Agro-Morphological Exploration of Some Unexplored Wild Vigna Legumes for Domestication

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Preparation
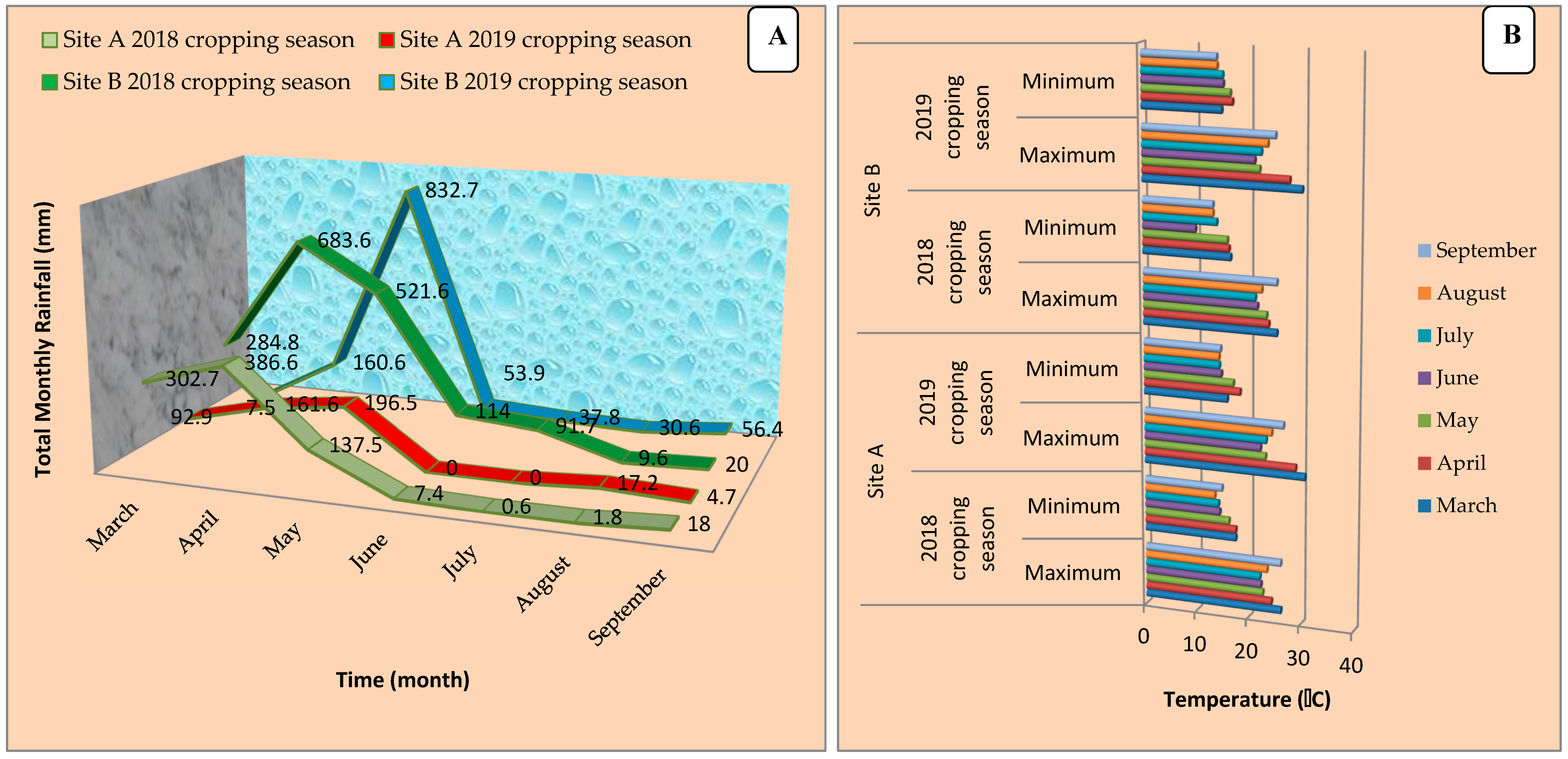
2.2. Study Sites and Meteorological Considerations
2.3. Experimental Design and Planting Process
2.4. Data Collection and Analysis
3. Results
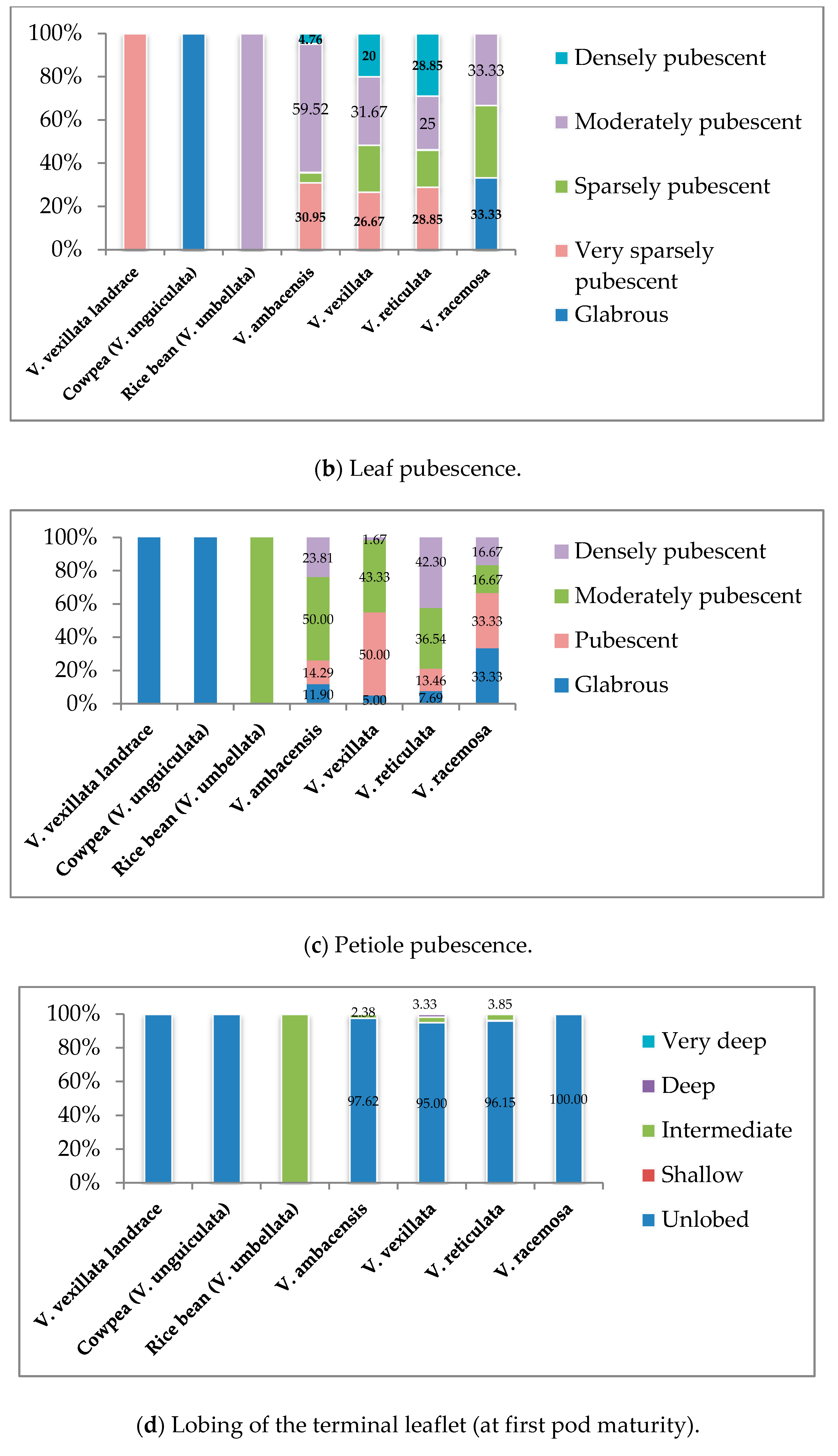
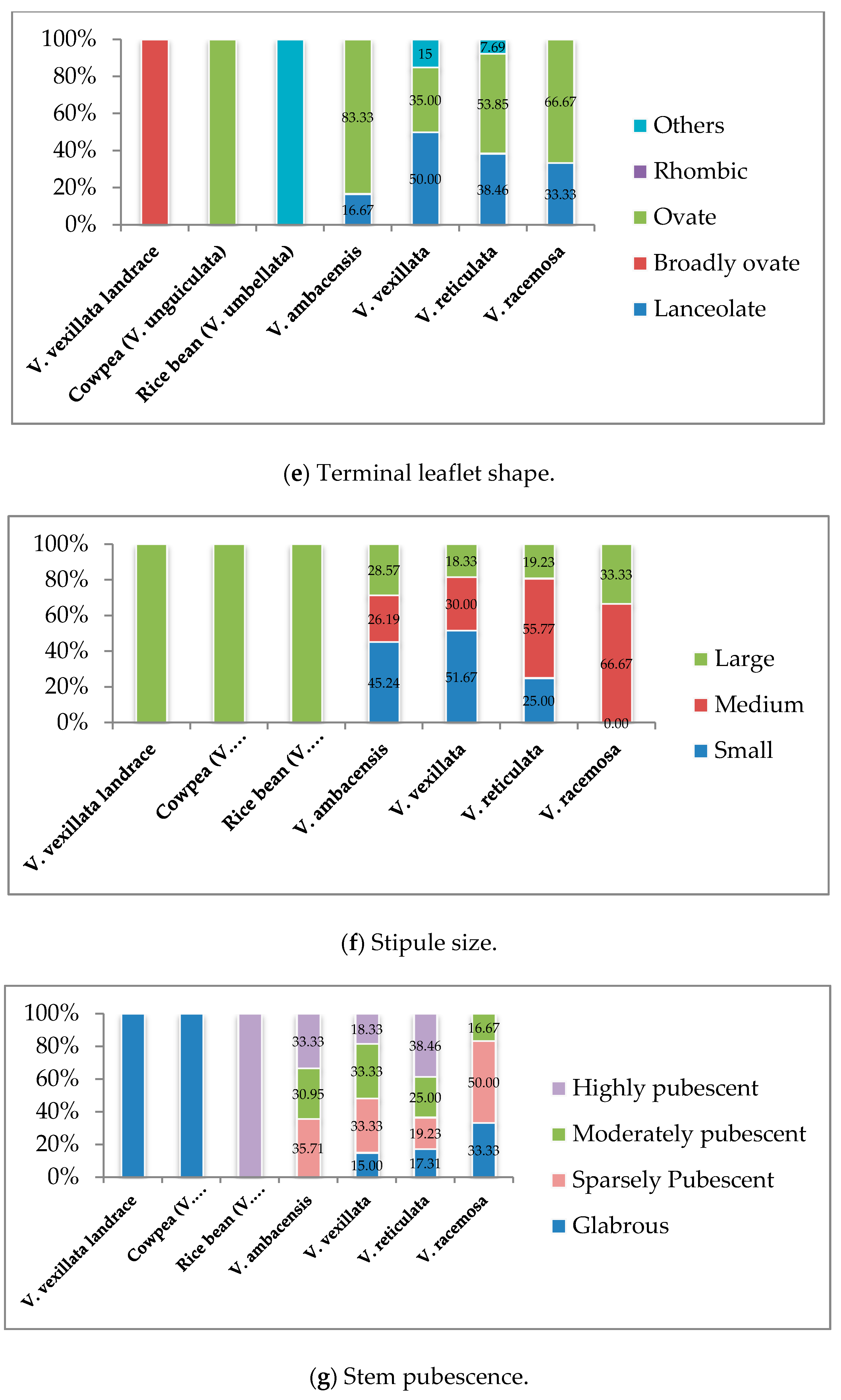
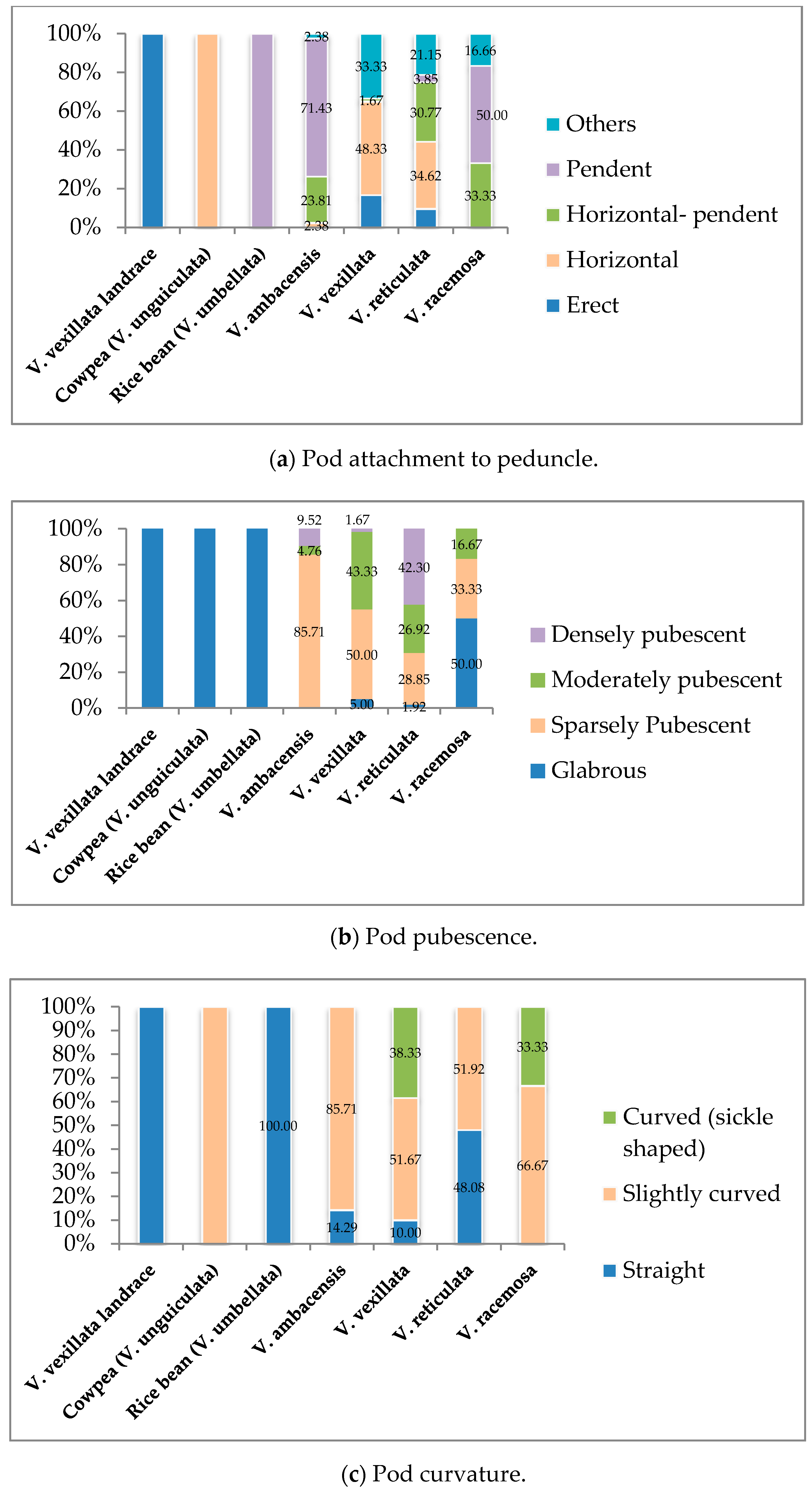
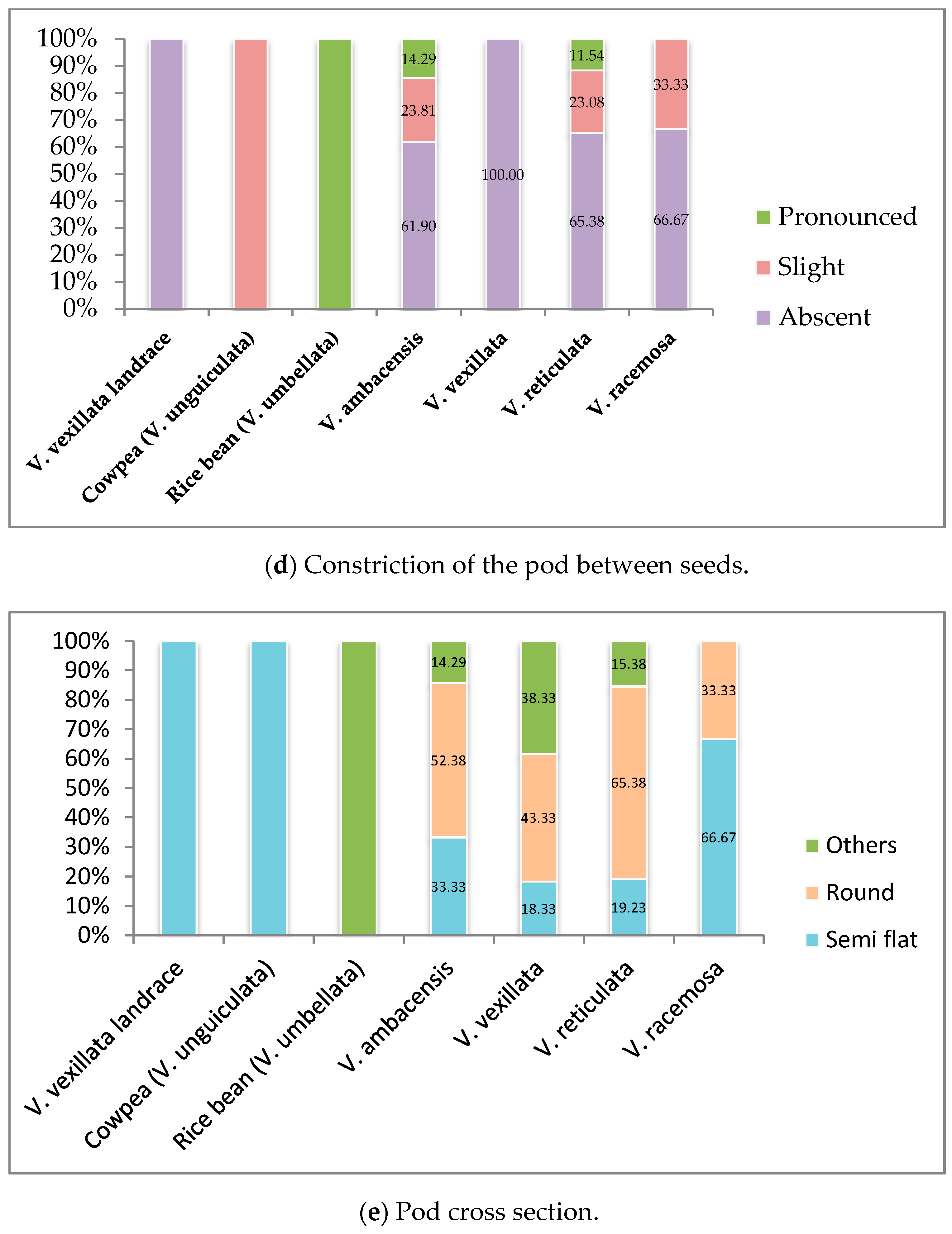
3.1. Qualitative Traits Exploration of the Wild Unexplored Vigna Species
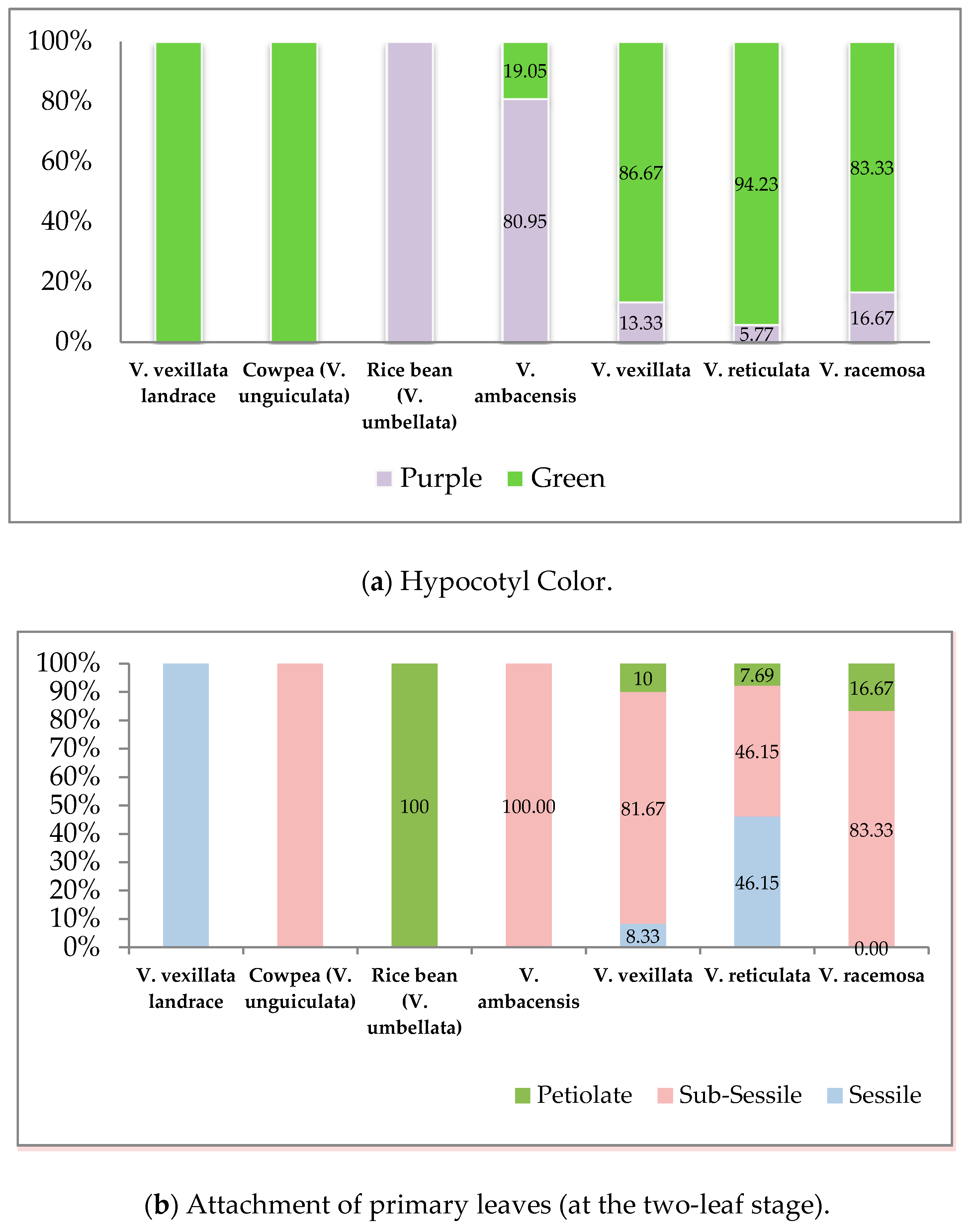
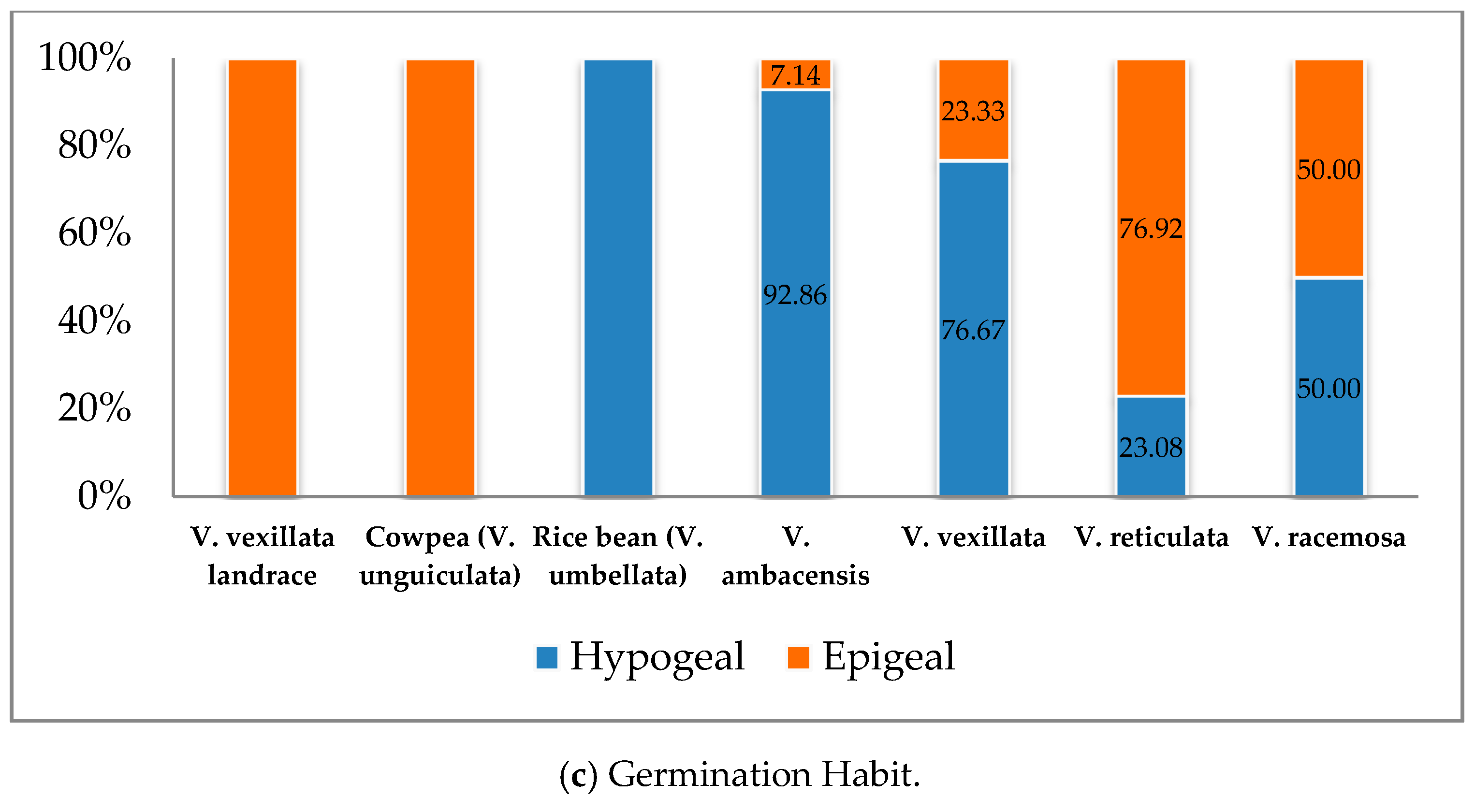
3.1.1. Germination Stage
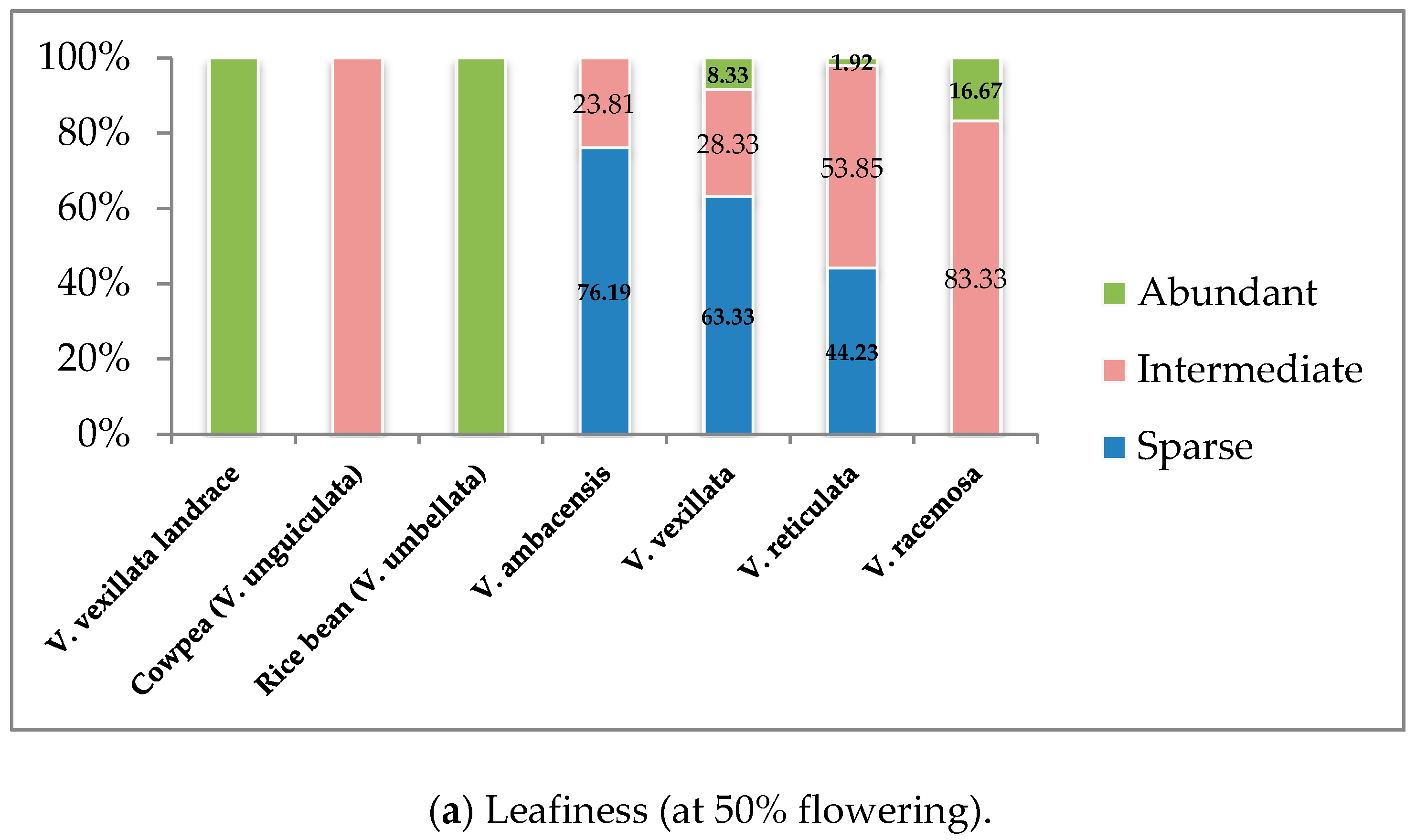
3.1.2. Vegetative Stage
3.1.3. Pod Formation and Maturity Stage
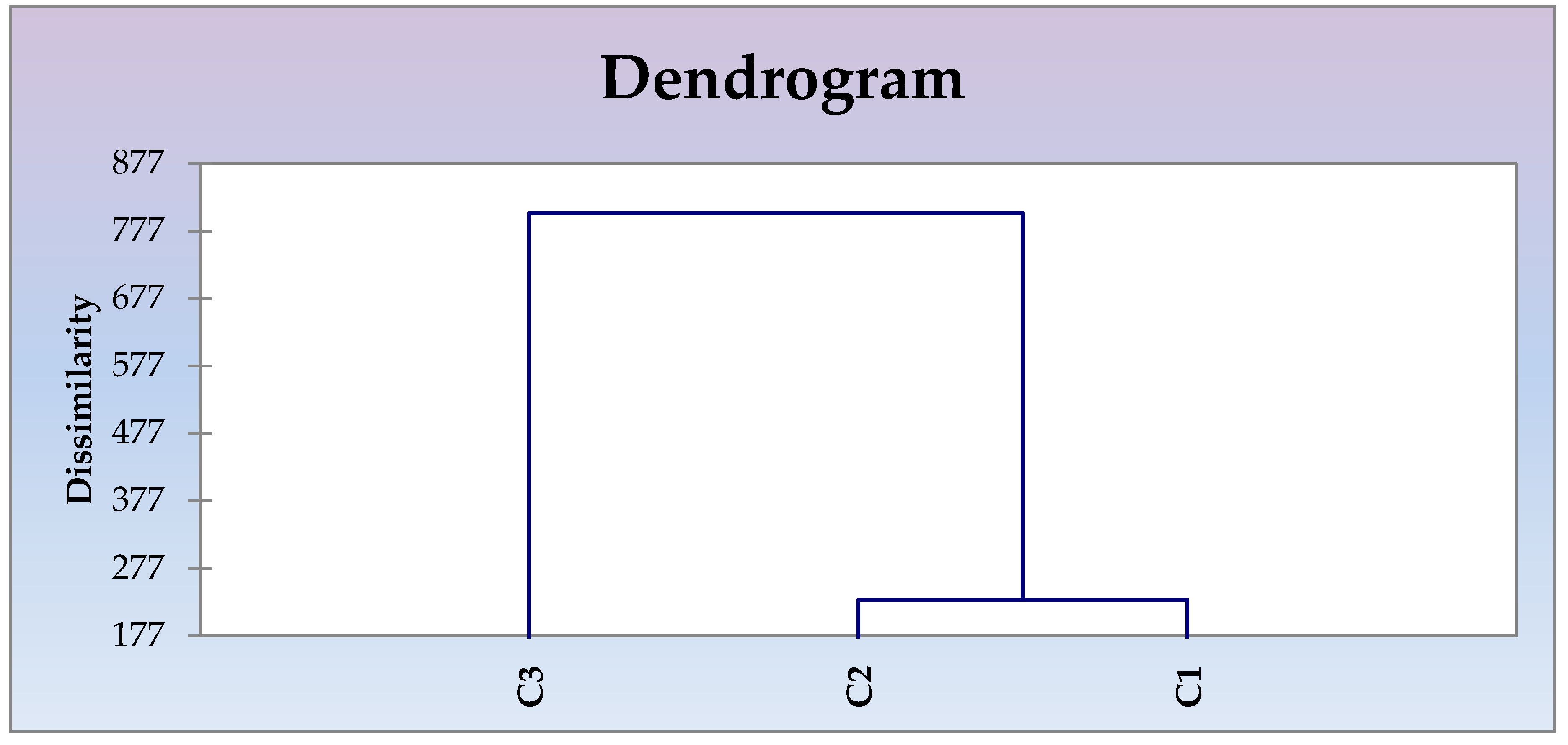
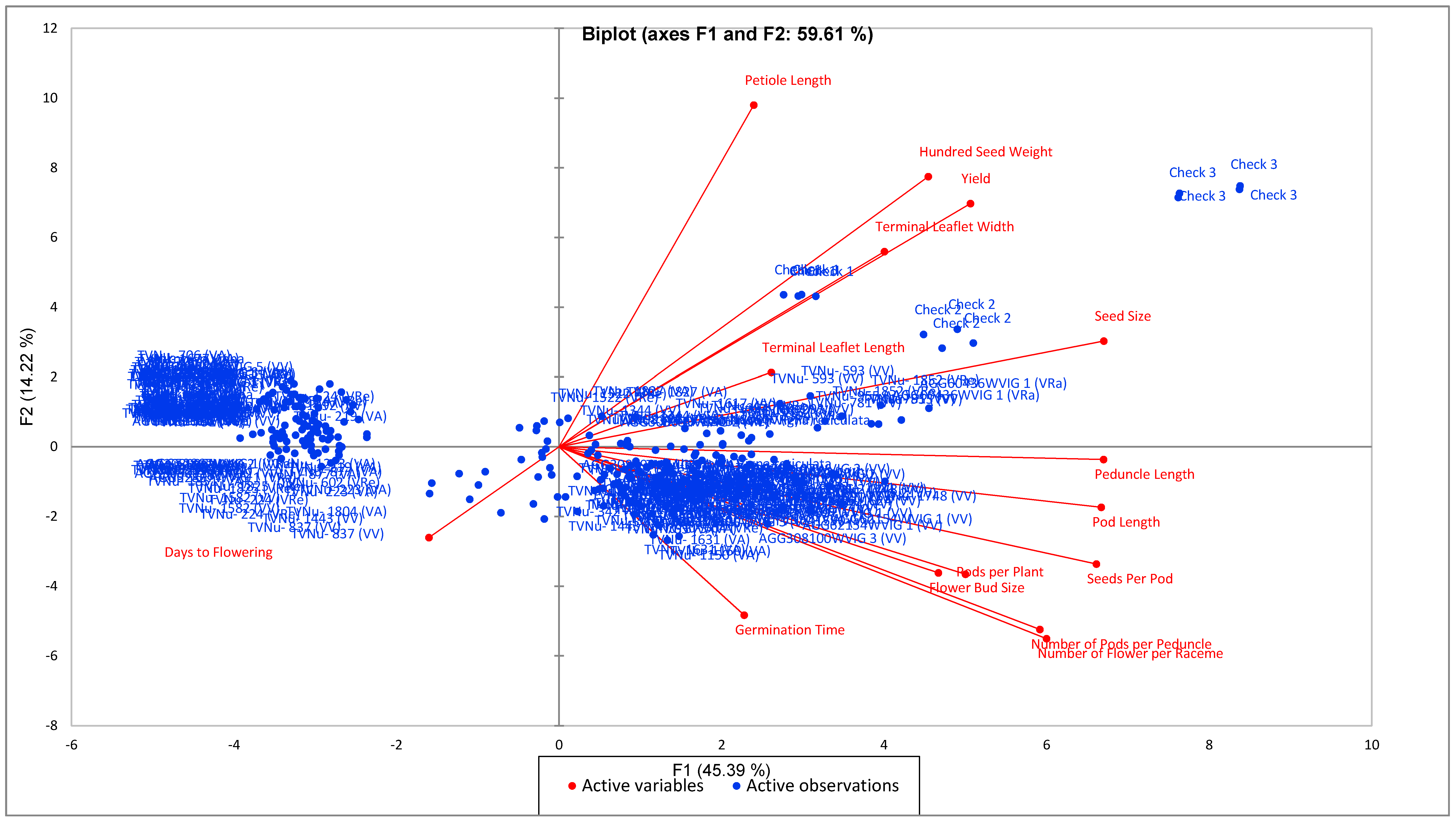
3.2. Quantitative Traits Exploration of the Wild Unexplored Vigna Species
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Singh, A.; Dubey, P.K.; Chaurasia, R.; Dubey, R.K.; Pandey, K.K.; Singh, G.S.; Abhilash, P.C. Domesticating the Undomesticated for Global Food and Nutritional Security: Four Steps. Agronomy 2019, 9, 491. [Google Scholar] [CrossRef] [Green Version]
- Kew, R.B.G. The State of the World’s Plants Report; Kew:Royal Botanic Gardens: London, UK, 2016. [Google Scholar]
- Padulosi, S.; Thompson, J.; Rudebjer, P. Fighting Poverty, Hunger and Malnutrition with Neglected and Underutilized Species (NUS): Needs, Challenges and the Way Forward; Bioversity International: Rome, Italy, 2013; ISBN 9789290439417. [Google Scholar]
- Diouf, J. FAO’s Director-general on how to feed the world in 2050. Popul. Dev. Rev. 2009, 35, 837–839. [Google Scholar]
- Harouna, D.V.; Venkataramana, P.B.; Ndakidemi, P.A.; Matemu, A.O. Under-exploited wild Vigna species potentials in human and animal nutrition: A review. Glob. Food Sec. 2018, 18, 1–11. [Google Scholar] [CrossRef]
- Bhat, R.; Karim, A.A. Exploring the nutritional potential of wild and underutilized legumes. Compr. Rev. Food Sci. Food Saf. 2009, 8, 305–331. [Google Scholar] [CrossRef]
- Ojiewo, C.; Monyo, E.; Desmae, H.; Boukar, O.; Mukankusi-Mugisha, C.; Thudi, M.; Pandey, M.K.; Saxena, R.K.; Gaur, P.M.; Chaturvedi, S.K.; et al. Genomics, genetics and breeding of tropical legumes for better livelihoods of smallholder farmers. Plant Breed. 2018, 1–13. [Google Scholar] [CrossRef]
- Pratap, A.; Malviya, N.; Tomar, R.; Gupta, D.S.; Jitendra, K. Vigna. In Alien Gene Transfer in Crop Plants, Volume 2: Achievements and Impacts; Pratap, A., Kumar, J., Eds.; Springer Science+Business Media, LLC: Berlin/Heidelberg, Germany, 2014; pp. 163–189. ISBN 9781461495727. [Google Scholar]
- Tomooka, N.; Naito, K.; Kaga, A.; Sakai, H.; Isemura, T.; Ogiso-Tanaka, E.; Iseki, K.; Takahashi, Y. Evolution, domestication and neo-domestication of the genus Vigna. Plant Genet. Resour. Characterisation Util. 2014, 12, S168–S171. [Google Scholar] [CrossRef]
- Tomooka, N.; Kaga, A.; Isemura, T.; Vaughan, D.; Srinives, P.; Somta, P.; Thadavong, S.; Bounphanousay, C.; Kanyavong, K.; Inthapanya, P. Vigna Genetic Resources. In Proceedings of the 14th NIAS International Workshop on Genetic Resources: Genetic Resources and Comparative Economics of Legumes (Glycine and Vigna), Tsukuba, Japan, 14 September 2009; pp. 11–21. [Google Scholar]
- Smýkal, P.; Nelson, M.N.; Berger, J.D.; Von Wettberg, E.J.B. The impact of genetic changes during crop domestication. Agronomy 2018, 8, 119. [Google Scholar] [CrossRef] [Green Version]
- Bisht, I.S.; Bhat, K.V.; Lakhanpaul, S.; Latha, M.; Jayan, P.K.; Biswas, B.K.; Singh, A.K. Diversity and genetic resources of wild Vigna species in India. Genet. Resour. Crop Evol. 2005, 52, 53–68. [Google Scholar] [CrossRef]
- IASRI Augmented Block Designs. Available online: http://www.iasri.res.in/design/Augmented Designs/home.htm (accessed on 18 January 2019).
- Gore, P.G.; Tripathi, K.; Pratap, A.; Bhat, K.V.; Umdale, S.D.; Gupta, V.; Pandey, A. Delineating taxonomic identity of two closely related Vigna species of section Aconitifoliae: V. trilobata (L.) Verdc. and V. stipulacea (Lam.) Kuntz in India. Genet. Resour. Crop Evol. 2019, 66, 1155–1165. [Google Scholar] [CrossRef]
- Oyatomi, O.; Fatokun, C.; Boukar, O.; Abberton, M.; Ilori, C. Screening Wild Vigna Species and Cowpea (Vigna unguiculata) Landraces for Sources of Resistance to Striga gesnerioides. In Enhancing Crop Genepool Use: Capturing Wild Relatives and Landrace Diversity for Crop Improvement; CAB International: Wallingford, UK, 2016; pp. 27–31. [Google Scholar]
- Popoola, J.; Adebambo, A.; Ejoh, S.; Agre, P.; Adegbite, A.; Omonhinmin, C. Morphological Diversity and Cytological Studies in Some Accessions of Vigna vexillata (L.) A. Richard. Annu. Res. Rev. Biol. 2017, 19, 1–12. [Google Scholar] [CrossRef]
- Swanson, B.G.; Hughes, J.S.; Rasmussen, P.H. Seed Microstructure: Review of Water Imbibition in Legumes. Food Struct. 1985, 4, 115–124. [Google Scholar]
- Smykal, P.; Vernoud, V.; Blair, M.W.; Soukup, A.; Thompson, R.D. The role of the testa during development and in establishment of dormancy of the legume seed. Front. Plant Sci. 2014, 5, 351. [Google Scholar] [PubMed] [Green Version]
- Jiang, H.; Egli, D.B. Shade Induced Changes in Flower and Pod Number and Flower and Fruit Abscission in Soybean. Agron. J. 1993, 85, 221–225. [Google Scholar] [CrossRef]
- Morgan, J.B.; Connolly, E.L. Plant-Soil Interactions: Nutrient Uptake. Nat. Educ. Knowl. 2013, 4, 2. [Google Scholar]
- Kigel, J.; Konsens, I.; Rosen, N.; Rotem, G.; Kon, A.; Fragman-Sapir, O. Relationships between flowering time and rainfall gradients across Mediterranean-desert transects. Isr. J. Ecol. Evol. 2011, 57, 91–109. [Google Scholar] [CrossRef]
- Satish, D.; Rc, J.; Patil, R.K.; Masuthi, D. Genotype x environment interaction and stability analysis in Recombinant inbred lines of French bean for growth and yield components. J. Pharmacogn. Phytochem. 2017, 6, 216–219. [Google Scholar]
- Sabaghnia, N.; Dehghani, H.; Sabaghpourb, S.H. Graphic Analysis of Genotype by Environment Interaction for Lentil Yield in Iran. Agron. J. 2008, 100, 760–764. [Google Scholar] [CrossRef]
- Arteaga, S.; Yabor, L.; Torres, J.; Solbes, E.; Muñoz, E.; Vicente, O.; Boscaiu, M. Morphological and agronomic characterization of Spanish landraces of Phaseolus vulgaris L. Agriculture 2019, 9, 149. [Google Scholar] [CrossRef] [Green Version]
- Adewale, B.D.; Okonji, C.; Oyekanmi, A.A.; Akintobi, D.A.C.; Aremu, C.O. Genotypic variability and stability of some grain yield components of Cowpea. Afr. J. Agric. Res. 2010, 5, 874–880. [Google Scholar]
- Harouna, D.V.; Venkataramana, P.B.; Matemu, A.O.; Ndakidemi, P.A. Assessment of Water Absorption Capacity and Cooking Time of Wild Under-Exploited Vigna Species towards their Domestication. Agronomy 2019, 9, 509. [Google Scholar] [CrossRef] [Green Version]
- Harouna, D.V.; Venkataramana, P.B.; Matemu, A.O.; Ndakidemi, P.A. Wild Vigna Legumes: Farmers’ Perceptions, Preferences, and Prospective Uses for Human Exploitation. Agronomy 2019, 9, 284. [Google Scholar] [CrossRef] [Green Version]
- Henry, R. Domesticating Vigna Stipulacea: A Potential Legume Crop With Broad Resistance to Biotic Stresses. Front. Plant Sci. 2019, 10, 1–12. [Google Scholar]
- Smýkal, P.; Nelson, M.N.; Berger, J.D.; Von Wettberg, E.J.B. The impact of genetic changes during crop domestication on healthy food development. Agronomy 2018, 8, 26. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Descriptors | |
|---|---|---|
| Qualitative Traits | ||
| 1 | Seed germination habit | 1. Epigeal, 2. Hypogeal |
| 2 | Attachment of primary leaves (at two-leaf stage) | 1. Sessile, 2. Sub-sessile, 3. Petiolate |
| 3 | Growth habit (recorded at first pod maturity) | 1. Erect, 2. Semi-erect, 3. Spreading, 4. Semi-prostrate, 5. Prostrate, 6. Climbing |
| 4 | Leafiness (at 50% flowering) | 1. Sparse, 2. Intermediate, 3. Abundant |
| 5 | Leaf pubescence | 1. Glabrous, 2. Very sparsely pubescent, 3. Sparsely pubescent, 4. Moderately Pubescent, 5. Densely pubescent |
| 6 | Petiole pubescence | 1. Glabrous, 2. Pubescent, 3. Moderately pubescent, 4. Densely pubescent |
| 7 | Lobing of terminal leaflet (at first pod maturity) | 1. Unlobed, 2. Shallow, 3. Intermediate, 4. Deep, 5. Very deep |
| 8 | Terminal leaflet shape | 1. Lanceolate, 2. Broadly ovate, 3. Ovate, 4. Rhombic, 5. Others |
| 9 | Stipule size | 1. Small, 2. Medium, 3. Large |
| 10 | Hypocotyl color | 1. Green; 2. Purple, 3. Others |
| 11 | Stem pubescence | 1. Glabrous, 2. Sparsely pubescent, 3. Moderately pubescent, 4. Highly pubescent |
| 12 | Pod attachment to peduncle | 1. Erect, 2. Horizontal, 3. Horizontal-pendent 4. Pendent, 5. Others |
| 13 | Pod pubescence | 1. Glabrous, 2. Sparsely pubescent, 3. Moderately pubescent, 4. Densely pubescent |
| 14 | Pod curvature | 1. Straight, 2. Slightly curved, 3. Curved (sickle shaped) |
| 15 | Constriction of pod between seeds | 1. Absent, 2. Slight, 3. Pronounced |
| Pod cross section | 1. Semi flat, 2. Round, 3. Others | |
| Quantitative Traits | ||
| 1 | Germination time (days) | |
| 2 | Terminal leaflet length (cm) | |
| 3 | Terminal leaflet width (cm) | |
| 4 | Petiole length (cm) | |
| 5 | Days to flowering | |
| 6 | Flower bud size (cm) | |
| 7 | Number of flowers per raceme | |
| 8 | Peduncle length (cm) | |
| 9 | Pods per peduncle | |
| 10 | Pod length (cm) | |
| 11 | Pods per plant | |
| 12 | Seeds per pod | |
| 13 | Seed size (mm2) | |
| 14 | 100-Seed weight (g) | |
| 15 | Yield (Kg/ha) | |
| (a) | |||||||||||||||
| Traits | ANOVA | Type III Sum of Squares Analysis | |||||||||||||
| Model | Block Effect | Accession Effect | |||||||||||||
| DF | Sum of Squares | Mean Squares | F | Pr > F | DF | Sum of Squares | Mean Squares | F | p | DF | Sum of Squares | Mean Squares | F | p | |
| 1 | 172 | 230.50 | 1.34 | 0.00 | <0.0001 | 7 | 0.00 | 0.00 | - | - | 165 | 230.19 | 1.40 | 0.00 | <0.0001 |
| 2 | 172 | 2513.26 | 14.61 | 11.86 | <0.0001 | 7 | 36.22 | 5.17 | 4.20 | 0.0019 | 165 | 1512.31 | 9.17 | 7.44 | <0.0001 |
| 3 | 172 | 685.22 | 3.98 | 2.99 | 0.0001 | 7 | 15.18 | 2.17 | 1.63 | 0.1600 | 165 | 675.48 | 4.09 | 3.07 | 0.0001 |
| 4 | 172 | 1999.94 | 11.63 | 6.57 | <0.0001 | 7 | 72.68 | 10.38 | 5.86 | 0.0001 | 165 | 1908.16 | 11.56 | 6.53 | <0.0001 |
| 5 | 172 | 41,888.19 | 243.54 | 269.88 | <0.0001 | 7 | 2.67 | 0.38 | 0.42 | 0.8818 | 165 | 41,253.80 | 250.02 | 277.07 | <0.0001 |
| 6 | 172 | 92.42 | 0.54 | 21.64 | <0.0001 | 7 | 0.63 | 0.09 | 3.65 | 0.0046 | 165 | 75.72 | 0.46 | 18.48 | <0.0001 |
| 7 | 172 | 5907.56 | 34.35 | 14.23 | <0.0001 | 7 | 24.98 | 3.57 | 1.48 | 0.2069 | 165 | 165 | 4000.12 | 24.24 | 10.05 |
| 8 | 172 | 16,033.67 | 93.22 | 68.36 | <0.0001 | 7 | 25.65 | 3.66 | 2.69 | 0.0245 | 165 | 12,837.61 | 77.80 | 57.05 | <0.0001 |
| 9 | 172 | 759.23 | 4.41 | 4.96 | <0.0001 | 7 | 10.55 | 1.51 | 1.69 | 0.1427 | 165 | 539.13 | 3.27 | 3.67 | <0.0001 |
| 10 | 172 | 4990.46 | 29.01 | 27.94 | <0.0001 | 7 | 13.24 | 1.89 | 1.82 | 0.1139 | 165 | 3894.89 | 23.61 | 22.73 | <0.0001 |
| 11 | 172 | 298,644.75 | 1736.31 | 365.40 | <0.0001 | 7 | 81.81 | 11.69 | 2.46 | 0.0367 | 165 | 221,943.38 | 1345.11 | 283.07 | <0.0001 |
| 12 | 172 | 3427.67 | 19.93 | Infini | <0.0001 | 7 | 0.00 | 0.00 | 165 | 2597.62 | 15.74 | Infini | <0.0001 | ||
| 13 | 172 | 26,079.22 | 151.62 | 29.99 | <0.0001 | 7 | 49.43 | 7.06 | 1.40 | 0.2377 | 165 | 23,910.77 | 144.91 | 28.66 | <0.0001 |
| 14 | 172 | 6155.01 | 35.78 | 14.06 | <0.0001 | 7 | 60.79 | 8.68 | 3.41 | 0.0070 | 165 | 5923.05 | 35.90 | 14.11 | <0.0001 |
| 15 | 172 | 225,200,114.2 | 1,309,303.0 | 14.99 | <0.0001 | 7 | 2,007,022.1 | 286,717.4 | 3.28 | 0.0087 | 165 | 218,001,052.5 | 1,321,218.5 | 15.13 | <0.0001 |
| (b) | |||||||||||||||
| Traits | Contrast (Differences) | ||||||||||||||
| Among Accessions | Among Checks | Check vs. Accession | |||||||||||||
| DF | Sum of Squares | Mean Squares | F | Pr > F | DF | Sum of Squares | Mean Squares | F | p | DF | Sum of Squares | Mean Squares | F | p | |
| 1 | 53 | 36.59 | 0.69 | Infini | <0.0001 | 3 | 16.00 | 5.33 | Infini | <0.0001 | 1 | 49.51 | 49.51 | Infini | <0.0001 |
| 2 | 53 | 397.53 | 7.50 | 6.09 | <0.0001 | 3 | 80.92 | 26.97 | 21.89 | <0.0001 | 1 | 49.72 | 49.72 | 40.35 | <0.0001 |
| 3 | 53 | 181.78 | 3.43 | 2.58 | 0.0019 | 3 | 28.15 | 9.38 | 7.05 | 0.0008 | 1 | 130.73 | 130.73 | 98.16 | <0.0001 |
| 4 | 53 | 779.54 | 14.71 | 8.31 | <0.0001 | 3 | 132.98 | 44.33 | 25.03 | <0.0001 | 1 | 343.41 | 343.41 | 193.94 | <0.0001 |
| 5 | 53 | 17,494.29 | 330.08 | 365.79 | <0.0001 | 3 | 55.09 | 18.36 | 20.35 | <0.0001 | 1 | 1095.79 | 1095.79 | 1214.34 | <0.0001 |
| 6 | 53 | 17.56 | 0.33 | 13.34 | <0.0001 | 3 | 0.29 | 0.10 | 3.84 | 0.0178 | 1 | 0.29 | 0.29 | 11.70 | 0.0016 |
| 7 | 53 | 984.25 | 18.57 | 7.70 | <0.0001 | 3 | 137.32 | 45.77 | 18.97 | <0.0001 | 1 | 159.39 | 159.39 | 66.05 | <0.0001 |
| 8 | 53 | 1790.74 | 33.79 | 24.78 | <0.0001 | 3 | 240.59 | 80.20 | 58.81 | <0.0001 | 1 | 1751.07 | 1751.07 | 1284.07 | <0.0001 |
| 9 | 53 | 141.29 | 2.67 | 3.00 | 0.0004 | 3 | 27.70 | 9.23 | 10.38 | <0.0001 | 1 | 10.15 | 10.15 | 11.41 | 0.0018 |
| 10 | 53 | 822.94 | 15.53 | 14.95 | <0.0001 | 3 | 516.15 | 172.049 | 165.66 | <0.0001 | 1 | 91.85 | 91.85 | 88.45 | <0.0001 |
| 11 | 53 | 98,712.64 | 1862.50 | 391.96 | <0.0001 | 3 | 1092.09 | 364.03 | 76.61 | <0.0001 | 1 | 6392.80 | 6392.80 | 1345.35 | <0.0001 |
| 12 | 53 | 377.84 | 7.13 | Infini | <0.0001 | 3 | 288.00 | 96.00 | Infini | <0.0001 | 1 | 82.51 | 82.51 | Infini | <0.0001 |
| 13 | 53 | 8853.77 | 167.05 | 33.04 | <0.0001 | 3 | 5.64 | 1.88 | 0.37 | 0.7736 | 1 | 1092.99 | 1092.99 | 216.20 | <0.0001 |
| 14 | 53 | 2595.73 | 48.98 | 19.24 | <0.0001 | 3 | 59.56 | 19.85 | 7.80 | 0.0004 | 1 | 379.86 | 379.86 | 149.26 | <0.0001 |
| 15 | 53 | 103,443,899.9 | 1,951,771.7 | 22.35 | <0.0001 | 3 | 13,494,835.8 | 4,498,278.6 | 51.52 | <0.0001 | 1 | 5,333,366.9 | 5,333,366.9 | 61.08 | <0.0001 |
| (a) | |||||||||||||||
| Traits | ANOVA | Type III Sum of Square Analysis | |||||||||||||
| Model | Block Effect | Accession Effect | |||||||||||||
| DF | Sum of Squares | Mean Squares | F | Pr > F | DF | Sum of Squares | Mean Squares | F | p | DF | Sum of Squares | Mean Squares | F | p | |
| 1 | 172 | 2564.76 | 14.91 | Infini | <0.0001 | 7 | 0.00 | 0.00 | 165 | 1217.73 | 7.38 | Infini | <0.0001 | ||
| 2 | 172 | 2986.01 | 17.36 | 11.86 | <0.0001 | 7 | 43.03 | 6.15 | 4.20 | 0.0019 | 165 | 1796.78 | 10.89 | 7.44 | <0.0001 |
| 3 | 172 | 829.12 | 4.82 | 2.99 | 0.0001 | 7 | 18.36 | 2.62 | 1.63 | 0.1600 | 165 | 817.33 | 4.95 | 3.07 | 0.0001 |
| 4 | 172 | 2121.73 | 12.33 | 6.57 | <0.0001 | 7 | 77.10 | 11.01 | 5.86 | 0.0001 | 165 | 2024.37 | 12.27 | 6.53 | <0.0001 |
| 5 | 172 | 45,758.01 | 266.03 | 233.27 | <0.0001 | 7 | 4.33 | 0.62 | 0.54 | 0.80 | 165 | 44,869.16 | 271.93 | 238.44 | <0.0001 |
| 6 | 172 | 133.09 | 0.77 | 21.64 | <0.0001 | 7 | 0.91 | 0.13 | 3.65 | 0.0046 | 165 | 109.03 | 0.66 | 18.48 | <0.0001 |
| 7 | 172 | 4330.26 | 25.18 | 8.55 | <0.0001 | 7 | 22.38 | 3.20 | 1.09 | 0.3929 | 165 | 3090.50 | 18.73 | 6.36 | <0.0001 |
| 8 | 172 | 17,010.12 | 98.90 | 68.36 | <0.0001 | 7 | 27.21 | 3.89 | 2.69 | 0.0245 | 165 | 13,619.42 | 82.54 | 57.05 | <0.0001 |
| 9 | 172 | 748.83 | 4.35 | 4.82 | <0.0001 | 7 | 10.10 | 1.57 | 1.74 | 0.1314 | 165 | 538.18 | 3.26 | 3.61 | <0.0001 |
| 10 | 172 | 4792.84 | 27.87 | 27.94 | <0.0001 | 7 | 12.72 | 1.817 | 1.82 | 0.1139 | 165 | 3740.65 | 22.67 | 22.73 | <0.0001 |
| 11 | 172 | 60,475.56 | 351.60 | 365.40 | <0.0001 | 7 | 16.57 | 2.37 | 2.46 | 0.0367 | 165 | 44,943.53 | 272.39 | 283.07 | <0.0001 |
| 12 | 172 | 3387.31 | 19.69 | 236.32 | <0.0001 | 7 | 0.58 | 0.08 | 1.00 | 0.4478 | 165 | 2581.24 | 15.64 | 187.73 | <0.0001 |
| 13 | 172 | 23,536.50 | 136.84 | 29.99 | <0.0001 | 7 | 44.61 | 6.37 | 1.40 | 0.2377 | 165 | 21,579.47 | 130.78 | 28.66 | <0.0001 |
| 14 | 172 | 4993.97 | 29.03 | 14.40 | <0.0001 | 7 | 48.15 | 6.88 | 3.41 | 0.0070 | 165 | 4836.06 | 29.31 | 14.54 | <0.0001 |
| 15 | 172 | 182,274,678.9 | 1,059,736.5 | 15.32 | <0.0001 | 7 | 1,589,762.2 | 227,108.9 | 3.28 | 0.0087 | 165 | 177,020,077.70 | 1,072,849.00 | 15.51 | <0.0001 |
| (b) | |||||||||||||||
| Traits | Contrast (Differences) | ||||||||||||||
| Among Accessions | Among Checks | Check vs. Accession | |||||||||||||
| DF | Sum of Squares | Mean Squares | F | Pr > F | DF | Sum of Squares | Mean Squares | F | p | DF | Sum of Squares | Mean Squares | F | p | |
| 1 | 53 | 67.20 | 1.27 | Infini | <0.0001 | 3 | 16.00 | 5.33 | Infini | <0.0001 | 1 | 118.26 | 118.26 | Infini | <0.0001 |
| 2 | 53 | 472.30 | 8.91 | 6.09 | <0.0001 | 3 | 96.14 | 32.04 | 21.89 | <0.0001 | 1 | 59.07 | 59.07 | 40.35 | <0.0001 |
| 3 | 53 | 219.95 | 4.15 | 2.58 | 0.0019 | 3 | 34.07 | 11.36 | 7.05 | 0.0008 | 1 | 158.18 | 158.18 | 98.16 | <0.0001 |
| 4 | 53 | 827.02 | 15.60 | 8.31 | <0.0001 | 3 | 141.08 | 47.03 | 25.03 | <0.0001 | 1 | 364.33 | 364.33 | 193.94 | <0.0001 |
| 5 | 53 | 17,480.49 | 329.82 | 289.20 | <0.0001 | 3 | 58.84375 | 19.61 | 17.20 | <0.0001 | 1 | 872.40 | 872.40 | 764.95 | <0.0001 |
| 6 | 53 | 25.28 | 0.47 | 13.34 | <0.0001 | 3 | 0.41 | 0.14 | 3.84 | 0.0178 | 1 | 0.42 | 0.42 | 11.70 | 0.0016 |
| 7 | 53 | 805.51 | 15.20 | 5.16 | <0.0001 | 3 | 125.78 | 41.93 | 14.25 | <0.0001 | 1 | 169.17 | 169.17 | 57.48 | <0.0001 |
| 8 | 53 | 1899.80 | 35.84 | 24.78 | <0.0001 | 3 | 255.25 | 85.08 | 58.81 | <0.0001 | 1 | 1857.71 | 1857.71 | 1284.07 | <0.0001 |
| 9 | 53 | 138.13 | 2.61 | 2.89 | 0.0006 | 3 | 29.55375 | 9.85 | 10.92 | <0.0001 | 1 | 12.05 | 12.05 | 13.35 | 0.0008 |
| 10 | 53 | 790.35 | 14.91 | 14.95 | <0.0001 | 3 | 495.71 | 165.24 | 165.66 | <0.0001 | 1 | 88.22 | 88.22 | 88.45 | <0.0001 |
| 11 | 53 | 19,989.31 | 377.16 | 391.96 | <0.0001 | 3 | 221.15 | 73.72 | 76.61 | <0.0001 | 1 | 1294.54 | 1294.54 | 1345.35 | <0.0001 |
| 12 | 53 | 377.20 | 7.12 | 85.40 | <0.0001 | 3 | 276.38 | 92.13 | 1105.50 | <0.0001 | 1 | 78.07 | 78.070 | 936.85 | <0.0001 |
| 13 | 53 | 7990.53 | 150.76 | 33.04 | <0.0001 | 3 | 5.093673 | 1.70 | 0.37 | 0.7736 | 1 | 986.42 | 986.43 | 216.20 | <0.0001 |
| 14 | 53 | 2134.87 | 40.28 | 19.98 | <0.0001 | 3 | 47.18 | 15.73 | 7.80 | 0.0004 | 1 | 324.85 | 324.85 | 161.15 | <0.0001 |
| 15 | 53 | 83,831,969.77 | 1,581,735.28 | 22.87 | <0.0001 | 3 | 10,689,259.48 | 3,563,086.49 | 51.52 | <0.0001 | 1 | 4,556,054.50 | 4,556,054.50 | 65.87 | <0.0001 |
| (a) | |||||||||||||||
| Traits | ANOVA | Type III Sum of Square Analysis | |||||||||||||
| Model | Block Effect | AccessionEffect | |||||||||||||
| DF | Sum of Squares | Mean Squares | F | Pr > F | DF | Sum of Squares | Mean Squares | F | p | DF | Sum of Squares | Mean Squares | F | p | |
| 1 | 172 | 1886.23 | 10.97 | Infini | <0.0001 | 7 | 0.00 | 0.00 | 165 | 1236.69 | 7.50 | Infini | <0.0001 | ||
| 2 | 172 | 2312.36 | 13.44 | 11.86 | <0.0001 | 7 | 33.32 | 4.76 | 4.20 | 0.0019 | 165 | 1391.43 | 8.43 | 7.44 | <0.0001 |
| 3 | 172 | 627.56 | 3.65 | 2.99 | 0.0001 | 7 | 13.90 | 1.99 | 1.63 | 0.1600 | 165 | 618.64 | 3.75 | 3.07 | 0.0001 |
| 4 | 172 | 2185.86 | 12.71 | 6.57 | <0.0001 | 7 | 79.43 | 11.35 | 5.86 | 0.0001 | 165 | 2085.55 | 12.64 | 6.53 | <0.0001 |
| 5 | 172 | 45,758.01 | 266.03 | 233.27 | <0.0001 | 7 | 4.33 | 0.62 | 0.54 | 0.7960 | 165 | 44,869.16 | 271.93 | 238.44 | <0.0001 |
| 6 | 172 | 155.23 | 0.90 | 21.64 | <0.0001 | 7 | 1.07 | 0.15 | 3.65 | 0.0046 | 165 | 127.18 | 0.77 | 18.48 | <0.0001 |
| 7 | 172 | 108,256.43 | 629.40 | 8.55 | <0.0001 | 7 | 559.500 | 79.93 | 1.09 | 0.3929 | 165 | 77,262.23 | 468.26 | 6.36 | <0.0001 |
| 8 | 172 | 18,046.03 | 104.92 | 68.36 | <0.0001 | 7 | 28.86 | 4.12 | 2.69 | 0.0245 | 165 | 14,448.84 | 87.57 | 57.05 | <0.0001 |
| 9 | 172 | 1643.58 | 9.56 | 4.62 | <0.0001 | 7 | 26.73 | 3.82 | 1.84 | 0.1094 | 165 | 1181.19 | 7.16 | 3.46 | <0.0001 |
| 10 | 172 | 4918.26 | 28.59 | 27.94 | <0.0001 | 7 | 13.05 | 1.86 | 1.82 | 0.1139 | 165 | 3838.54 | 23.26 | 22.73 | <0.0001 |
| 11 | 172 | 216,024.75 | 1255.96 | 365.40 | <0.0001 | 7 | 59.18 | 8.45 | 2.46 | 0.0367 | 165 | 160,542.80 | 972.99 | 283.07 | <0.0001 |
| 12 | 172 | 4479.72 | 26.04 | 236.32 | <0.0001 | 7 | 0.77 | 0.11 | 1.00 | 0.4478 | 165 | 3413.69 | 20.69 | 187.73 | <0.0001 |
| 13 | 172 | 24,009.58 | 139.59 | 29.99 | <0.0001 | 7 | 45.51 | 6.50 | 1.40 | 0.2377 | 165 | 22,013.22 | 133.41 | 28.66 | <0.0001 |
| 14 | 172 | 5554.90 | 32.30 | 14.06 | <0.0001 | 7 | 54.86 | 7.84 | 3.41 | 0.0070 | 165 | 5345.55 | 32.40 | 14.11 | <0.0001 |
| 15 | 172 | 203,243,103.1 | 1,181,645.9 | 14.99 | <0.0001 | 7 | 1,811,337.5 | 258,762.5 | 3.28 | 0.0087 | 165 | 196,745,949.9 | 1,192,399.7 | 15.13 | <0.0001 |
| (b) | |||||||||||||||
| Traits | Contrast (Differences) | ||||||||||||||
| Among Accessions | Among Checks | Check vs. Accession | |||||||||||||
| DF | Sum of Squares | Mean Squares | F | Pr > F | DF | Sum of Squares | Mean Squares | F | p | DF | Sum of Squares | Mean Squares | F | p | |
| 1 | 53 | 44.13 | 0.83 | Infini | <0.0001 | 3 | 16.00 | 5.33 | Infini | <0.0001 | 1 | 103.45 | 103.45 | Infini | <0.0001 |
| 2 | 53 | 365.75 | 6.90 | 6.09 | <0.0001 | 3 | 74.45 | 24.82 | 21.89 | <0.0001 | 1 | 45.74 | 45.74 | 40.35 | <0.0001 |
| 3 | 53 | 166.48 | 3.14 | 2.58 | 0.0019 | 3 | 25.78 | 8.59 | 7.05 | 0.0008 | 1 | 119.73 | 119.73 | 98.16 | <0.0001 |
| 4 | 53 | 852.01 | 16.08 | 8.31 | <0.0001 | 3 | 145.34 | 48.45 | 25.03 | <0.0001 | 1 | 375.34 | 375.34 | 193.94 | <0.0001 |
| 5 | 53 | 17,480.49 | 329.82 | 289.20 | <0.0001 | 3 | 58.84 | 19.61 | 17.20 | <0.0001 | 1 | 872.40 | 872.40 | 764.95 | <0.0001 |
| 6 | 53 | 29.49 | 0.56 | 13.34 | <0.0001 | 3 | 0.48 | 0.16 | 3.84 | 0.0178 | 1 | 0.49 | 0.49 | 11.70 | 0.0016 |
| 7 | 53 | 20,137.78 | 379.96 | 5.16 | <0.0001 | 3 | 3144.59 | 1048.20 | 14.25 | <0.0001 | 1 | 4229.18 | 4229.18 | 57.48 | <0.0001 |
| 8 | 53 | 2015.50 | 38.03 | 24.78 | <0.0001 | 3 | 270.79 | 90.26 | 58.81 | <0.0001 | 1 | 1970.84 | 1970.84 | 1284.07 | <0.0001 |
| 9 | 53 | 295.70 | 5.58 | 2.70 | 0.0013 | 3 | 63.23 | 21.08 | 10.18 | <0.0001 | 1 | 25.27 | 25.27 | 12.21 | 0.0013 |
| 10 | 53 | 811.04 | 15.30 | 14.95 | <0.0001 | 3 | 508.68 | 169.56 | 165.66 | <0.0001 | 1 | 90.53 | 90.53 | 88.45 | <0.0001 |
| 11 | 53 | 71,403.81 | 1347.24 | 391.96 | <0.0001 | 3 | 789.97 | 263.32 | 76.61 | <0.0001 | 1 | 4624.23 | 4624.23 | 1345.35 | <0.0001 |
| 12 | 53 | 498.84 | 9.41 | 85.40 | <0.0001 | 3 | 365.51 | 121.84 | 1105.50 | <0.0001 | 1 | 103.25 | 103.25 | 936.85 | <0.0001 |
| 13 | 53 | 8151.14 | 153.80 | 33.04 | <0.0001 | 3 | 5.20 | 1.70 | 0.37 | 0.7736 | 1 | 1006.25 | 1006.25 | 216.20 | <0.0001 |
| 14 | 53 | 2342.65 | 44.20 | 19.24 | <0.0001 | 3 | 53.76 | 17.92 | 7.80 | 0.0004 | 1 | 342.83 | 342.83 | 149.26 | <0.0001 |
| 15 | 53 | 93,358,119.63 | 1,761,473.96 | 22.35 | <0.0001 | 3 | 12,179,089.35 | 4,059,696.45 | 51.52 | <0.0001 | 1 | 4,813,363.60 | 4,813,363.60 | 61.08 | <0.0001 |
| S/N | Traits | p-Values for Site Effects | p-Values for Season Effects | ||||
|---|---|---|---|---|---|---|---|
| Site | Accession | Site x Accession | Season | Accession | Accession x Season | ||
| 1 | Germination time (days) | <0.0001 | 0.000 | 0.153 | 0.097 | 0.000 | 0.979 |
| 2 | Terminal leaflet length (cm) | <0.0001 | <0.0001 | 1.000 | <0.0001 | <0.0001 | 0.961 |
| 3 | Terminal leaflet width (cm) | 0.000 | <0.0001 | 1.000 | <0.0001 | <0.0001 | 0.998 |
| 4 | Petiole length (cm) | 0.000 | <0.0001 | 1.000 | 0.009 | <0.0001 | 1.000 |
| 5 | Days to flowering | <0.0001 | <0.0001 | 0.032 | <0.0001 | <0.0001 | 1.000 |
| 6 | Flower bud size (cm) | <0.0001 | <0.0001 | 0.078 | <0.0001 | <0.0001 | 0.899 |
| 7 | Number of flowers per raceme | <0.0001 | <0.0001 | 0.995 | <0.0001 | 0.000 | 0.003 |
| 8 | Peduncle length (cm) | 0.003 | <0.0001 | 1.000 | 0.003 | <0.0001 | 1.000 |
| 9 | Pods per peduncle | 0.742 | <0.0001 | 0.973 | <0.0001 | <0.0001 | 0.054 |
| 10 | Pod length (cm) | 0.194 | <0.0001 | 1.000 | 0.371 | <0.0001 | 1.000 |
| 11 | Pods per plant | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| 12 | Seeds per pod | 0.894 | <0.0001 | 1.000 | <0.0001 | <0.0001 | 0.712 |
| 13 | Seed size (mm2) | <0.0001 | <0.0001 | 0.052 | 0.013 | <0.0001 | 0.506 |
| 14 | 100-Seed weight (g) | <0.0001 | <0.0001 | 0.037 | <0.0001 | <0.0001 | 0.068 |
| 15 | Yield (Kg/ha) | <0.0001 | <0.0001 | 0.032 | <0.0001 | <0.0001 | 0.055 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harouna, D.V.; Venkataramana, P.B.; Matemu, A.O.; Ndakidemi, P.A. Agro-Morphological Exploration of Some Unexplored Wild Vigna Legumes for Domestication. Agronomy 2020, 10, 111. https://doi.org/10.3390/agronomy10010111
Harouna DV, Venkataramana PB, Matemu AO, Ndakidemi PA. Agro-Morphological Exploration of Some Unexplored Wild Vigna Legumes for Domestication. Agronomy. 2020; 10(1):111. https://doi.org/10.3390/agronomy10010111
Chicago/Turabian StyleHarouna, Difo Voukang, Pavithravani B. Venkataramana, Athanasia O. Matemu, and Patrick Alois Ndakidemi. 2020. "Agro-Morphological Exploration of Some Unexplored Wild Vigna Legumes for Domestication" Agronomy 10, no. 1: 111. https://doi.org/10.3390/agronomy10010111