
Application of Chitinous Materials in Production and Purification of a Poly(l-lactic acid) Depolymerase from Pseudomonas tamsuii TKU015



Abstract
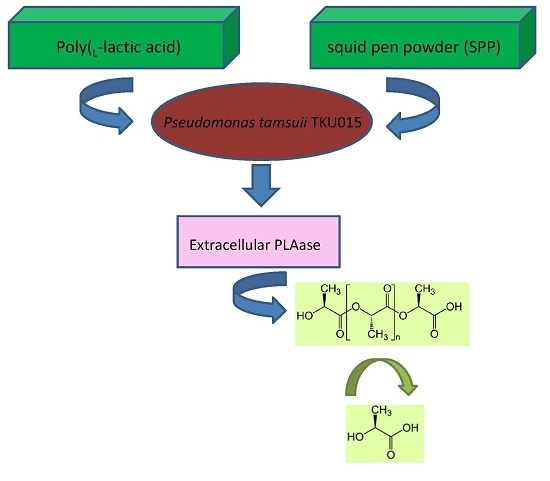
:
1. Introduction
2. Experimental
2.1. Materials
2.2. Strains
2.3. Screening of PLA-Degrading Microorganisms
2.4. Investigation of Flask Culture Conditions
2.5. PLA-Degrading Activity Assays
2.6. Purification of PLA-Degrading Enzyme
2.7. Protein Determination
2.8. Sodium Dodecyl Sulphate-Polyacrylamide Gel Electrophoresis
2.9. Effects of pH and Temperature on PLA-Degrading Activity
2.10. Activities of the Purified PLA Depolymerase for Various Substrates
3. Results and Discussion
3.1. Selection of a PLA Depolymerase-Producing Strain
3.2. Culture Conditions for PLA Depolymerase Production
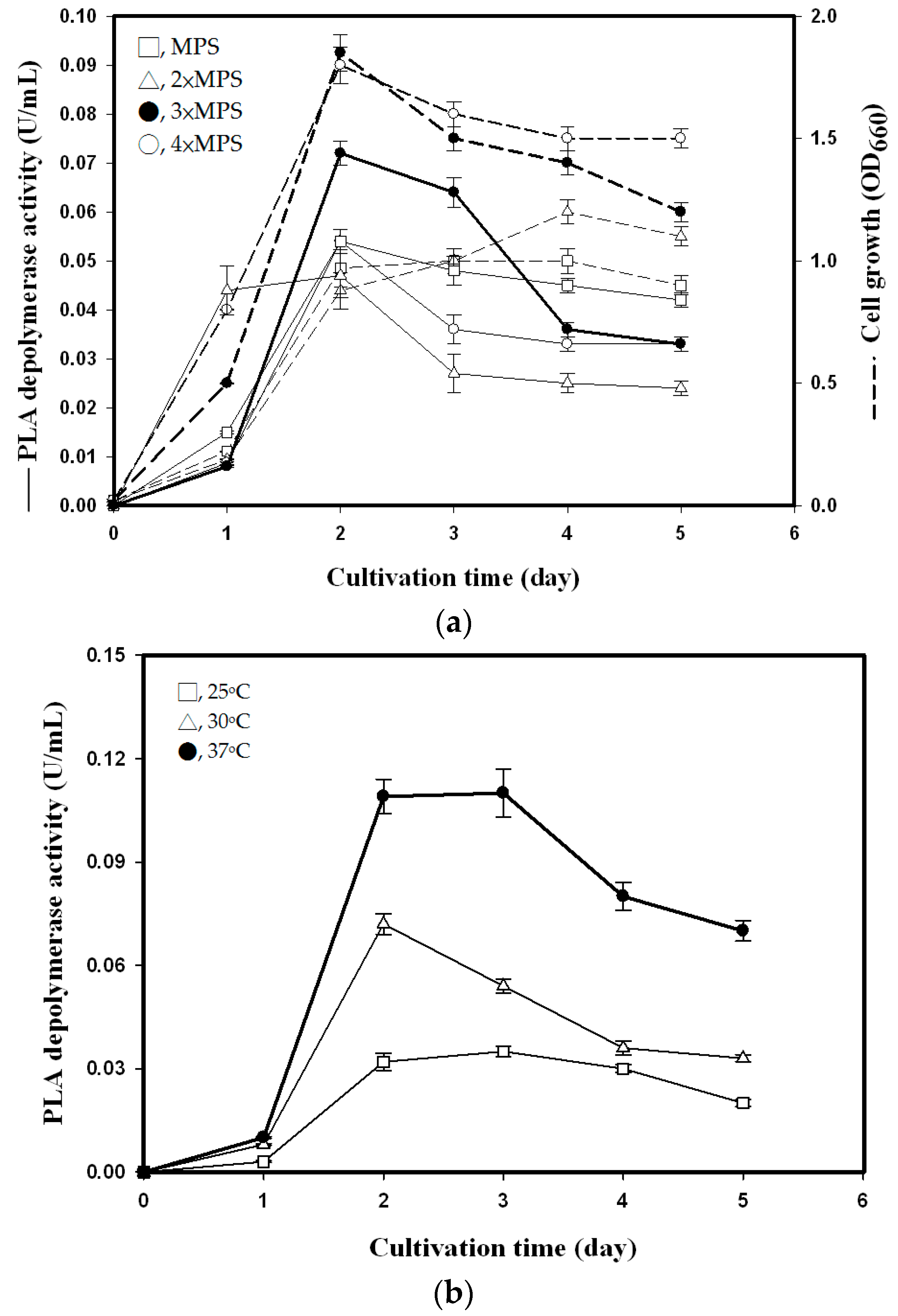
3.2.1. Effect of MPS Concentration
3.2.2. Effect of Culture Volume, Initial pH, and Temperature
3.2.3. Time Course of PLA Depolymerase Production
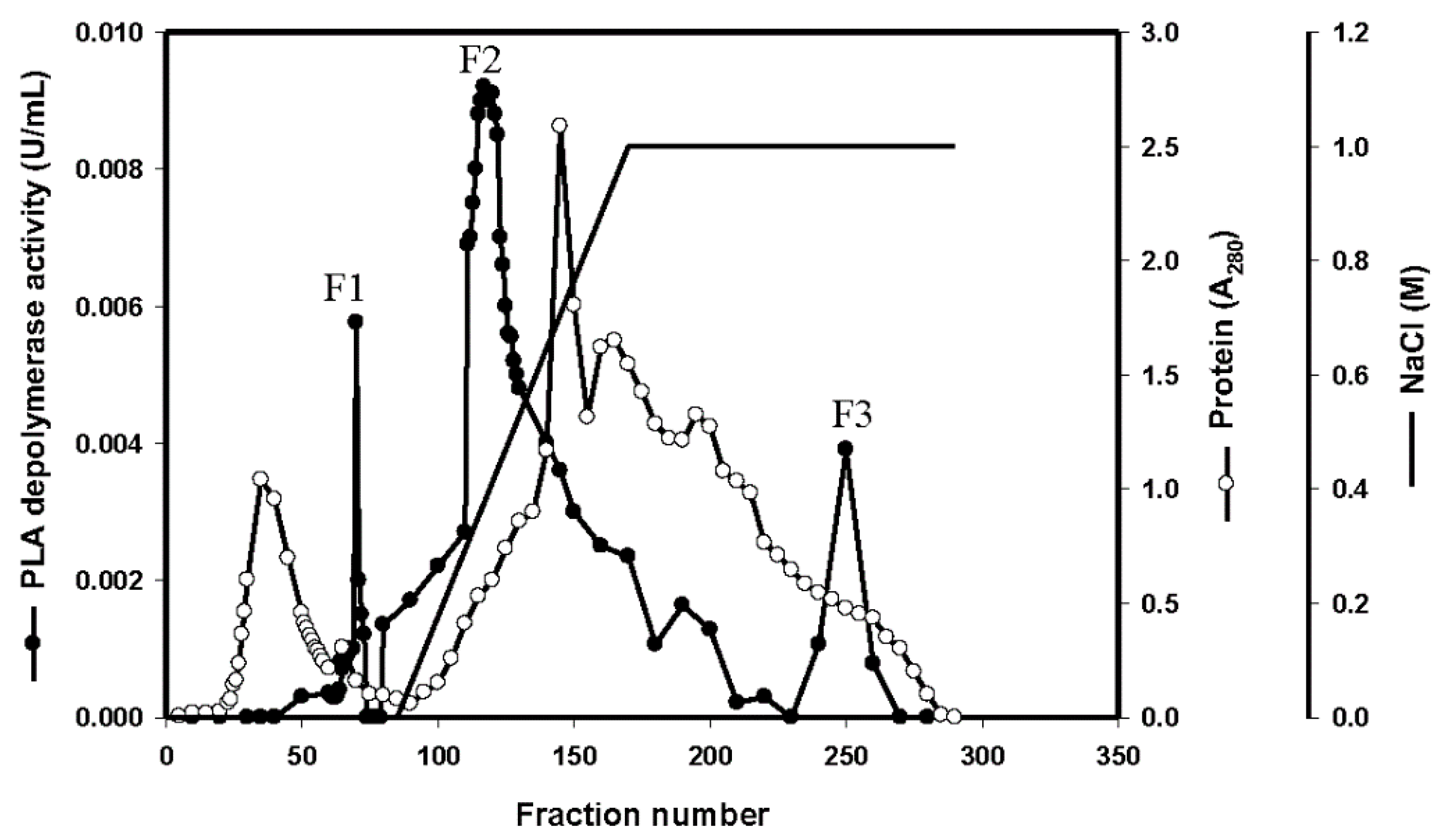
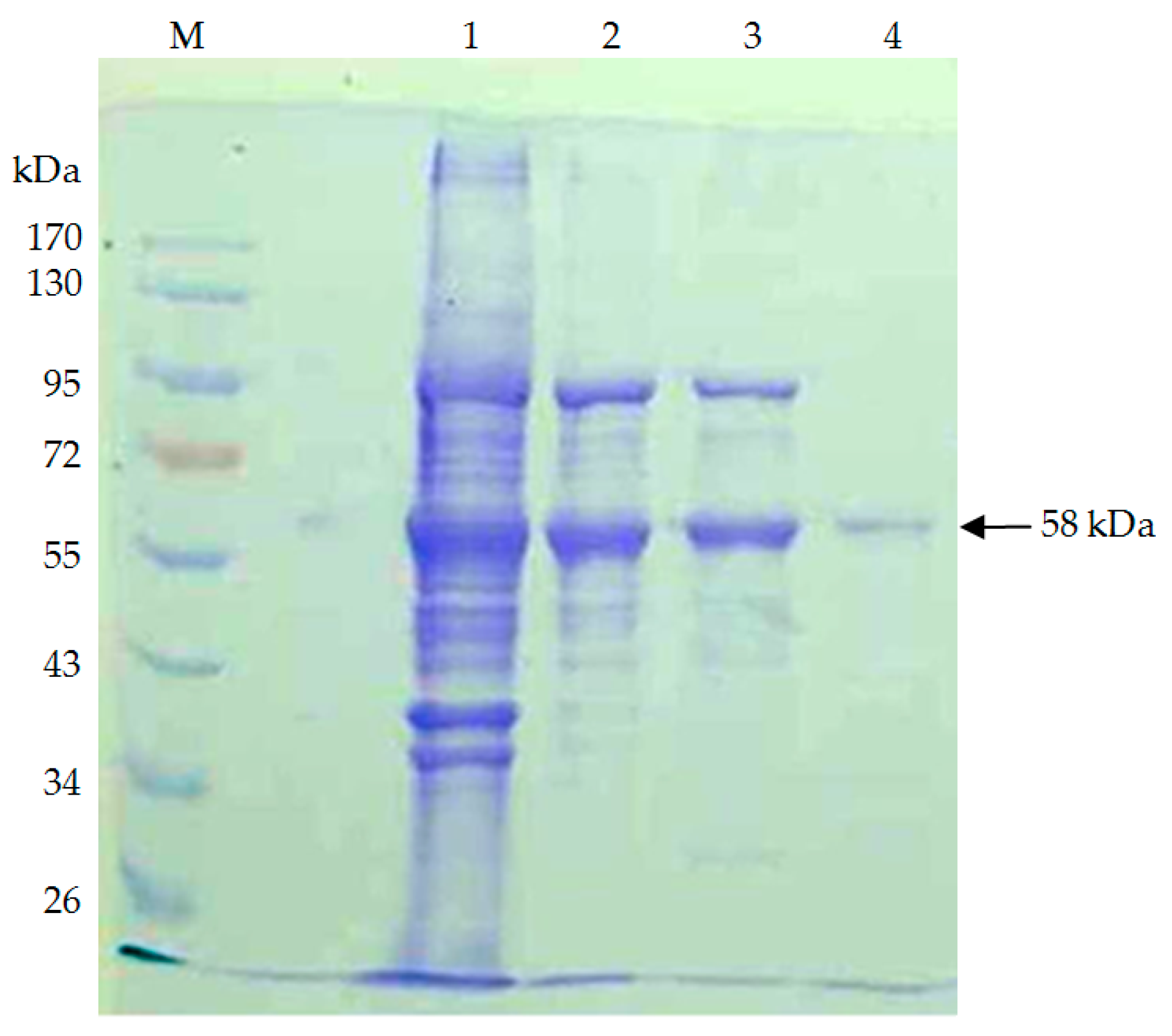
3.3. Purification of the Enzyme
3.4. Effects of pH and Temperature
3.5. Substrate Specificity
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wang, Z.; Wang, Y.; Guo, Z.; Li, F.; Chen, S. Purification and characterization of poly(l-lactic acid) depolymerase from Pseudomonas sp. strain DS04-T. Polym. Eng. Sci. 2011, 51, 454–459. [Google Scholar] [CrossRef]
- Fukuzaki, H.; Yoshida, M.; Asano, M.; Kumakura, M. Synthesis of copoly(d,l-lactic acid) with relatively low molecular weight and in vitro degradation. Eur. Polym. J. 1989, 25, 1019–1026. [Google Scholar] [CrossRef]
- Reeve, S.M.; McCarthy, S.P.; Downey, M.J.; Gross, R.A. Polylactide stereochemistry: Effect on enzymatic degradability. Macromolecules 1994, 27, 825–831. [Google Scholar] [CrossRef]
- Williams, D.F. Enzymic hydrolysis of polylactic acid. Eng. Med. 1981, 10, 5–7. [Google Scholar] [CrossRef]
- Jarerat, A.; Tokiwa, Y.; Tanaka, H. Production of poly(l-lactide)-degrading enzyme by Amycolatopsis orientalis for biological recycling of poly(l-lactide). Appl. Microbiol. Biotechnol. 2006, 72, 726–731. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, A.; Isono, Y. Degradation of aliphatic polyester films by commercially available lipases with special reference to rapid and complete degradation of poly(l-lactide) film by lipase PL derived from Alcaligenes sp. Biodegradation 2002, 13, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Akutsu-Shigeno, Y.; Teeraphatpornchai, T.; Teamtisong, K.; Nomura, N.; Uchiyama, H.; Nakahara, T.; Nakajima-Kambe, T. Cloning and sequencing of a poly(d,l-lactic acid) depolymerase gene from Paenibacillus amylolyticus strain TB-13 and its functional expression in Escherichia coli. Appl. Environ. Microbiol. 2003, 69, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Masaki, K.; Kamini, N.R.; Ikeda, H.; Iefuji, H. Cutinase-like enzyme from the yeast Cryptococcus sp. strain S-2 hydrolyzes polylactic acid and other biodegradable plastics. Appl. Environ. Microbiol. 2005, 71, 7548–7550. [Google Scholar] [CrossRef] [PubMed]
- Akutsu, H.; Nakajima-Kambe, T.; Nomura, N.; Nakahara, T. Purification and properties of a polyester polyurethane-degrading enzyme from Comamonas acidovorans TB-35. Appl. Environ. Microbiol. 1998, 64, 62–67. [Google Scholar] [PubMed]
- Tokiwa, Y.; Calabia, B.P. Biodegradability and biodegradation of poly(lactide). Appl. Microbiol. Biotechnol. 2006, 72, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Pranamuda, H.; Tokiwa, Y.; Tanaka, H. Polylactide degradation by an Amycolatopsis sp. Appl. Environ. Microbiol. 1997, 63, 1637–1640. [Google Scholar] [PubMed]
- Pranamuda, H.; Tokiwa, Y. Degradation of poly(l-lactide) by strains belonging to genus Amycolatopsis. Biotechnol. Lett. 1999, 21, 901–905. [Google Scholar] [CrossRef]
- Nakamura, K.; Tomita, T.; Abe, N.; Kamio, Y. Purification and characterization of an extracellular poly(l-lactic acid) depolymerase from a soil isolate, Amycolatopsis sp. strain K104–1. Appl. Environ. Microbiol. 2001, 67, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Pranamuda, H.; Tsuchii, A.; Tokiwa, Y. Poly (l-lactide)-degrading enzyme produced by Amycolatopsis sp. Macromol. Biosci. 2001, 1, 25–29. [Google Scholar] [CrossRef]
- Jarerat, A.; Pranamuda, H.; Tokiwa, Y. Poly(l-lactide)-degrading activity in various actinomycetes. Macromol. Biosci. 2002, 2, 420–428. [Google Scholar] [CrossRef]
- Wang, S.L.; Chen, S.J.; Wang, C.L. Purification and characterization of chitinases and chitosanases from a new species strain Pseudomonas sp. TKU015 using shrimp shells as a substrate. Carbohydr. Res. 2008, 343, 1171–1179. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.L.; Chen, H.J.; Liang, T.W.; Lin, Y.D. A novel nattokinase produced by Pseudomonas sp. TKU015 using shrimp shells as substrate. Process Biochem. 2009, 44, 70–76. [Google Scholar] [CrossRef]
- Liang, T.W.; Chen, C.H.; Wang, S.L. Production of insecticidal materials from Pseudomonas tamsuii. Res. Chem. Intermed. 2015, 41, 7965–7971. [Google Scholar] [CrossRef]
- Wang, S.L.; Lin, T.Y.; Yen, Y.H.; Liao, H.F.; Chen, Y.J. Bioconversion of shellfish chitin wastes for the production of Bacillus subtilis W-118 chitinase. Carbohydr. Res. 2006, 341, 2507–2515. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; Yamagata, Y.; Abe, K.; Hasegawa, F.; Machida, M.; Ishioka, R.; Gomi, K.; Nakajima, T. Purification and characterization of a biodegradable plastic-degrading enzyme from Aspergillus oryzae. Appl. Microbiol. Biotechnol. 2005, 67, 778–788. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, K.; Yoshida, Y.; Toida, J.; Sekiguchi, J. Purification and characterization of a novel lipolytic enzyme from Aspergillus oryzae. J. Ferment. Bioeng. 1994, 78, 413–419. [Google Scholar] [CrossRef]
- Teeraphatpornchai, T.; Nakajima-Kambe, T.; Shigeno-Akutsu, Y.; Nakayama, M.; Nomura, N.; Nakahara, T.; Uchiyama, H. Isolation and characterization of a bacterium that degrades various polyester-based biodegradable plastics. Biotechnol. Lett. 2003, 25, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Shinozaki, Y.; Yoshida, S.; Koitabashi, M.; Sameshima-Yamashita, Y.; Fujii, T.; Fukuoka, T.; Kitamoto, H.K. Xylose induces the phyllosphere yeast Pseudozyma antarctica to produce a cutinase-like enzyme which efficiently degrades biodegradable plastics. J. Biosci. Bioeng. 2014, 117, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Wang, S.; Liu, W.; Chen, G. Purification and characterization of poly(l-lactic acid)-degrading enzymes from Amycolatopsis orientalis ssp. orientalis. FEMS Microbiol. Lett. 2008, 282, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Jarerat, A.; Tokiwa, Y. Degradation of poly(L-lactide) by a fungus. Macromol. Biosci. 2001, 1, 136–140. [Google Scholar] [CrossRef]
- Torres, A.; Li, S.M.; Roussos, S.; Vert, M. Screening of microorganisms for biodegradation of poly(lactic acid) and lactic acid-containing polymers. Appl. Environ. Microbiol. 1996, 62, 2392–2397. [Google Scholar]
- Watanabe, T.; Shinozaki, Y.; Suzuki, K.; Koitabashi, M.; Yoshida, S.; Sameshima-Yamashita, Y.; Kitamoto, H.K. Production of a biodegradable plastic-degrading enzyme from cheese whey by the phyllosphere yeast Pseudozyma antarctica GB-4(1)W. J. Biosci. Bioeng. 2014, 118, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Suzuki, K.; Shinozaki, Y.; Yarimizu, T.; Yoshida, S.; Sameshima-Yamashita, Y.; Koitabashi, M.; Kitamoto, H.K. A UV-induced mutant of Cryptococcus flavus GB-1 with increased production of a biodegradable plastic-degrading enzyme. Process Biochem. 2015, 50, 1718–1724. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Step | Total Protein (mg) | Total Activity (U) | Specific Activity (U/mg Protein) | Purification Fold | Yield (%) |
|---|---|---|---|---|---|
| Culture supernatant | 1425.0 | 3.6 | 0.003 | 1.0 | 100 |
| (NH4)2SO4 ppt | 220.1 | 0.9 | 0.004 | 1.3 | 25 |
| DEAE-Sepharose | 42.1 | 0.4 | 0.010 | 3.3 | 11 |
| Macro-prep DEAE | 35.5 | 0.4 | 0.011 | 3.7 | 11 |
| Sephacryl S-100 | 10.8 | 0.3 | 0.028 | 9.3 | 8 |
| Substrate | Hydrolysis Activity (U/mg) |
|---|---|
| Casein | <0.10 ± 0.002 c |
| Fibrinogen | 7.22 ± 0.25 a |
| Poly(β-hydroxybutyrate) | <0.10 ± 0.001 c |
| Tributyrin | 3.51 ± 0.16 b |
| Triolein | <0.10 ± 0.001 c |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, T.-W.; Jen, S.-N.; Nguyen, A.D.; Wang, S.-L. Application of Chitinous Materials in Production and Purification of a Poly(l-lactic acid) Depolymerase from Pseudomonas tamsuii TKU015. Polymers 2016, 8, 98. https://doi.org/10.3390/polym8030098
Liang T-W, Jen S-N, Nguyen AD, Wang S-L. Application of Chitinous Materials in Production and Purification of a Poly(l-lactic acid) Depolymerase from Pseudomonas tamsuii TKU015. Polymers. 2016; 8(3):98. https://doi.org/10.3390/polym8030098
Chicago/Turabian StyleLiang, Tzu-Wen, Shan-Ni Jen, Anh Dzung Nguyen, and San-Lang Wang. 2016. "Application of Chitinous Materials in Production and Purification of a Poly(l-lactic acid) Depolymerase from Pseudomonas tamsuii TKU015" Polymers 8, no. 3: 98. https://doi.org/10.3390/polym8030098