Orchard Planting Density and Tree Development Stage Affects Physiological Processes of Apple (Malus domestica Borkh.) Tree



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growing Conditions
2.2. Measurements
2.3. Biochemical Analyses
2.3.1. Determination of Chlorophylls (a and b) and Carotenes (α and β) by High-Performance Liquid Chromatography (HPLC)
2.3.2. Determination of Soluble Sugars Byultra-Performance Liquid Chromatography (UPLC)
2.3.3. Determination of Total Starch by Spectrophotometric Method
2.4. Statistical Analysis
2.5. Meteorological Conditions
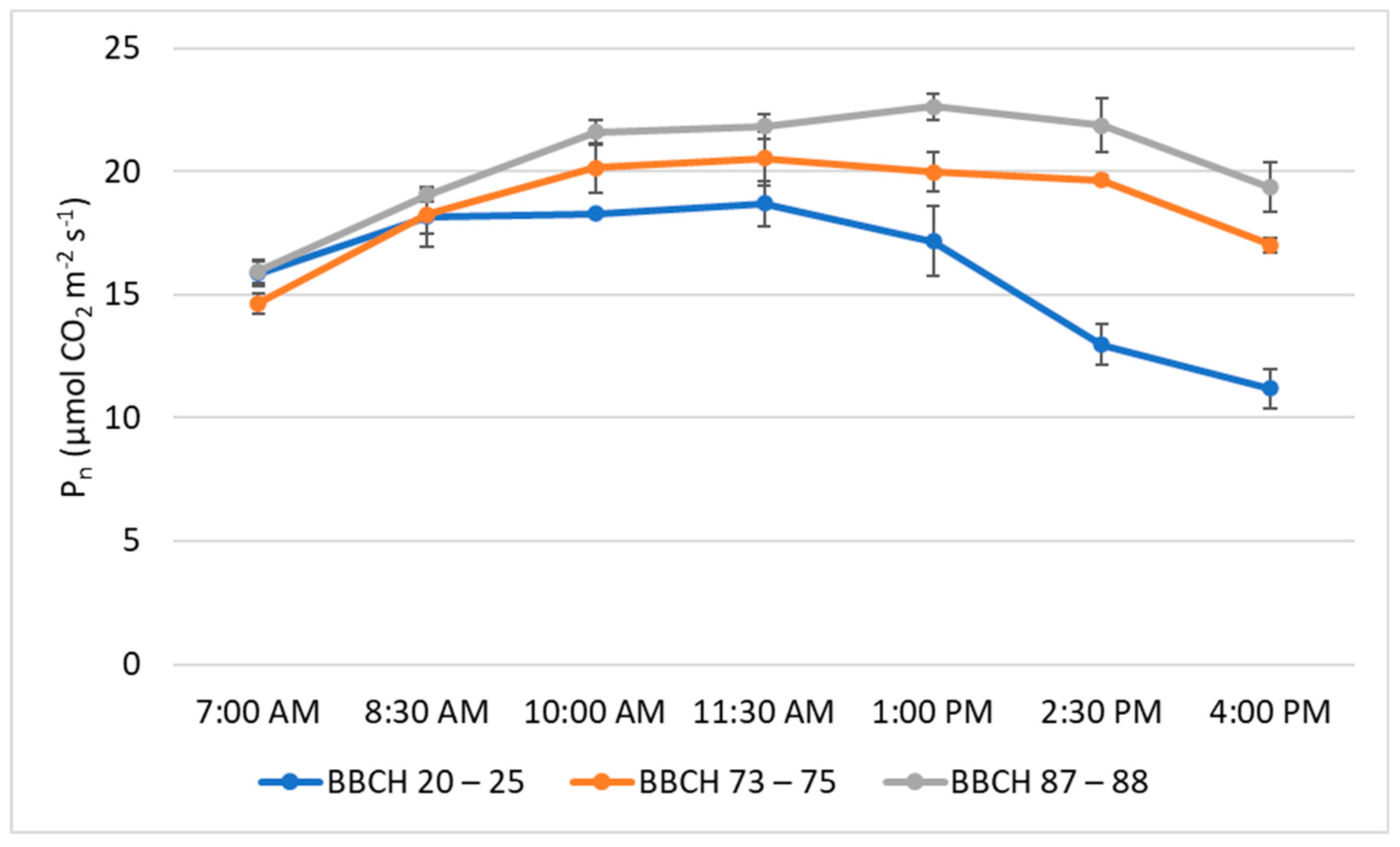
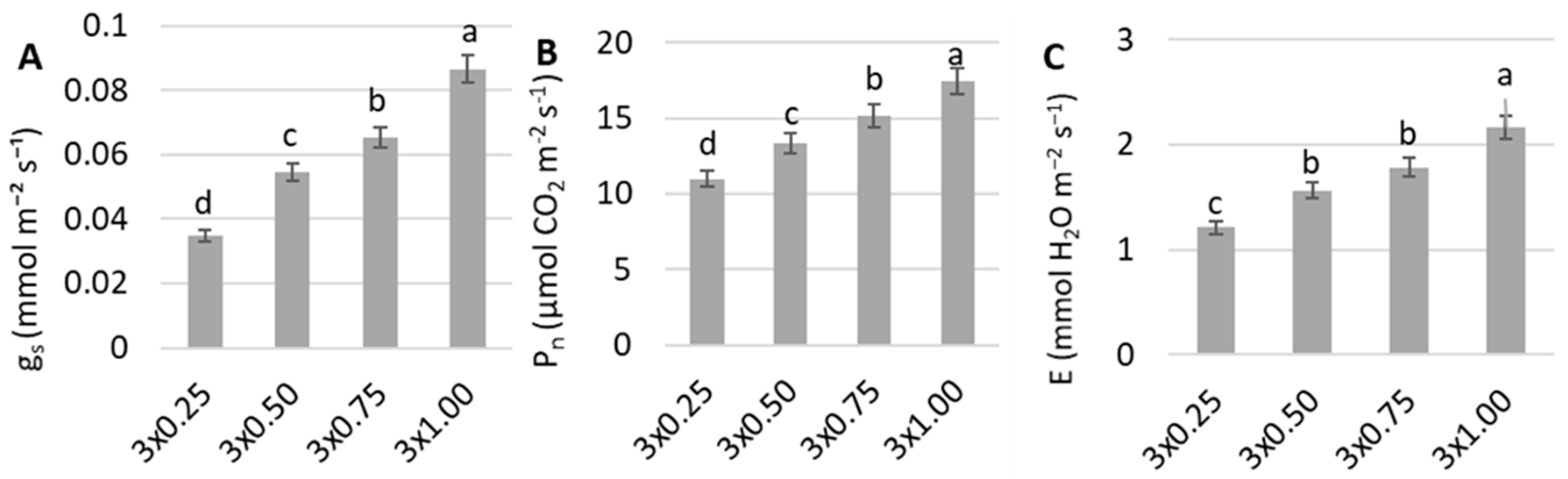
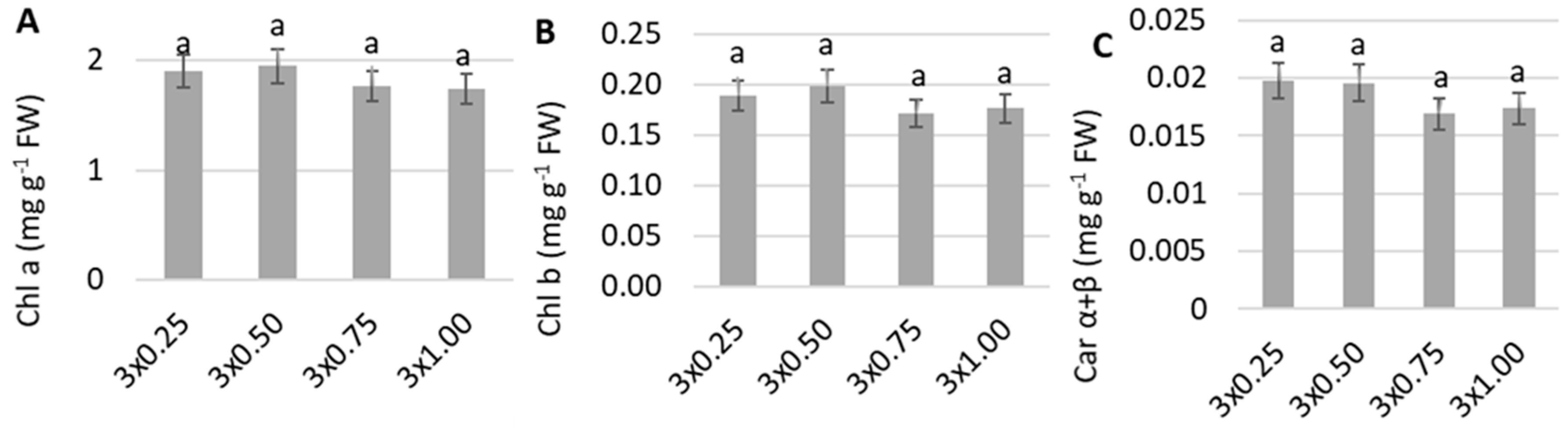
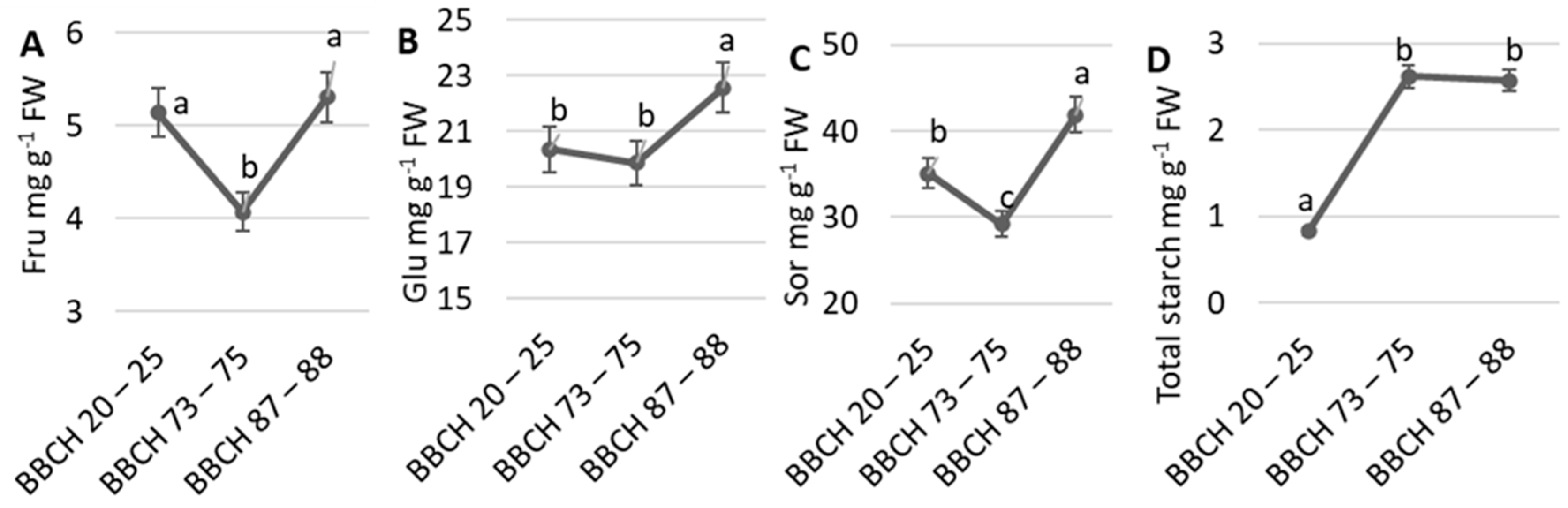
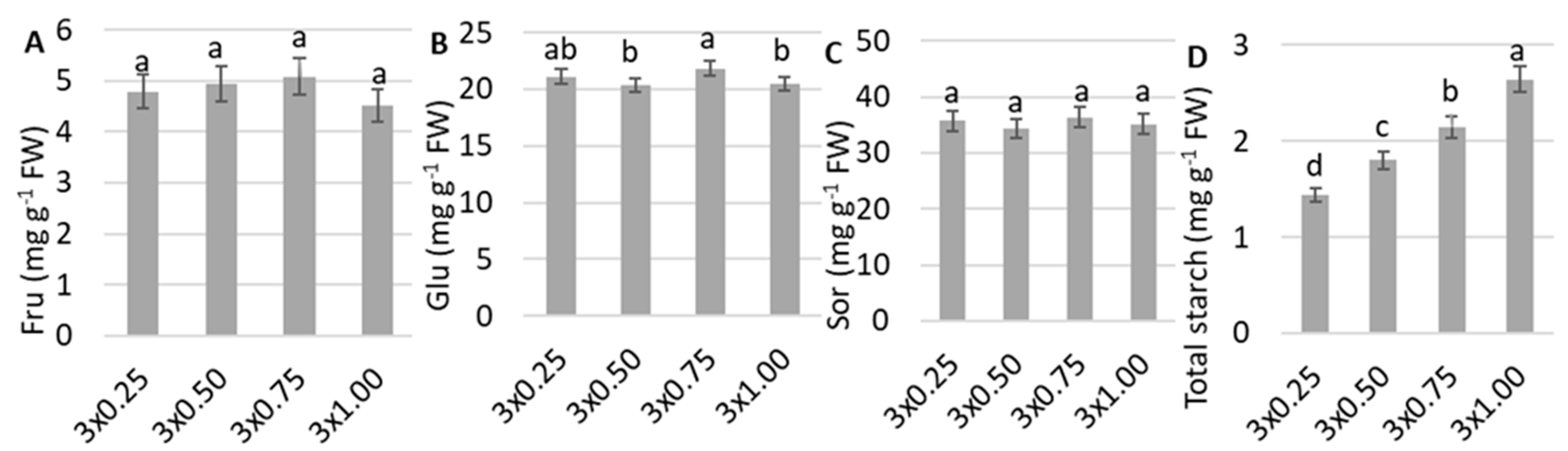
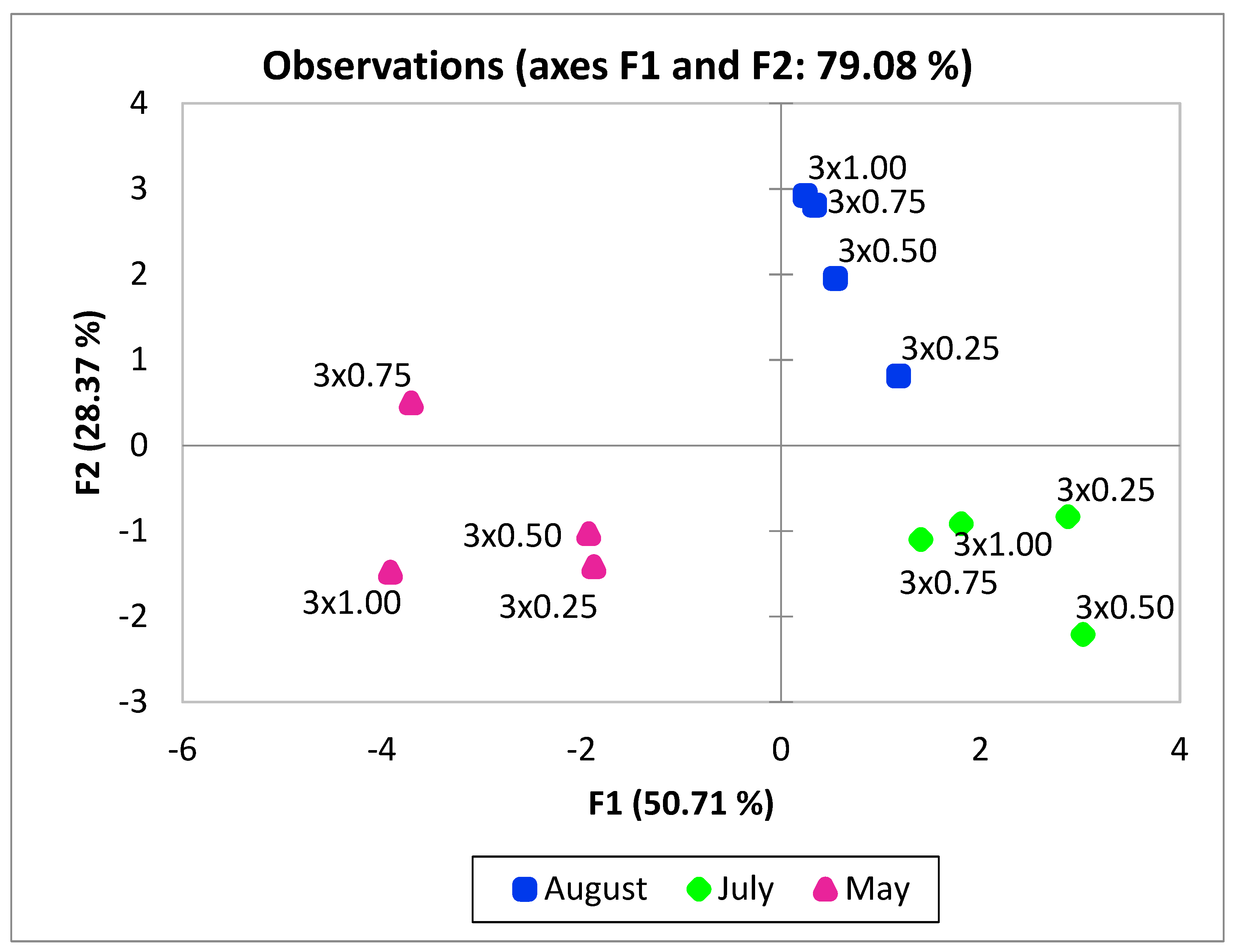
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Thorne, H.C.; Jones, K.H.; Peters, S.P.; Archer, S.N.; Dijk, D.-J. Daily and Seasonal Variation in the Spectral Composition of Light Exposure in Humans. Chrono Int. 2009, 26, 854–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Zhang, J.; Li, C.; Zhang, Z.; Ma, F.; Li, M. Response of sugar metabolism in apple leaves subjected to short-term drought stress. Plant Physiol. Biochem. 2019, 141, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, A.Q.; Pandit, A.K.; Ganai, B.A. Seasonal variation in chlorophyll content of some selected plant species of Yousmarg grassland ecosystem. Asian J. Plant Sci. Res. 2017, 7, 33–36. [Google Scholar]
- Marron, N.; Brignolas, F.; Delmotte, F.M.; Dreyer, E. Modulation of leaf physiology by age and in response to abiotic constraints in young cuttings of two Populus deltoides times P. nigra genotypes. Ann. For. Sci. 2008, 65, 1. [Google Scholar] [CrossRef]
- Rodríguez-Calcerrada, J.; Limousin, J.-M.; Martin-StPaul, N.K.; Jaeger, C.; Rambal, S. Gas exchange and leaf aging in an evergreen oak: Causes and consequences for leaf carbon balance and canopy respiration. Tree Physiol. 2012, 32, 464–477. [Google Scholar] [CrossRef] [PubMed]
- Bielczynski, L.W.; Łącki, M.K.; Hoefnagels, I.; Gambin, A.; Croce, R. Leaf and Plant Age Affects Photosynthetic Performance and Photoprotective Capacity. Plant Physiol. 2017, 175, 1634–1648. [Google Scholar] [CrossRef]
- Croce, R.; Van Amerongen, H. Light-harvesting and structural organization of Photosystem II: From individual complexes to thylakoid membrane. J. Photochem. Photobiol. B Biol. 2011, 104, 142–153. [Google Scholar] [CrossRef] [Green Version]
- Esteban, R.; Barrutia, O.; Artetxe, U.; Fernández-Marín, B.; Hernández, A.; García-Plazaola, J.I. Internal and external factors affecting photosynthetic pigment composition in plants: A meta-analytical approach. New Phytol. 2015, 206, 268–280. [Google Scholar] [CrossRef]
- Cazzaniga, S.; Li, Z.; Niyogi, K.K.; Bassi, R.; Dall’Osto, L. The Arabidopsis szl1 mutant reveals a critical role of β-carotene in photosystem I photoprotection. Plant Physiol. 2012, 159, 1745–1758. [Google Scholar] [CrossRef] [Green Version]
- Croce, R.; Van Amerongen, H. Light-harvesting in photosystem I. Photosynth. Res. 2013, 116, 153–166. [Google Scholar] [CrossRef] [Green Version]
- Rattan, K. Comparative analyses of physiological assays and chlorophyll a variable fluorescence parameters: Investigating the importance of phosphorus availability in oligotrophic and eutrophic freshwater systems. Aquat. Ecol. 2017, 51, 359–375. [Google Scholar] [CrossRef]
- Young, A.J.; Britton, G. The distribution of α-carotene in the photosynthetic pigment-protein complexes of higher plants. Plant Sci. 1989, 64, 179–183. [Google Scholar] [CrossRef]
- Morganti, P. The photoprotective activity of nutraceuticals. Clin. Dermatol. 2009, 27, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Niyogi, K.K.; Björkman, O.; Grossman, A.R. The roles of specific xanthophylls in photoprotection. Proc. Natl. Acad. Sci. USA 1997, 94, 14162–14167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holt, N.E.; Zigmantas, D.; Valkunas, L.; Li, X.P.; Niyogi, K.K.; Fleming, G.R. Carotenoid cation formation and the regulation of photosynthetic light harvesting. Science 2005, 307, 433–436. [Google Scholar] [CrossRef] [Green Version]
- Rafieian-Kopaei, M.; Baradaran, A.; Rafieian, M. Plants antioxidants: From laboratory to clinic. J. Nephropathol. 2013, 2, 152–153. [Google Scholar] [CrossRef]
- Leisso, R.; Buchanan, D.A.; Lee, J.; Mattheis, J.P.; Sater, C.; Hanrahan, I.; Watkins, C.B.; Gapper, N.; Johnston, J.W.; Schaffer, R.J.; et al. Chilling-related cell damage of apple (Malus & times; domesticaBorkh.) fruit cortical tissue impacts antioxidant, lipid and phenolic metabolism. Physiol. Plant. 2014, 153, 204–220. [Google Scholar] [CrossRef]
- Rosa, M.; Prado, C.; Podazza, G.; Interdonato, R.; González, J.A.; Hilal, M.; Prado, F.E. Soluble sugars: Metabolism, sensing and abiotic stress: A complex network in the life of plants. Plant Signal. Behav. 2009, 4, 388–393. [Google Scholar] [CrossRef] [Green Version]
- Ruan, Y.-L. Sucrose Metabolism: Gateway to Diverse Carbon Use and Sugar Signaling. Annu. Rev. Plant Biol. 2014, 65, 33–67. [Google Scholar] [CrossRef]
- Cheng, L.; Zhou, R.; Reidel, E.J.; Sharkey, T.D.; Dandekar, A.M. Antisense inhibition of sorbitol synthesis leads to up-regulation of starch synthesis without altering CO2 assimilation in apple leaves. Planta 2005, 220, 767–776. [Google Scholar] [CrossRef]
- Escobar-Gutierrez, A.J. Gaudillere, J.P. Carbon partitioning in source leaves of peach, a sorbitol synthesizing species, is modified by photosynthetic rate. Physiol. Plant 2007, 100, 353–360. [Google Scholar] [CrossRef]
- Filip, M.; Vlassa, M.; Coman, V.; Halmagyi, A. Simultaneous determination of glucose, fructose, sucrose and sorbitol in the leaf and fruit peel of different apple cultivars by the HPLC & ndash; RI optimized method. Food Chem. 2016, 199, 653–659. [Google Scholar] [CrossRef] [PubMed]
- Dai, M.; Shi, Z.; Xu, C.-J. Genome-Wide Analysis of Sorbitol Dehydrogenase (SDH) Genes and Their Differential Expression in Two Sand Pear (Pyrus pyrifolia) Fruits. Int. J. Mol. Sci. 2015, 16, 13065–13083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, D.; Li, C.-L.; Park, H.-J.; González, J.; Wang, J.; Dandekar, A.M.; Turgeon, B.G.; Cheng, L. Sorbitol Modulates Resistance to Alternaria alternata by Regulating the Expression of an NLR Resistance Gene in Apple. Plant Cell 2018, 30, 1562–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, T.; Cai, Y.; Yang, Q.; Ogutu, C.O.; Liao, L.; Han, Y. Analysis of sorbitol content variation in wild and cultivated apples. J. Sci. Food Agric. 2020, 100, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, W.; Wang, Z.; Ma, L.; Guo, Y.; Ren, X.; Mei, L. Influence of shading on photosynthesis and antioxidative activities of enzymes in apple trees. Photosynthetica 2019, 57, 857–865. [Google Scholar] [CrossRef] [Green Version]
- Pagter, M.; Andersen, U.B.; Andersen, L. Winter warming delays dormancy release, advances budburst, alters carbohydrate metabolism and reduces yield in a temperate shrub. AoB Plants 2015, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- Nii, N. Changes of Starch and Sorbitol in Leaves Before and After Removal of Fruits from Peach Trees. Ann. Bot. 1997, 79, 139–144. [Google Scholar] [CrossRef] [Green Version]
- Patrick, J.W.; Offler, C.E. Compartmentation of transport and transfer events in developing seeds. J. Exp. Bot. 2001, 52, 551–564. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, J.; Yang, C.; Du, S.; Yang, W. Photosynthetic performance of soybean plants to water deficit under high and low light intensity. South Afr. J. Bot. 2016, 105, 279–287. [Google Scholar] [CrossRef]
- Li, Y.; Li, H.; Li, Y.; Zhang, S. Improving water-use efficiency by decreasing stomatal conductance and transpiration rate to maintain higher ear photosynthetic rate in drought-resistant wheat. Crop. J. 2017, 5, 231–239. [Google Scholar] [CrossRef]
- Lopez, G.; Boini, A.; Manfrini, L.; Torres-Ruiz, J.; Pierpaoli, E.; Zibordi, M.; Losciale, P.; Morandi, B.; Corelli-Grappadelli, L. Effect of shading and water stress on light interception, physiology and yield of apple trees. Agric. Water Manag. 2018, 210, 140–148. [Google Scholar] [CrossRef]
- Berry, Z.C.; Goldsmith, G. Diffuse light and wetting differentially affect tropical tree leaf photosynthesis. New Phytol. 2019, 225, 143–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kairiūkštis, L.; Malinauskas, A. The influence of the initial density on spruce (Picea abies Karsten) wood quality. Balt. For. 2001, 7, 8–17. [Google Scholar]
- Kuliešis, A.; Saladis, J.; Kuliešis, A.A. Development and productivity of young scots pine stands by regulating density. Balt. For. 2010, 16, 235–246. [Google Scholar]
- Castedo-Dorado, F.; Crecente-Campo, F.; Alvarez-Alvarez, P.; Barrio-Anta, M. Development of a stand density management diagram for radiata pine stands including assessment of stand stability. Forestry 2009, 82, 1–16. [Google Scholar] [CrossRef]
- Tombesi, S.; Nardini, A.; Frioni, T.; Soccolini, M.; Zadra, C.; Farinelli, D.; Poni, S.; Palliotti, A. Stomatal closure is induced by hydraulic signals and maintained by ABA in drought-stressed grapevine. Sci. Rep. 2015, 5, 12449. [Google Scholar] [CrossRef]
- Wang, Z.; Li, G.; Sun, H.; Ma, L.; Guo, Y.; Zhao, Z.; Gao, H.; Mei, L. Effects of drought stress on photosynthesis and photosynthetic electron transport chain in young apple tree leaves. Biol. Open 2018, 7, bio035279. [Google Scholar] [CrossRef] [Green Version]
- Bhusal, N.; Han, S.-G.; Yoon, T.-M. Impact of drought stress on photosynthetic response, leaf water potential, and stem sap flow in two cultivars of bi-leader apple trees (Malus & times; domestica Borkh.). Sci. Hortic. 2019, 246, 535–543. [Google Scholar] [CrossRef]
- Kviklys, D.; Kviklienė, N.; Bielicki, P.; Bite, A.; Lepsis, J.; Univer, T.; Univer, N.; Uselis, N.; Lanauskas, J. Baltic fruit rootstock studies: Evaluation of apple (Malus domestica Borkh.) new rootstocks. Zemdirbyste-Agriculture 2013, 100, 441–446. [Google Scholar] [CrossRef]
- Samuolienė, G.; Viškelienė, A.; Sirtautas, R.; Kviklys, D. Relationships between apple tree rootstock, crop-load, plant nutritional status and yield. Sci. Hortic. 2016, 211, 167–173. [Google Scholar] [CrossRef]
- Kviklys, D.; Lanauskas, J.; Uselis, N.; Viškelis, J.; Viškelienė, A.; Buskienė, L. Samuolienė, G. Rootstock vigour and leaf colour affect apple tree nutrition. Zemdirb. Agric. 2017, 104, 185–190. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.; Sun, Z.; Chen, C.; Zhang, L.; Zhu, S. Simultaneous separation and determination of fructose, sorbitol, glucose and sucrose in fruits by HPLC & ndash;ELSD. Food Chem. 2014, 145, 784–788. [Google Scholar] [CrossRef] [PubMed]
- Brons, C.; Olieman, C. Study of the high-performance liquid chromatographic separation of reducing sugars, applied to the determination of lactose in milk. J. Chromatogr. A 1983, 259, 79–86. [Google Scholar] [CrossRef]
- Greer, D.H. Limitations to photosynthesis of leaves of apple (Malus domestica) trees across the growing season prior to and after harvest. Photosynthetica 2019, 57, 483–490. [Google Scholar] [CrossRef] [Green Version]
- Susanti, R.A.; Hadley, P.; Daymond, A.J.; Bastide, P.; Lambert, S.; Ingram, K.; Motamayor, J.C. The effect of pruning on photosynthetic rate of cacao tree in a novel cropping system. In Proceedings of the 2017 International Symposium on Cocoa Research (ISCR), Lima, Peru, 13 November 2017. [Google Scholar]
- Laužikė, K.; Sirgedaitė-Šėžienė, V.; Uselis, N.; Samuolienė, G. The Impact of Stress Caused by Light Penetration and Agrotechnological Tools on Photosynthetic Behavior of Apple Trees. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef]
- Zeng, G.; Guo-Yun, W.; Xu, J.; Hu, M.; Zheng, J.; Wu, Z. Partial shade optimizes photosynthesis and growth in bayberry (Myrica rubra) trees. Hortic. Environ. Biotechnol. 2017, 58, 203–211. [Google Scholar] [CrossRef]
- Charbonnier, F.; Roupsard, O.; Le Maire, G.; Guillemot, J.; Casanoves, F.; Lacointe, A.; Vaast, P.; Allinne, C.; Audebert, L.; Cambou, A.; et al. Increased light-use efficiency sustains net primary productivity of shaded coffee plants in agroforestry system. Plant Cell Environ. 2017, 40, 1592–1608. [Google Scholar] [CrossRef]
- Muhidin, M.; Syam’Un, E.; Kaimuddin, M.; Musa, Y.; Sadimantara, G.R.; Usman, M.; Leomo, S.; Rakian, T.C. The effect of shade on chlorophyll and anthocyanin content of upland red rice. IOP Conf. Ser. Earth Environ. Sci. 2018, 122, 012030. [Google Scholar] [CrossRef]
- Chen, H.; Li, Q.-P.; Zeng, Y.-L.; Deng, F.; Ren, W.-J. Effect of different shading materials on grain yield and quality of rice. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Furze, M.; Huggett, B.A.; Aubrecht, D.M.; Stolz, C.D.; Carbone, M.S.; Richardson, A.D. Whole-tree nonstructural carbohydrate storage and seasonal dynamics in five temperate species. New Phytol. 2019, 221, 1466–1477. [Google Scholar] [CrossRef] [PubMed]
- Mesa, K.; Serra, S.; Masia, A.; Gagliardi, F.; Bucci, D.; Musacchi, S. Seasonal trends of starch and soluble carbohydrates in fruits and leaves of ‘Abbé Fétel’ pear trees and their relationship to fruit quality parameters. Sci. Hortic. 2016, 211, 60–69. [Google Scholar] [CrossRef]
- Berüter, J.; Feusi, M.E.S.; Rüedi, P. Sorbitol and sucrose partitioning in the growing apple fruit. J. Plant Physiol. 1997, 151, 269–276. [Google Scholar] [CrossRef]
- Teo, G.; Suzuki, Y.; Uratsu, S.L.; Lampinen, B.; Ormonde, N.; Hu, W.K.; DeJong, T.M.; Dandekar, A.M. Silencing leaf sorbitol synthesis alters long-distance partitioning and apple fruit quality. Proc. Natl. Acad. Sci. USA 2006, 103, 18842–18847. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Lei, H.; Zhao, X.; Tian, R.; Li, T. Characterization of Three Sorbitol Transporter Genes in Micropropagated Apple Plants Grown under Drought Stress. Plant Mol. Biol. Rep. 2011, 30, 123–130. [Google Scholar] [CrossRef]
- Oren, I.; Mannerheim, N.; Dumbur, R.; Fangmeier, A.; Buchmann, N.; Grünzweig, J.M. Patterns and dynamics of canopy–root coupling in tropical tree saplings vary with light intensity but not with root depth. New Phytol. 2019, 225, 727–739. [Google Scholar] [CrossRef]
- Hampson, C.R.; Quamme, H.A.; Kappel, F.; Brownlee, R.T. Varying Density with Constant Rectangularity: II. Effects on Apple Tree Yield, Fruit Size, and Fruit Color Development in Three Training Systems over Ten Years. Hortic. Sci. 2004, 39, 507–511. [Google Scholar] [CrossRef] [Green Version]
- Šabajevienė, G.; Kviklys, D.; Duchovskis, P. Rootstock effect on photosynthetic pigment system formation in leaves of apple cv.‘Auksis’. Sodininkystė ir Daržininkystė 2006, 25, 357. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Distances between trees | 3 × 0.25 m | 3 × 0.50 m | 3 × 0.75 m | 3 × 1.00 m |
| Tree high | 2.5 ± 0.05 m | 2.5 ± 0.05 m | 2.5 ± 0.05 m | 2.5 ± 0.05 m |
| Canopy high | 2.3 ± 0.08 m | 2.3 ± 0.08 m | 2.3 ± 0.08 m | 2.3 ± 0.08 m |
| Crown radius | 0.50 ± 0.03 m | 0.81 ± 0.04 m | 0.94 ± 0.04 m | 1.13 ± 0.06 m |
| January | February | March | April | May | June | July | August | September | October | November | December | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Multiannual | −5.1 | −4.6 | −0.7 | 5.4 | 11.9 | 15.4 | 16.7 | 16.2 | 11.9 | 7.2 | 2.0 | −2.4 |
| 2018 | −1.4 | −6.5 | −1.7 | 10.6 | 16.7 | 17.7 | 20.3 | 19.2 | 14.5 | 8.0 | 3.6 | −1.0 |
| 2019 | −4.6 | 1.8 | 5.2 | 9.1 | 12.5 | 17.8 | 17.4 | 21.0 | 11.7 | 8.2 | 3.4 | 1.6 |
| January | February | March | April | May | June | July | August | September | October | November | December | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Multiannual | 52.0 | 30.0 | 37.0 | 42.0 | 52.0 | 68.0 | 79.0 | 76.0 | 68.0 | 60.0 | 65.0 | 56.0 |
| 2018 | 55.0 | 8.6 | 14.8 | 66.8 | 18.2 | 22.8 | 117.8 | 86.2 | 17.8 | 39.4 | 13.8 | 32.4 |
| 2019 | 40.0 | 50.8 | 45.8 | 4.6 | 43.4 | 37.0 | 121.2 | 207.2 | 89.6 | 85.4 | 24.4 | 30.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laužikė, K.; Uselis, N.; Kviklys, D.; Samuolienė, G. Orchard Planting Density and Tree Development Stage Affects Physiological Processes of Apple (Malus domestica Borkh.) Tree. Agronomy 2020, 10, 1912. https://doi.org/10.3390/agronomy10121912
Laužikė K, Uselis N, Kviklys D, Samuolienė G. Orchard Planting Density and Tree Development Stage Affects Physiological Processes of Apple (Malus domestica Borkh.) Tree. Agronomy. 2020; 10(12):1912. https://doi.org/10.3390/agronomy10121912
Chicago/Turabian StyleLaužikė, Kristina, Nobertas Uselis, Darius Kviklys, and Giedrė Samuolienė. 2020. "Orchard Planting Density and Tree Development Stage Affects Physiological Processes of Apple (Malus domestica Borkh.) Tree" Agronomy 10, no. 12: 1912. https://doi.org/10.3390/agronomy10121912