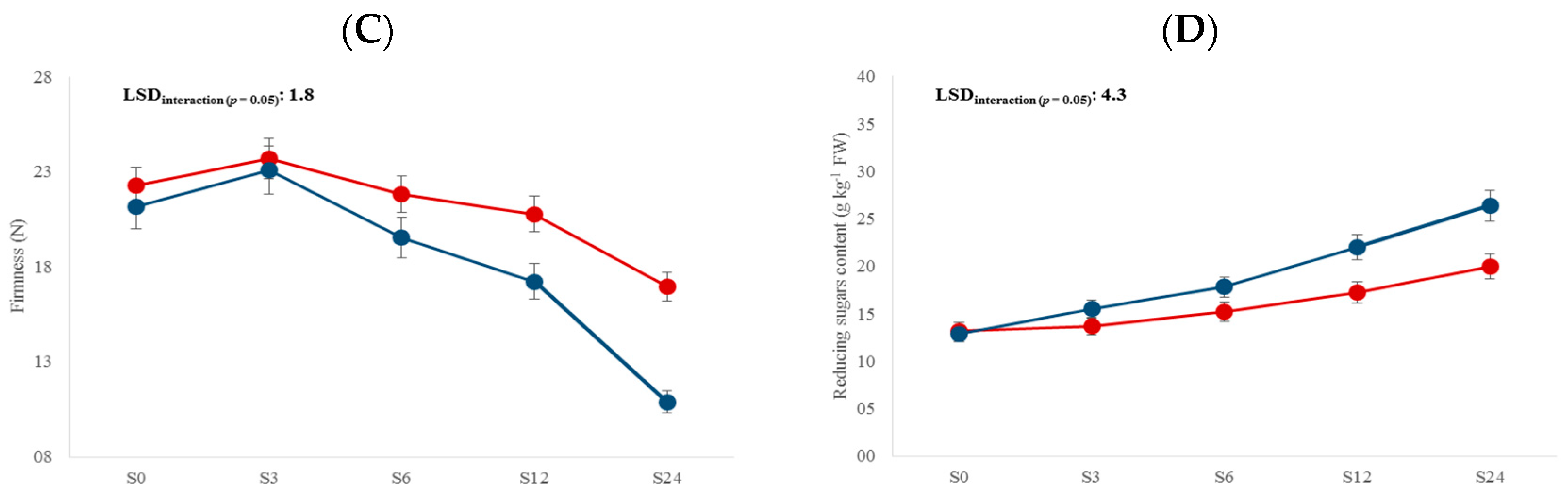
3. Results and Discussion
3.1. Physicochemical Traits
3.2. Compositional Traits
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Que, F.; Hou, X.L.; Wang, G.L.; Xu, Z.S.; Tan, G.F.; Li, T.; Wang, Y.H.; Khadr, A.; Xiong, A.S. Advances in research on the carrot, an important root vegetable in the Apiaceae family. Hortic. Res. 2019, 6, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leja, M.; Kamińska, I.; Kramer, M.; Maksylewicz-Kaul, A.; Kammerer, D.; Carle, R.; Baranski, R. The Content of Phenolic Compounds and Radical Scavenging Activity Varies with Carrot Origin and Root Color. Plant Foods Hum. Nutr. 2013, 68, 163–170. [Google Scholar] [CrossRef] [Green Version]
- Istat.it. Available online: http://dati.istat.it/Index.aspx?DataSetCode=DCSP_COLTIVAZIONI (accessed on 5 November 2020).
- Giuffrida, F.; Cassaniti, C.; Scuderi, D.; Caturano, E.; Leonardi, C. Tomato yield, nitrogen uptake and use efficiency in relation to nitrogen fertilization levels. Acta Hortic. 2012, 927, 149–154. [Google Scholar] [CrossRef]
- Reid, J.B.; Hunt, A.G.; Johnstone, P.R.; Searle, B.P.; Jesson, L.K. On the responses of carrots (Daucus carota L.) to nitrogen supply. N. Z. J. Crop Hortic. Sci. 2018, 46, 298–318. [Google Scholar] [CrossRef]
- Ierna, A.; Mauro, R.P.; Mauromicale, G. Improved yield and nutrient efficiency in two globe artichoke genotypes by balancing nitrogen and phosphorus supply. Agron. Sustain. Dev. 2012, 32, 773–780. [Google Scholar] [CrossRef]
- Prusky, D. Reduction of the incidence of postharvest quality losses, and future prospects. Food Secur. 2011, 3, 463–474. [Google Scholar] [CrossRef]
- Suojala-Ahlfors, T.; Laamanen, T.L. Effect of calcium amendment on the calcium content and storage quality of carrot (Daucus carota L.). Eur. J. Hortic. Sci. 2014, 79, 278–282. [Google Scholar]
- Larsen, H.; Wold, A.B. Effect of modified atmosphere packaging on sensory quality, chemical parameters and shelf life of carrot roots (Daucus carota L.) stored at chilled and abusive temperatures. Postharvest Biol. Technol. 2016, 114, 76–85. [Google Scholar] [CrossRef]
- Al Ubeed, H.M.S.; Wills, R.B.H.; Bowyer, M.C.; Golding, J.B. Inhibition of postharvest senescence of green leafy vegetables by exogenous D-cysteine and L-cysteine as precursors of hydrogen sulphide. J. Hortic. Sci. Biotechnol. 2019, 94, 620–626. [Google Scholar] [CrossRef]
- Mauro, R.P.; Occhipinti, A.; Longo, A.M.G.; Mauromicale, G. Effects of shading on chlorophyll content, chlorophyll fluorescence and photosynthesis of subterranean clover. J. Agron. Crop Sci. 2011, 197, 57–66. [Google Scholar] [CrossRef]
- Osservatorio Delle Acque. Available online: www.osservatorioacque.it (accessed on 26 July 2020).
- USDA United States Department of Agriculture. Soil Conservation Service. Soil Taxonomy—A Basic System of Soil Classification for Making and Interpreting Soil Surveys, 2nd ed.; Handbook, 436; U.S. Government Printing Office: Washington, DC, USA, 1999; p. 886.
- Violante, P. Metodi di Analisi Chimica del Suolo; Franco Angeli: Milano, Italy, 2000; p. 536. [Google Scholar]
- Tesi, R.; Lenzi, A. Nutrizione azotata per un’orticoltura sostenibile. Italus Hortus 2005, 12, 57–73. [Google Scholar]
- Ierna, A.; Mauromicale, G. Sustainable and profitable nitrogen fertilization management of potato. Agronomy 2019, 9, 582. [Google Scholar] [CrossRef] [Green Version]
- McGuire, R.G. Reporting of objective color measurements. HortScience 1992, 27, 1254–1255. [Google Scholar] [CrossRef] [Green Version]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. Food Sci. Technol. 2007, 28, 25–30. [Google Scholar] [CrossRef]
- Singh, G.; Kawatra, A.; Sehgal, S. Nutritional composition of selected green leafy vegetables, herbs and carrots. Plant Foods Hum. Nutr. 2001, 56, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Slinkard, K.; Singleton, V. Total phenol analysis: Automation and comparison with manual methods. Am. J. Enol. Vitic. 1977, 28, 49–55. [Google Scholar]
- Gusakov, A.V.; Kondratyeva, E.G.; Sinitsyn, A.P. Comparison of two methods for assaying reducing sugars in the determination of carbohydrase activities. Int. J. Anal. Chem. 2011, 2011. [Google Scholar] [CrossRef] [Green Version]
- Kaack, K.; Nielsen, M.; Christensen, L.P.; Thorup-Kristensen, K. Nutritionally important chemical constituents and yield of carrot (Daucus carota L.) roots grown organically using ten levels of green manure. Acta Agric. Scand. Sect. B Soil Plant Sci. 2001, 51, 125–136. [Google Scholar] [CrossRef]
- Rembiałkowska, E. Quality of plant products from organic agriculture. J. Sci. Food Agric. 2007, 87, 2757–2762. [Google Scholar] [CrossRef]
- Grinder-Pedersen, L.; Rasmussen, S.E.; Bügel, S.; Jørgensen, L.V.; Dragsted, L.O.; Gundersen, V.; Sandström, B. Effect of diets based on foods from conventional versus organic production on intake and excretion of flavonoids and markers of antioxidative defense in humans. J. Agric. Food Chem. 2003, 51, 5671–5676. [Google Scholar] [CrossRef]
- Rajasree, G.; Pillai, G.R. Effect of nitrogen nutrition on fruit quality and shelf life of cucurbitaceous vegetable bitter gourd. J. Plant Nutr. 2012, 35, 1139–1153. [Google Scholar] [CrossRef]
- Gajewski, M.; Szymczak, P.; Danilcenko, H. Changes of physical and chemical traits of roots of different carrot cultivars under cold store conditions. Veg. Crop. Res. Bull. 2010, 72, 115–127. [Google Scholar] [CrossRef]
- Rozek, S.; Leja, M.; Wojciechowska, R. Effect of differentiated nitrogen fertilization on changes of certain compounds in stored carrot roots. Folia Hortic. 2000, 12, 21–34. [Google Scholar]
- Herppich, W.B.; Mempel, H.; Geyer, M. Effects of postharvest mechanical and climatic stress on carrot tissue water relations. Postharvest Biol. Technol. 1999, 16, 43–49. [Google Scholar] [CrossRef]
- Robinson, J.E.; Browne, K.M.; Burton, W.G. Storage characteristics of some vegetables and soft fruits. Ann. Appl. Biol. 1975, 81, 399–408. [Google Scholar] [CrossRef]
- Larsen, H.; Liland, K.H. Determination of O2 and CO2 transmission rate of whole packages and single perforations in micro-perforated packages for fruit and vegetables. J. Food Eng. 2013, 119, 271–276. [Google Scholar] [CrossRef] [Green Version]
- Conte, A.; Scrocco, C.; Brescia, I.; Mastromatteo, M.; Del Nobile, M.A. Shelf life of fresh-cut Cime di rapa (Brassica rapa L.) as affected by packaging. LWT Food Sci. Technol. 2011, 44, 1218–1225. [Google Scholar] [CrossRef]
- Dawange, S.P.; Dash, S.K.; Bal, L.M.; Panda, M.K. Quality of minimally processed carrots in perforation-mediated modified-atmosphere packaging (PM-MAP). J. Food Meas. Charact. 2016, 10, 746–754. [Google Scholar] [CrossRef]
- Matimati, I.; Verboom, G.A.; Cramer, M.D. Nitrogen regulation of transpiration controls mass-flow acquisition of nutrients. J. Exp. Bot. 2014, 65, 159–168. [Google Scholar] [CrossRef] [Green Version]
- Seljåsen, R.; Kristensen, H.L.; Lauridsen, C.; Wyss, G.S.; Kretzschmar, U.; Birlouez-Aragone, I.; Kahl, J. Quality of carrots as affected by pre- and postharvest factors and processing. J. Sci. Food Agric. 2013, 93, 2611–2626. [Google Scholar] [CrossRef] [Green Version]
- Seljåsen, R.; Lea, P.; Torp, T.; Riley, H.; Berentsen, E.; Thomsen, M.; Bengtsson, G.B. Effects of genotype, soil type, year and fertilisation on sensory and morphological attributes of carrots (Daucus carota L.). J. Sci. Food Agric. 2012, 92, 1786–1799. [Google Scholar] [CrossRef] [PubMed]
- Dattner, M.; Bohn, D. Characterization of print quality in terms of colorimetric aspects. In Printing on Polymers: Fundamentals and Applications; Elsevier Inc.: Amsterdam, The Nederland, 2015; pp. 329–345. [Google Scholar]
- Sorensen, J.N. Nitrogen effects on vegetable crop production and chemical composition. Acta Hortic. 1999, 506, 41–49. [Google Scholar] [CrossRef]
- Smoleń, S.; Sady, W.; Ledwozyw-Smoleń, I.; Strzetelski, P.; Liszka-Skoczylas, M.; Rozek, S. Quality of fresh and stored carrots depending on iodine and nitrogen fertilization. Food Chem. 2014, 159, 316–322. [Google Scholar] [CrossRef]
- Ierna, A. Influence of harvest date on nitrate contents of three potato varieties for off-season production. J. Food Compos. Anal. 2009, 22, 551–555. [Google Scholar] [CrossRef]
- Sharma, K.D.; Karki, S.; Thakur, N.S.; Attri, S. Chemical composition, functional properties and processing of carrot-A review. J. Food Sci. Technol. 2012, 49, 22–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaller, R.G.; Schnitzler, W.H. Nitrogen nutrition and flavour compounds of carrots (Daucus carota L.) cultivated in Mitscherlich pots. J. Sci. Food Agric. 2000, 80, 49–56. [Google Scholar] [CrossRef]
- Gutezeit, B. Yield and nitrate content of carrots (Daucus carota L.) as affected by nitrogen supply. Acta Hortic. 1999, 506, 87–91. [Google Scholar] [CrossRef]
- Ilić, Z.S.; Šunić, L. Nitrate content of root vegetables during different storage conditions. Acta Hortic. 2015, 1079, 659–665. [Google Scholar] [CrossRef]
- Cserni, I.; Prohaszka, K.; Patocs, I. The effect of different N-doses on changes in the nitrate-sugar and carotene contents of carrot. Acta Agron. Hung. 1989, 38, 341–348. [Google Scholar]
- Tang, G.Q.; Sturm, A. Antisense repression of sucrose synthase in carrot (Daucus carota L.) affects growth rather than sucrose partitioning. Plant Mol. Biol. 1999, 41, 465–479. [Google Scholar] [CrossRef]
- Ilić, Z.S.; Šunić, L.; Milenković, L.; Pestorić, M.; Belović, M.; Kevrešan, Ž.; Mastilović, J. Nutrients content and texture changes as affected of harvest time, postharvest treatments and storage condition of carrot. Acta Sci. Pol. Hortorum Cultus 2017, 16, 63–75. [Google Scholar] [CrossRef]
- Bryant, J.P.; Stuart Chapin, F.; Klein, D.R. Carbon/nutrient balance of boreal plants in relation to vertebrate herbivory. Oikos 1983, 40, 357–369. [Google Scholar] [CrossRef] [Green Version]
- Smoleń, S.; Sady, W. The effect of various nitrogen fertilization and foliar nutrition regimes on the concentrations of sugars, carotenoids and phenolic compounds in carrot (Daucus carota L.). Sci. Hortic. 2009, 120, 315–324. [Google Scholar] [CrossRef]
- Nicolle, C.; Simon, G.; Rock, E.; Amouroux, P.; Rémésy, C. Genetic variability influences carotenoid, vitamin, phenolic, and mineral content in white, yellow, purple, orange, and dark-orange carrot cultivars. J. Am. Soc. Hortic. Sci. 2004, 129, 523–529. [Google Scholar] [CrossRef] [Green Version]
- Mareczek, A.; Wyzgolik, G.; Leja, M. Antioxidative ability of carrot as related to urea foliar application. Hort. Veg. Grow. 2004, 23, 329–335. [Google Scholar]
- Leja, M.; Rożek, S.; Mareczek, A. Effect of fertilization with reduced nitrogen forms on phenolic metabolism in carrot roots. Umbelliferae Improv. Newsl. 1998, 8, 20–22. [Google Scholar]
- Lombardo, S.; Restuccia, C.; Muratore, G.; Barbagallo, R.N.; Licciardello, F.; Pandino, G.; Scifò, G.O.; Mazzaglia, A.; Ragonese, F.; Mauromicale, G. Effect of nitrogen fertilisation on the overall quality of minimally processed globe artichoke heads. J. Sci. Food Agric. 2017, 97, 650–658. [Google Scholar] [CrossRef]
- Salveit, M.E. Physical and physiological changes in minimally processed fruits and vegetable. In Phytochemistry of Fruit and Vegetables; Tomás-Barberán, A., Ed.; Oxford University Press: New York, NY, USA, 1997; pp. 205–220. [Google Scholar]
- Ismail, A.; Zamaliah, M.M.; Chin, W.F. Total antioxidant activity and phenolic content in selected vegetables. Food Chem. 2004, 87, 581–586. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Df | CWL (g kg−1 FW) | Root DM Content (%) | Firmness (N) | ΔE*ab | Nitrate Content (mg kg−1 FW) | RSc Content (g kg−1 FW) | TPC (mg GAE kg−1 FW) | TEAC (mg Trolox kg−1 FW) | |
|---|---|---|---|---|---|---|---|---|---|
| Nitrogen rate (N) | 1 | 162.2 *** | 8.8 *** | 5.0 * | 323.8 *** | 103.3 *** | NS | 51.4 *** | 5.3 * |
| Leaf presence (L) | 1 | 50.4 *** | NS | 62.2 *** | NS | 20.0 *** | 13.8 *** | NS | NS |
| Storage time (S) | 4 | 65.6 *** | NS | 66.9 *** | 52.7 *** | 3.5 * | 26.1 *** | 49.1 *** | 105.6 *** |
| N × L | 1 | 28.1 *** | NS | NS | NS | NS | NS | NS | NS |
| N × S | 4 | 25.8 *** | NS | 6.7 *** | NS | NS | NS | 3.1 * | 19.6 *** |
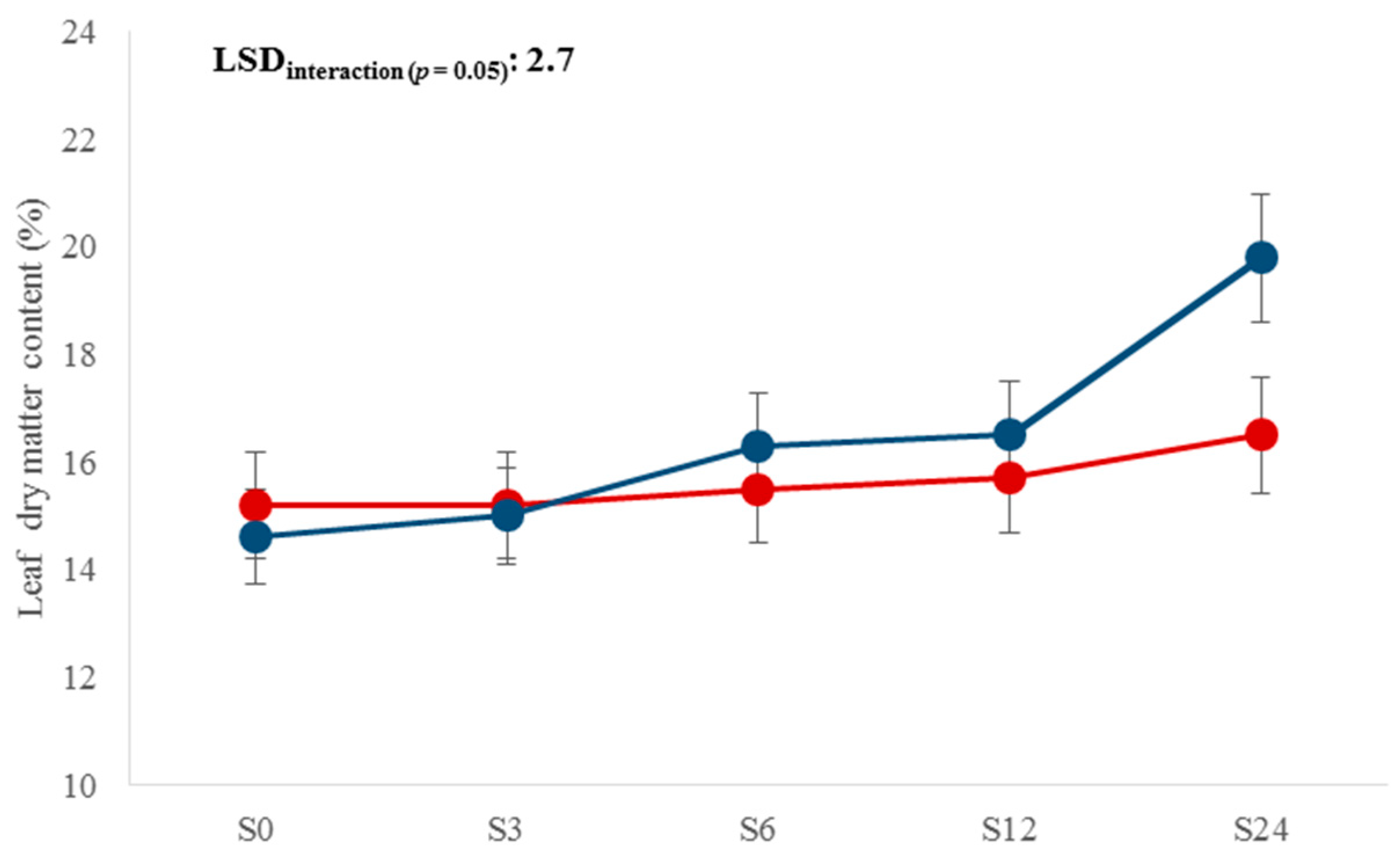
| L × S | 4 | 11.3 *** | 4.9 *** | 5.1 ** | NS | NS | 4.4 ** | NS | NS |
| N × L × S | 4 | NS | NS | NS | NS | NS | NS | NS | NS |
| CWL (g kg−1 FW) | Leaf DM Content (%) | Root DM Content (%) | Firmness (N) | ΔE*ab | ||
|---|---|---|---|---|---|---|
| N fertilization | N120 | 23.0 b | 15.6 b | 11.7 b | 19.3 b | 0.38 b |
| N240 | 66.5 a | 16.4 a | 12.2 a | 20.2 a | 0.86 a | |
| Leaf presence | leaf– | 32.6 b | - | 11.8 a | 21.3 a | 0.63 a |
| leaf+ | 56.9 a | - | 12.4 a | 18.2 b | 0.62 a | |
| Storage time | S0 | - | 14.9 d | 11.7 a | 21.7 b | - |
| S3 | 15.1 d | 15.1 cd | 11.9 a | 23.4 a | 0.38 d | |
| S6 | 28.5 c | 15.9 bc | 11.8 a | 20.7 b | 0.55 c | |
| S12 | 60.7 b | 16.1 b | 12.1 a | 19.0 c | 0.74 b | |
| S24 | 74.7 a | 18.2 a | 12.7 a | 13.9 d | 0.81 a |
| Nitrate Content (mg kg−1 FW) | RSc Content (g kg−1 FW) | TPC (mg GAE kg−1 FW) | TEAC (mg Trolox kg−1 FW) | ||
|---|---|---|---|---|---|
| N fertilization | N120 | 54.9 b | 17.4 a | 512 a | 194 a |
| N240 | 105.9 a | 17.3 a | 440 b | 181 b | |
| Leaf presence | leaf− | 91.6 a | 16.0 b | 485 a | 199 a |
| leaf+ | 69.2 b | 18.7 a | 467 a | 176 a | |
| Storage time | S0 | 91.8 a | 13.0 e | 395 e | 93 d |
| S3 | 89.7 a | 14.5 d | 424 d | 175 c | |
| S6 | 78.1 b | 16.5 c | 472 c | 181 c | |
| S12 | 76.5 b | 19.6 b | 490 b | 214 b | |
| S24 | 65.9 c | 23.2 a | 600 a | 272 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ierna, A.; Mauro, R.P.; Leonardi, C.; Giuffrida, F. Shelf-Life of Bunched Carrots as Affected by Nitrogen Fertilization and Leaf Presence. Agronomy 2020, 10, 1982. https://doi.org/10.3390/agronomy10121982
Ierna A, Mauro RP, Leonardi C, Giuffrida F. Shelf-Life of Bunched Carrots as Affected by Nitrogen Fertilization and Leaf Presence. Agronomy. 2020; 10(12):1982. https://doi.org/10.3390/agronomy10121982
Chicago/Turabian StyleIerna, Anita, Rosario Paolo Mauro, Cherubino Leonardi, and Francesco Giuffrida. 2020. "Shelf-Life of Bunched Carrots as Affected by Nitrogen Fertilization and Leaf Presence" Agronomy 10, no. 12: 1982. https://doi.org/10.3390/agronomy10121982