Rootstock Influence on Growth and Mineral Content of Citrus limon and Citrus sinensis cv. Valencia Inoculated with Candidatus Liberibacter Asiaticus


 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Bacterial Inoculation of Plant Materials
2.3. Bacteria Identification
2.4. Test Parameters and Data Analysis
2.4.1. Test Parameters
2.4.2. Mineral Content
2.4.3. Data Analysis
3. Results
3.1. Bacteria Identification
3.2. Plant Height and Stem Diameter
3.3. Mineral Concentration of Citrus limon and Citrus sinensis in Uninfected and Infected Plants
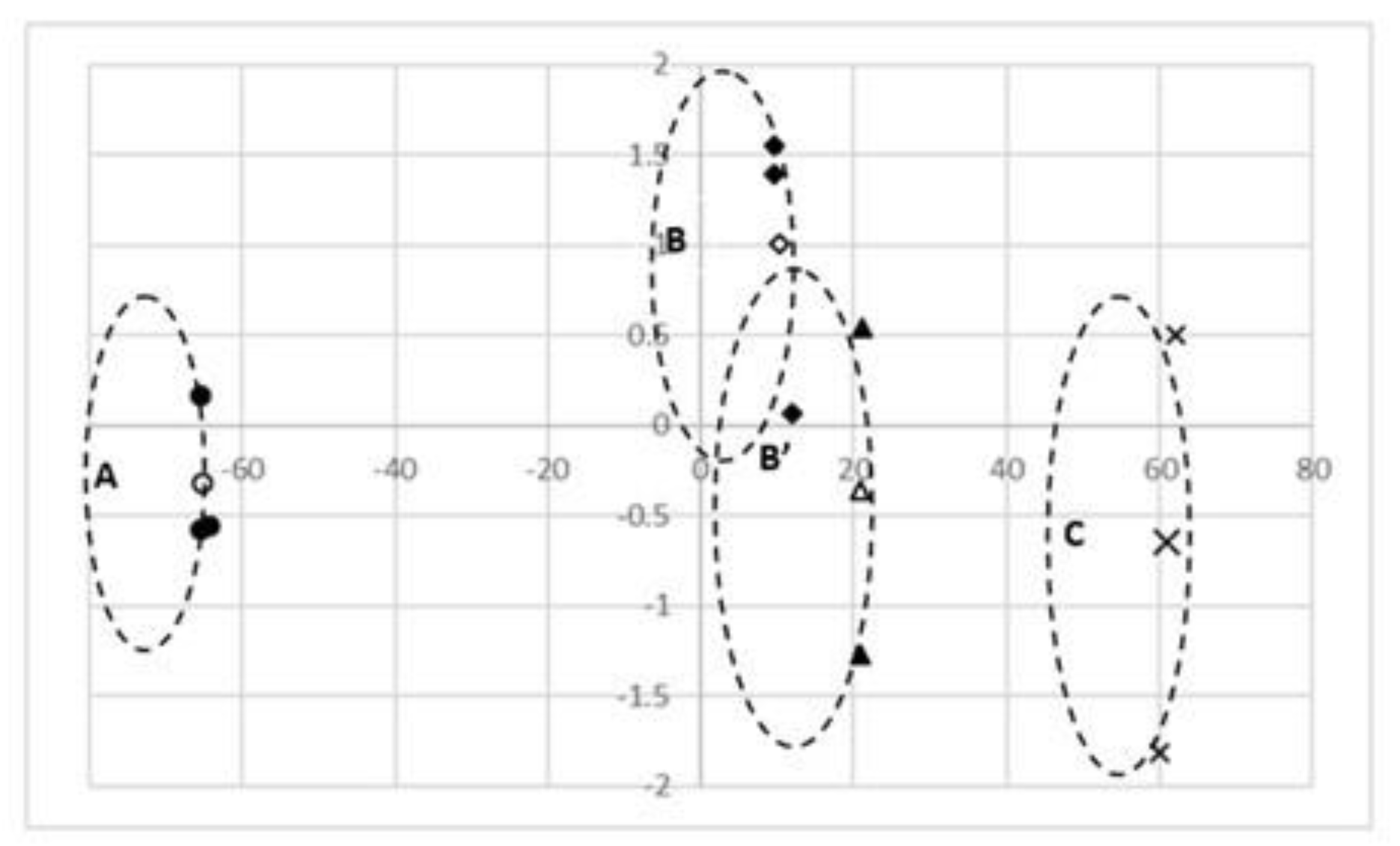
3.4. Standardized Canonical (Can) Nutrient Coefficients, Plant Height, and Rootstock Stem Diameter
3.5. Significance of the Mahalanobis Distances between Each of the Groups and Canonical Dispersions of Uninfected and Infected Species
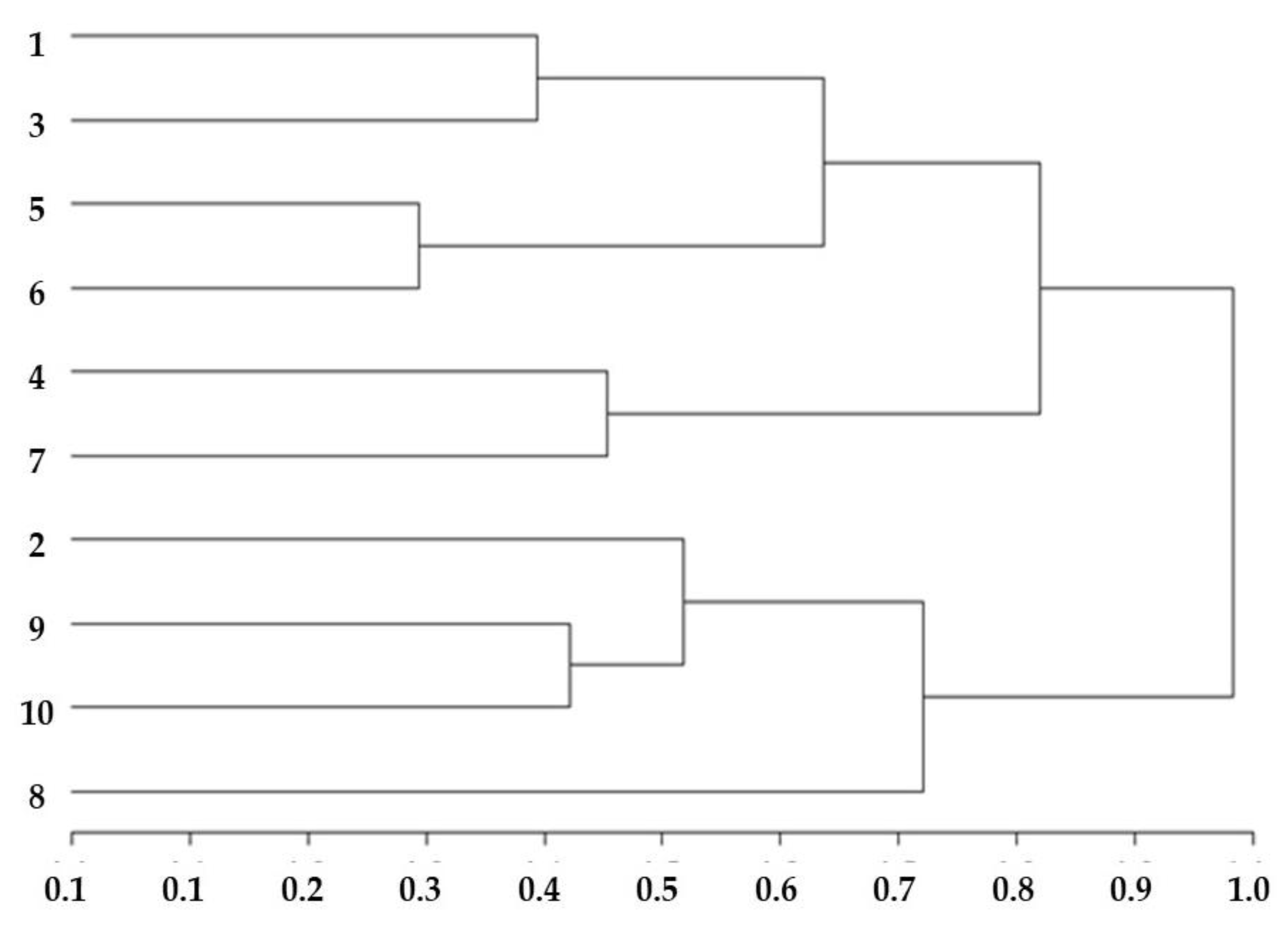
3.6. Dendrogram of Mineral Content in Citrus limon and Citrus sinensis (Graft/Rootstock) Treatments
4. Discussion
4.1. Detection of Ca. Liberibacter Asiaticus
4.2. Plant Height and Stem Diameter
4.3. Mineral Content of Citrus limon and Citrus sinensis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wang, N.; Pierson, E.A.; Setubal, J.A.; Xu, J.; Levy, J.G.; Zhang, Y.; Li, J.; Rangel, L.T.; Martins, J. The Candidatus Liberibacter–host interface: Insights into pathogenesis mechanisms and disease control. Ann. Rev. Phytopathol. 2017, 55, 451–482. [Google Scholar] [CrossRef]
- Robles-González, M.M.; Orozco-Santos, M.; Manzanilla-Ramírez, M.A.; Velázquez-Monreal, J.J.; Medina-Urrutia, V.M.; Sanches-Stuchi, E. Experiencias con Huanglongbing en limón mexicano, en el estado de Colima, México. Citrus Res. Technol. 2018, 39, 2–12. [Google Scholar] [CrossRef] [Green Version]
- Rao, M.J.; Ding, F.; Wang, N.; Deng, X. Metabolic mechanisms of host species against citrus Huanglongbing (Greening Disease). Crit. Rev. Plant Sci. 2019, 37, 1–16. [Google Scholar] [CrossRef]
- Salcedo-Baca, D.; Hinojosa, R.A.; Mora-Aguilera, G.; Covarrubias-Gutierrez, I.; DePaolis, F.J.; Mora-Flores, J.S.; Cíntora, C. Evaluación del impacto económico de la enfermedad de los cítricos Huanglongbing (hlb) en la cadena citrícola mexicana. IICA, SAGARPA, SENASICA. México. 2011, pp. 40–47. Available online: http://repiica.iica.int/docs/b2146e/b2146e.pdf (accessed on 20 June 2020).
- Hall, D.G. Incidence of “Candidatus Liberibacter asiaticus” in a Florida population of Asian citrus psyllid. J. Appl. Entomol. 2018, 142, 97–103. [Google Scholar] [CrossRef]
- Johnson, E.G.; Wu, J.; Bright, D.B.; Graham, J.H. Association of ‘Candidatus Liberibacter asiaticus’ root infection, but not phloem plugging with root loss on huanglongbing-affected trees prior to appearance of foliar symptoms. Plant Pathol. 2014, 63, 290–298. [Google Scholar] [CrossRef]
- Achor, D.; Welker, S.; Mahmoud, S.B.; Wang, C.; Folimonova, S.Y.; Dutt, M.; Gowda, S.; Levy, A. Dynamics of Candidatus Liberibacter asiaticus Movement and Sieve-Pore Plugging in Citrus Sink Cells. Plant Physiol. 2020, 182, 882–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louzada, E.S.; Vasquez, O.E.; Braswell, W.E.; Yanev, G.; Devanaboina, M.; Kunta, M. Distribution of ‘Candidatus Liberibacter asiaticus’ Above and Below Ground in Texas Citrus. Phytopathology 2016, 106, 702–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, H.; Achor, D.; Extberria, E.; Yu, Q.; Du, D.; Stanton, D.; Liang, G.; Gmitter, F.G., Jr. Phloem Regeneration Is a Mechanism for Huanglongbing-Tolerance of “Bearss” Lemon and “LB8-9” Sugar Belle® Mandarin. Plant Sci. 2019, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Da-Graca, J.V.; Douhan, G.W.; Halbert, S.E.; Keremane, M.L.; Lee, R.F.; Vidalakis, G. Huanglongbing: An overview of a complex athosystem ravaging the world’s citrus. J. Integr. Plant Biol. 2016, 58, 373–387. [Google Scholar] [CrossRef] [PubMed]
- Suh, J.H.; Niu, Y.S.; Wang, Z.; Gmitter, F.G.; Wang, Y. Metabolic analysis reveals altered long-chain fatty acid metabolism in the host by Huanglongbing disease. J. Agric. Food. Chem. 2018, 66, 1296–1304. [Google Scholar] [CrossRef] [PubMed]
- Dala-Paula, B.M.; Plotto, A.; Bai, J.; Manthey, J.A.; Baldwin, E.A.; Ferrarezi, R.S. Effect of Huanglongbing or greening disease on orange juice quality, a Review. Plant Sci. 2019, 9, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slisz, A.M.; Breksa, A.P.; Mishchuk, D.O.; McCollum, G.; Slupsky, C.M. Metabolomic analysis of citrus infection by ‘Candidatus Liberibacter’ reveals insight into pathogenicity. J. Proteome Res. 2012, 11, 4223–4230. [Google Scholar] [CrossRef]
- Gómez-Flores, W.; Garza-Saldaña, J.J.; Varela-Fuentes, S.E. Detection of Huanglongbing disease based on intensity-invariant texture analysis of images in the visible spectrum. Comput. Electron. Agric. 2019, 162, 825–835. [Google Scholar] [CrossRef]
- Li, J.; Li, L.; Kolbasov, V.; Ehsani, R.; Carter, E.; Wang, N. Developing citrus Huanglongbing management strategies based on the severity of symptoms in HLB-endemic citrus-producing regions. Phytopathology 2018, 109, 582–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albrecht, U.; Tripathi, I.; Kim, H.; Bowman, K.D. Rootstock effects on metabolite composition in leaves and roots of young navel orange (Citrus sinensis L. Osbeck) and pummelo (C. grandis L. Osbeck) trees. Trees 2018, 33, 243–265. [Google Scholar] [CrossRef]
- Martínez-Ballesta, M.C.; Alcaraz-López, C.; Muries, B.; Mota-Cadenas, C.; Carvajal, M. Physiological aspects of rootstock–scion interactions. Sci. Hortic. 2010, 127, 112–118. [Google Scholar] [CrossRef]
- Warschefsky, E.J.; Klein, L.L.; Frank, M.H.; Chitwood, D.H.; Londo, J.P.; Wettberg, V.E.J.B.; Miller, A.J. Rootstocks: Diversity, domestication, and impacts on shoot phenotypes. Plant. Sci. 2016, 21, 418–437. [Google Scholar] [CrossRef]
- Graham, J.; Gottwald, T.; Setamou, M. Status of Huanglongbing (HLB) outbreaks in Florida, California and Texas. Trop. Plant Pathol. 2020, 45, 265–278. [Google Scholar] [CrossRef]
- Stover, E.; Inch, S.; Richardson, M.L.; Hall, D.G. Conventional Citrus of Some Scion/Rootstock Combinations Show Field Tolerance under High Huanglongbing Disease Pressure. Hort. Sci. 2016, 51, 127–132. [Google Scholar] [CrossRef] [Green Version]
- Aksenov, A.A.; Pasamontes, A.; Peirano, D.J.; Zhao, W.; Dandekar, A.M.; Fiehn, O.; Eshani, N.; Davis, C.E. Detection of Huanglongbing Disease Using Differential Mobility Spectrometry. Anal. Chem. 2014, 86, 2481–2488. [Google Scholar] [CrossRef]
- Garza-Saldaña, J.J.; Varela-Fuentes, S.; Gómez-Flores, W. Métodos para la detección presuntiva de Huanglongbing (HLB) en cítricos. Ciencia UAT 2017, 11, 93–104. [Google Scholar] [CrossRef] [Green Version]
- Arredondo-Valdés, R.; Delgado-Ortíz, J.C.; Beltrán-Beache, M.; Anguiano-Cabello, J.; Cerna-Chávez, E.; Rodríguez-Pagasa, Y.; Ochoa-Fuentes, Y.M. A review of techniques for detecting Huanglongbing (greening) in citrus. Can. J. Microbiol. 2016, 62, 803–811. [Google Scholar] [CrossRef]
- Hitpold, I.; Demarta, L.; Johnson, S.N.; Moore, B.D.; Power, S.A.; Mitchel, C. Silicon and other essential element composition in roots sing X-ray fluorescence spectroscopy: A high throughput approach. In Proceedings of the Ninth ACGIE, 1st ed.; Invertebrate Ecology of Australasian Grasslands: Hawkesbury, Australia, 2017; pp. 191–196. [Google Scholar]
- Canteri, M.H.G.; Renard, C.M.G.; Le Bourvellec, C.; Bureau, S. ATR-FTIR spectroscopy to determine cell wall composition: Application on a large diversity of fruits and vegetables. Carbohydr. Polym. 2019, 212, 186–196. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Hartung, J.S.; Lew, L. Quantitative real-time PCR for detection and identification of Candidatus Liberibacter species associated with citrus Huanglongbing. J. Microbiol. Meth. 2006, 66, 104–115. [Google Scholar] [CrossRef] [PubMed]
- SAS Institute Inc. The SAS® System for Windows® (Ver. 9.0); SAS Institute Inc: Cary, NC, USA, 2004. [Google Scholar]
- Ammar, E.D.; Hall, D.G.; Alvarez, J.M. Effect of Cyantraniliprole, a Novel Insecticide, on the Inoculation of Candidatus Liberibacter Asiaticus Associated with Citrus Huanglongbing by the Asian Citrus Psyllid (Hemiptera: Liviidae). J. Econ. Entomol. 2015, 108, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Flores-Sánchez, J.-L.; Mora-Aguilera, G.; Loaeza-Kuk, E.; López-Arroyo, J.I.; Domínguez-Monge, S.; Acevedo-Sánchez, G.; Robles-García, P. Yield loss caused by Candidatus Liberibacter asiaticus in Persian lime, in Yucatan Mexico. Rev. Mex. Fitopatol. 2015, 33, 195–210. [Google Scholar]
- Folimonova, S.Y.; Robertson, C.J.; Garnsey, S.M.; Gowda, S.; Dawson, W.O. Examination of the Responses of Different Genotypes of Citrus to Huanglongbing (Citrus Greening) Under Different Conditions. Phytopathology 2009, 99, 1346–1354. [Google Scholar] [CrossRef] [Green Version]
- Shokrollah, H.; Abdullah, T.L.; Sijam, K.; Abdullah, S.N.A. Potential use of selected citrus rootstocks and interstocks against HLB disease in Malaysia. Crop Prot. 2011, 30, 521–525. [Google Scholar] [CrossRef]
- Bowman, K.D.; McCollum, G.; Albrecht, U. Performance of ‘Valencia’ orange (Citrus sinensis [L.] Osbeck) on 17 rootstocks in a trial severely affected by huanglongbing. Sci. Hortic. 2016, 201, 355–361. [Google Scholar] [CrossRef] [Green Version]
- Fan, J.; Chen, C.; Yu, Q.; Khalaf, A.; Achor, D.S.; Brlansky, R.H.; Moore, G.A.; Li, Z.G.; Gmitter, F.G. Comparative Transcriptional and Anatomical Analyses of Tolerant Rough Lemon and Susceptible Sweet Orange in Response to “Candidatus Liberibacter asiaticus” Infection. Mol. Plant-Microbe Interact. 2012, 25, 1396–1407. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, U.; McCollum, G.; Bowman, K.D. Influence of rootstock variety on Huanglongbing disease development in field-grown sweet orange (Citrus sinensis [L.] Osbeck) trees. Sci. Hortic. 2012, 138, 210–220. [Google Scholar] [CrossRef]
- Song, X.; Bhattarai, K.; Lv, D.; Gao, F.; Ying, X. Can CRISPR Win the Battle against Huanglongbing? J. Plant Pathol. Microbiol. 2017, 8, 1–5. [Google Scholar] [CrossRef]
- Widyaningsih, S.; Utami, S.N.H.; Joko, T.; Subandiyah, S. Development of disease and growth on six scion/rootstock combinations of citrus seedlings under Huanglongbing pressure. J. Agric. Sci. 2017, 9, 229–238. [Google Scholar] [CrossRef] [Green Version]
- Munir, S.; He, P.; Wu, Y.; He, P.; Khan, S.; Huang, M.; Cui, W.; He, Y. Huanglongbing Control: Perhaps the End of the Beginning. Microb. Ecol. 2017, 76, 192–204. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, U.; Fiehn, O.; Bowman, K.D. Metabolic variations in different citrus rootstock cultivars associated with different responses to Huanglongbing. Int. J. Plant Physiol. Biochem. 2016, 107, 33–44. [Google Scholar] [CrossRef]
- Eticha, D.; Kwast, A.; Chiachia, T.R.S.; Horowitz, N.; Stützel, H. Calcium nutrition of orange and its impact on growth, nutrient uptake and leaf cell wall. Citrus Res. Technol. 2017, 38, 62–70. [Google Scholar] [CrossRef]
- Killiny, N.; Etxeberria, E.; Ponce Flores, A.; González Blanco, P.; Flores Reyes, T.; Ponce Cabrera, L. Laser-induced breakdown spectroscopy (LIBS) as a novel technique for detecting bacterial infection in insects. Sci. Rep. 2019, 9, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Fageria, N.K. The Use of Nutrients in Crop Plants; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2009; 448p. [Google Scholar]
- Ebel, R.C.; Hamido, S.; Morgan, K.T. Interaction of Huanglongbing and Foliar Applications of Copper on Growth and Nutrient Acquisition of Citrus sinensis cv. Valencia. HortScience 2019, 54, 297–302. [Google Scholar] [CrossRef] [Green Version]
- Ranty, B.; Aldon, D.; Cotelle, V.; Galaud, J.P.; Thuleau, P.; Mazars, C. Calcium sensors as key hubs in plant responses to biotic and abiotic stresses. Plant Sci. 2016, 7, 327. [Google Scholar] [CrossRef] [Green Version]
- Demidchik, V.; Shabala, S.; Isayenkov, S.; Cuin, T.A.; Pottosin, I. Calcium transport across plant membranes: Mechanisms and functions. New. Phytol. 2018, 220, 49–69. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; McCollum, G.; Baldwin, E.; Bai, J. Impacts of Huanglongbing symptom severity on fruit detachment force and mechanical properties of sweet oranges (Citrus sinensis). Hort. Sci. 2016, 51, 356–361. [Google Scholar] [CrossRef] [Green Version]
- Gilani, K.; Naz, S.; Aslam, F.; Gurley, W. A Comparison of Zinc, Phosphorous and Potassium levels in leaves and fruit pulp of healthy and Huanglongbing affected citrus cultivars. J. Plant Physiol. Pathol. 2018, 7, 1–8. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.; Nahar, K.; Hossain, M.; Mahmud, J.; Hossen, M.; Fujita, M. Potassium: A vital regulator of plant responses and tolerance to abiotic stresses. Agronomy 2018, 8, 31. [Google Scholar] [CrossRef] [Green Version]
- Souza-Ferraz, R.L.; De Andrade-Barbosa, M.; Araújo-Wanderley, M.J.; Da Silva-Costa, P.; Dourado-Magalhães, I.; De Souza-Medeiros, A.; Miranda-Faria, H.; Marchioro, V.; Soares-de Melo, A.; Alves-dos Anjos, F. Potassium phosphite reduction of Candidatus Liberibacter spp. population on leaves of Ponkan tangerines tree with Huanglongbing. Afr. J. Microbiol. Res. 2018, 12, 248–253. [Google Scholar] [CrossRef] [Green Version]
- Magbalot-Fernández, A.; De Guzman, C.C. Potassium fertilization for higher flowering and fruit yield in ‘Magallanes’ Pummelo (Citrus maxima). AJAHR 2019, 3, 1–8. [Google Scholar] [CrossRef]
- Minzanova, S.T.; Mironov, V.F.; Mironova, L.G.; Nizameev, I.R.; Kholin, K.V.; Voloshina, A.D.; Milyukov, V.A. Synthesis, properties, and antimicrobial activity of pectin complexes with cobalt and nickel. Chem. Nat. Compd. 2016, 52, 26–31. [Google Scholar] [CrossRef]
- Xiao, J.X.; Hu, C.Y.; Chen, Y.Y.; Yang, B.; Hua, J. Effects of low magnesium and an arbuscular mycorrhizal fungus on the growth, magnesium distribution and photosynthesis of two citrus cultivars. Sci. Hortic. 2014, 177, 14–20. [Google Scholar] [CrossRef]
- Li, Q.; Chen, H.H.; Qi, Y.P.; Ye, X.; Yang, L.T.; Huang, Z.R.; Chen, L.S. Excess copper effects on growth, uptake of water and nutrients, carbohydrates, and PSII photochemistry revealed by OJIP transients in Citrus seedlings. Environ. Sci. Pollut. Res. 2019, 26, 30188–30205. [Google Scholar] [CrossRef]
- McGollum, G.; Baldwin, E. Huanglongbing: Devastating Disease of Citrus. In Horticultural Reviews, 1st ed.; Janick, J., Ed.; John Wiley & Sons, Inc: Hoboken, NJ, USA, 2017; Volume 44, pp. 315–362. [Google Scholar]
- Nwugo, C.C.; Duan, Y.; Lin, H. Study on Citrus Response to Huanglongbing Highlights a Down-Regulation of Defense-Related Proteins in Lemon Plants Upon ‘Ca. Liberibacter asiaticus’ Infection. PLoS ONE 2013, 8, e67442. [Google Scholar] [CrossRef]
- Razi, M.F.D.; Khan, I.A.; Jaskani, M.J. Citrus plant nutritional profile in relation to Huanglongbing prevalence in Pakistan. Pak. J. Agric. Sci. 2011, 48, 299–304. [Google Scholar]
- Shahzad, F.; Chun, C.; Schumann, A.; Vashisth, T. Nutrient Uptake in Huanglongbing-affected Sweet Orange: Transcriptomic and Physiological Analysis. J. Amer. Soc. Hort. Sci. 2020, 1–14. [Google Scholar] [CrossRef]
- Duan, Y.; Zhang, M.; Yang, C.; Powell, C.A.; Wang, J.; Huang, Y.; Avery, P.B. Field Evaluation of Integrated Management for Mitigating Citrus Huanglongbing in Florida. Plant Sci. 2019. [Google Scholar] [CrossRef] [Green Version]
- Tian, S.; Lu, L.; Labavitch, J.M.; Webb, S.M.; Xiaoe Yang, X.; Brown, P.H.; He, Z. Spatial imaging of Zn and other elements in Huanglongbingaffected grapefruit by synchrotron-based micro X-ray fluorescence investigation. J. Exp. Bot. 2014, 65, 953–964. [Google Scholar] [CrossRef] [Green Version]
- Shen, W.; Cevallos-Cevallos, J.M.; Rocha, U.N.; Arevalo, H.A.; Stansly, P.A.; Roberts, P.D.; Bruggen, A.H.C. Relation between plant nutrition, hormones, insecticide applications, bacterial endophytes, and Candidatus Liberibacter Ct values in citrus trees infected with Huanglongbing. Eur. J. Plant Pathol. 2013, 137, 727–742. [Google Scholar] [CrossRef]
- Ali, A. Best Practices of Integrated Plant Nutrition. In Best Practices of Integrated Plant Nutrition, System in SAARC Countries, 1st ed.; Jahan, F.N., Gurung, T.R., Eds.; SAARC Agriculture Centre: Dhaka, Bangladesh, 2017; Volume 8, pp. 104–134. [Google Scholar]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |



{kind=link}
{kind=link}
| C. limon | C. sinensis | ||||
|---|---|---|---|---|---|
| Plant | C. aurantium | C. macrophylla | C. volkameriana | C. aurantium | C. volkameriana |
| Cq Value | |||||
| 1 | 26.28 | 32.62 | 35.34 | 34.42 | 30.26 |
| 2 | 35.43 | 35.11 | 36.58 | 33.12 | 30.23 |
| 3 | 33.12 | 34.67 | 34.85 | 32.56 | 29.71 |
| 4 | 34.32 | 34.39 | 36.36 | 34.34 | 28.50 |
| 5 | 32.18 | 34.08 | 35.29 | 34.27 | 30.64 |
| 6 | 33.57 | 37.25 | 32.92 | 32.03 | 30.15 |
| 7 | 33.75 | 33.30 | 34.02 | 24.05 | 29.75 |
| 8 | 35.26 | 34.81 | 32.56 | 32.96 | 29.25 |
| 9 | 35.36 | 36.20 | 34.74 | 33.28 | 31.57 |
| 10 | 34.02 | 35.48 | 33.75 | 31.9 | 30.12 |
| 11 | 32.32 | 37.23 | 38.11 | 29.05 | 29.44 |
| 12 | 34.04 | 35.29 | 35.02 | 34.64 | 31.14 |
| 13 | 31.59 | 36.37 | 34.53 | 31.23 | 31.09 |
| 14 | 36.34 | 37.27 | 36.28 | 32.93 | 21.11 |
| 15 | 34.72 | 37.64 | 37.47 | 35.88 | 27.65 |
| 16 | 34.66 | 36.68 | 36.15 | 26.30 | 31.02 |
| 17 | 34.18 | 36.04 | 36.11 | 26.02 | 30.18 |
| 18 | 32.95 | 36.82 | 36.41 | 25.50 | 30.12 |
| 19 | 32.55 | 36.39 | 34.54 | 27.07 | 30.76 |
| 20 | 35.12 | 36.28 | 35.06 | 24.06 | 31.43 |
| 21 | 32.19 | 36.95 | 34.22 | 24.60 | 29.74 |
| 22 | 33.96 | 31.04 | 34.65 | 27.11 | 32.84 |
| 23 | 32.08 | 37.12 | 34.63 | 28.12 | 29.52 |
| 24 | 35.07 | 36.07 | 34.32 | 28.17 | 32.02 |
| 25 | 35.47 | 35.61 | 34.46 | 28.91 | 29.45 |
| 26 | 34.42 | 37.06 | 36.46 | 19.44 | 18.46 |
| 27 | 34.16 | 36.06 | 36.21 | 23.79 | 31.26 |
| 28 | 36.4 | 36.93 | 36.22 | 25.46 | 31.44 |
| 29 | 34.75 | 36.86 | 34.42 | 24.39 | 31.60 |
| 30 | 34.81 | 36.91 | 34.24 | 26.53 | 28.39 |
| 31 | 34.90 | 37.06 | 36.37 | 26.97 | 29.38 |
| 32 | 36.08 | 36.97 | 35.87 | 28.78 | 30.82 |
| 33 | 35.25 | 36.95 | 35.02 | 27.02 | 33.52 |
| 34 | 34.81 | 36.31 | 34.52 | 27.01 | 31.67 |
| 35 | 36.12 | 36.89 | 34.44 | 26.95 | 32.78 |
| 36 | 36.13 | 37.55 | 34.41 | 26.83 | 33.12 |
| 37 | 35.32 | 37.15 | 34.62 | 29.58 | 31.51 |
| 38 | 34.31 | 36.39 | 34.67 | 27.41 | 33.40 |
| 39 | 36.40 | 36.57 | 34.63 | 23.46 | 32.63 |
| 40 | 35.28 | 36.10 | 34.77 | 28.34 | 31.84 |
| Specie | Rootstock | Cq Values |
|---|---|---|
| C. limon | C. aurantium | 34.24 b |
| C. macrophylla | 36.06 a | |
| C. volkameriana | 35.13 ab | |
| C. sinensis | C. aurantium | 28.61 d |
| C. volkameriana | 30.23 c |
| Species | Presence | Height (cm) | Stem (mm) |
|---|---|---|---|
| C. limon | Not infected | 167.66 a | 16.44 a |
| Infected | 179.93 a | 16.81 a | |
| MSD | 13.14 | 1.25 | |
| C. sinensis | Not infected | 149.40 a | 15.26 a |
| Infected | 154.43 a | 15.74 a | |
| MSD | 19.83 | 2.08 |
| C. limon | ||||
| Rootstock | Not Infected | Infected | ||
| Height (cm) | Stem (mm) | Height (cm) | Stem (mm) | |
| C. volkameriana | 170.60 a | 17.82 a | 196.88 a | 19.35 a |
| C. macrophylla | 162.40 a | 16.86 a | 176.27 b | 16.32 b |
| C. aurantium | 167.0 a | 15.18 a | 166.63 b | 14.76 c |
| MSD | 51.95 | 4.31 | 10.70 | 0.67 |
| C. sinensis | ||||
| Rootstock | Not Infected | Infected | ||
| Height (cm) | Stem (mm) | Height (cm) | Stem (mm) | |
| C. volkameriana | 149.20 a | 18.02 a | 169.45 a | 17.48 a |
| C. aurantium | 149.60 a | 13.46 b | 131.61 b | 13.04 b |
| MSD | 22.05 | 2.73 | 12.17 | 0.99 |
| C. limon (ppm) | ||||||||
| Mg | Al | Si | P | S | Cl | K | Ca | |
| NI | 7.40 a | 1.02 a | 0.32 a | 1.38 a | 3.97 a | 6.15 a | 27.89 a | 50.33 a |
| I | 8.47 a | 1.04 a | 0.56 a | 1.40 a | 3.27 b | 7.63 a | 24.56 a | 51.47 a |
| MSD | 1.23 | 0.34 | 0.96 | 0.49 | 0.32 | 2.01 | 4.98 | 6.7 |
| Sc | Ti | V | Cr | Mn | Fe | Cu | Zn | |
| NI | 0.11 a | 0.0009 a | 0.00002 a | 0.00001 a | 0.08 a | 0.23 a | 0.08 b | 0.03 a |
| I | 0.11 a | 0.00009 a | 0.00002 a | 0.00001 a | 0.08 a | 0.24 a | 0.01 a | 0.03 a |
| MSD | 0.03 | 0.01 | 0.0000005 | 0.0000002 | 0.01 | 0.12 | 0.02 | 0.02 |
| C. sinensis (ppm) | ||||||||
| Mg | Al | Si | P | S | Cl | K | Ca | |
| NI | 10.18 a | 0.98 a | 0.56 a | 1.51 a | 2.99 a | 9.88 a | 20.52 a | 51.79 a |
| I | 9.8 a | 0.99 a | 0.46 a | 1.48 a | 3.63 a | 8.38 a | 17.42 a | 56.22 a |
| MSD | 8.75 | 1.10 | 0.60 | 0.38 | 6.51 | 10.30 | 54.12 | 27.70 |
| Sc | Ti | V | Cr | Mn | Fe | Cu | Zn | |
| NI | 0.11 a | 0.0050 a | 0.00003 a | 0.00002 a | 0.08 a | 0.20 b | 0.10 a | 0.03 a |
| I | 0.07 a | 0.00008 a | 0.00003 a | 0.000009 a | 0.07 a | 0.25 a | 0.08 a | 0.03 a |
| MSD | 0.98 | 0.06 | 0.0001 | 0.0000006 | 0.29 | 0.03 | 0.04 | 0.06 |
| Variable | Can1 | Can2 | Can3 |
|---|---|---|---|
| Cl | 89.39737621 | 2.96360649 | −0.78333747 |
| K | 57.79235462 | 3.53450965 | −1.92557449 |
| Ca | 90.20094892 | 2.60397644 | −2.05580445 |
| Height | 28.91402537 | 1.13598256 | −0.16235885 |
| Stem | −15.01864614 | −0.50627275 | 0.78861065 |
| Presence | 0 | 1 | 2 | 3 |
|---|---|---|---|---|
| 0 | 0 | 5695 | 7398 | 15914 |
| 1 | 5695 * | 0 | 114.04 | 2573 |
| 2 | 7398 * | 114.04 § | 0 | 1614 |
| 3 | 15914 * | 2573 * | 1614 * | 0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sáenz-Pérez, C.A.; Osorio-Hernández, E.; Estrada-Drouaillet, B.; Castro-Nava, S.; Delgado-Martínez, R.; López-Badillo, C.M.; Rodríguez-Herrera, R. Rootstock Influence on Growth and Mineral Content of Citrus limon and Citrus sinensis cv. Valencia Inoculated with Candidatus Liberibacter Asiaticus. Agronomy 2020, 10, 1564. https://doi.org/10.3390/agronomy10101564
Sáenz-Pérez CA, Osorio-Hernández E, Estrada-Drouaillet B, Castro-Nava S, Delgado-Martínez R, López-Badillo CM, Rodríguez-Herrera R. Rootstock Influence on Growth and Mineral Content of Citrus limon and Citrus sinensis cv. Valencia Inoculated with Candidatus Liberibacter Asiaticus. Agronomy. 2020; 10(10):1564. https://doi.org/10.3390/agronomy10101564
Chicago/Turabian StyleSáenz-Pérez, Criseida Alhelí, Eduardo Osorio-Hernández, Benigno Estrada-Drouaillet, Sergio Castro-Nava, Rafael Delgado-Martínez, Claudia Magdalena López-Badillo, and Raúl Rodríguez-Herrera. 2020. "Rootstock Influence on Growth and Mineral Content of Citrus limon and Citrus sinensis cv. Valencia Inoculated with Candidatus Liberibacter Asiaticus" Agronomy 10, no. 10: 1564. https://doi.org/10.3390/agronomy10101564