
Manipulating Microbial Cell Morphology for the Sustainable Production of Biopolymers

 , , ,
, , ,
Abstract
:1. Introduction
2. Biopolymers
2.1. Polyhydroxyalkanoates
2.2. Polylactides
2.3. Polycaprolactones
2.4. Other Polymers
3. Sustainable Production of Biopolymers
3.1. Bioengineering of Microbial Cells
3.1.1. Cell Morphology
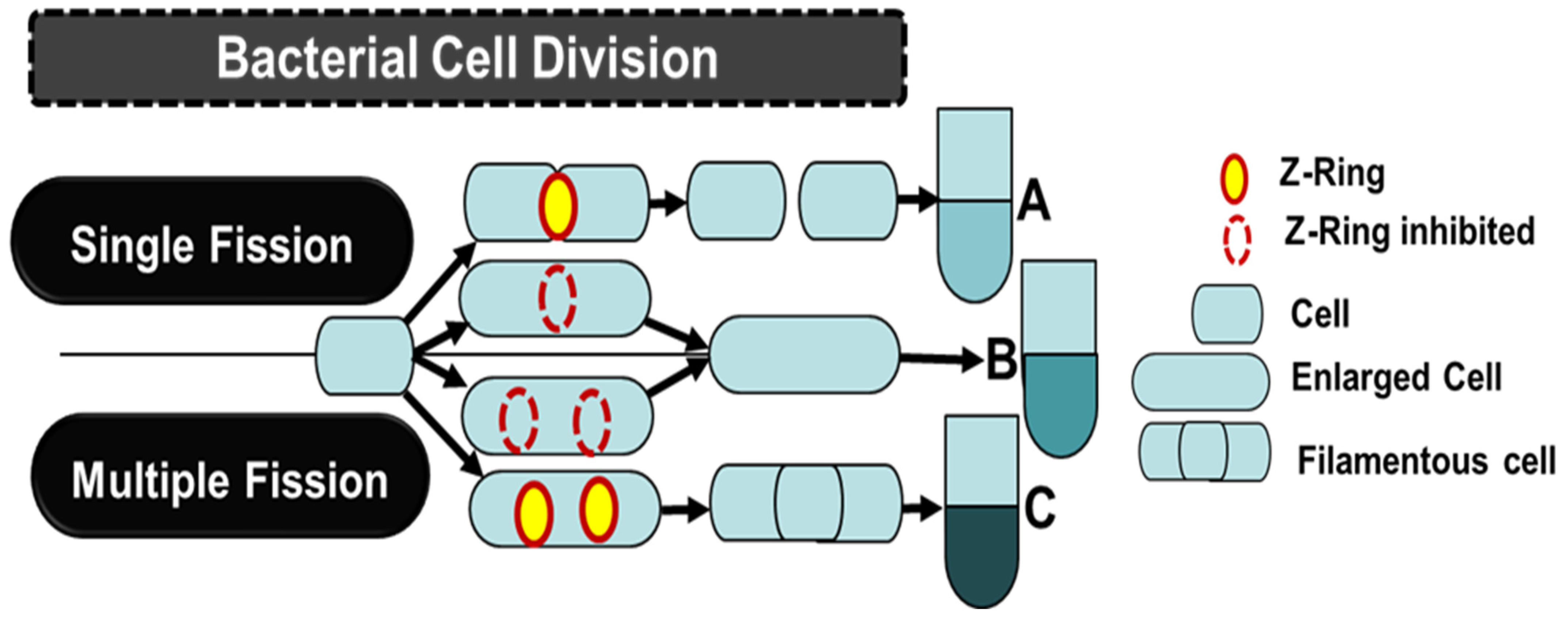
3.1.2. Cell Division

{kind=link}
| Gene | Gene Product | Cell Function/Activity | Reference |
|---|---|---|---|
| dxs | 1-deoxyxylulose-5-phosphate synthase | Cell wall synthesis | [65] |
| glmU | N-acetylglucosamine-1-phosphate uridyltransferase | Cell wall synthesis | |
| murA | UDP- N -acetylglucosamine enolpyruvoyl transferase | Cell wall synthesis | |
| murC | UDP-N-acetylmuramate-alanine ligase | Cell wall synthesis | |
| murD | UDP-N-acetylmuramoyl-L-alanine:D-glutamate ligase | Cell wall synthesis | |
| murE | Ligase | Cell wall synthesis | |
| murJ | Putative lipid II flippase | Regulates peptidoglycan incorporation to the septum | [67] |
| ftsZ | Bacterial fission ring formation protein | Recruiting divisiome proteins and z-ring stabilization | [64,73,74] |
| ftsA | Cell division protein | Divisiome | [69] |
| ftsW | Peptidoglycan glycosyltransferase, lipid II flippase | Divisiome | [65] |
| ftsL, ftsN, ftsQ | Cell division proteins | Divisiome | [67] |
| sulA | Cell division inhibitor protein | Divisiome, induces FtsZ inhibition | [72,75,78] |
| slmA | Nucleoid-associated FtsZ binding protein | Divisiome | [60] |
| minC | Z-ring positioning protein | Divisiome: actin-related proteins, inhibits FtsZ polymerization | [57,76] |
| minD | Z-ring positioning protein | Divisiome: actin-related proteins, recruits MinC | [57,76] |
| envC | Murein hydrolase activator | Divisiome | [74] |
| zipA | Integral inner membrane protein | Divisiome | [62] |
| PBP1, PBP3 | Penicillin binding proteins | Divisiome | [68] |
| envC | Regulate amidase activity | Cell division | [68,77] |
| nlpD | Murein hydrolase activator, peptidoglycan degradation | Cell division | [74,77,78] |
| mreB | Dynamic cytoskeletal protein | Rod complex and cell division | [55,59,72] |
| RodZ | Transmembrane protein | Rod complex | [69] |
| RodA | Transglycosylase, lipid II flippase | Rod complex | [63] |
| PBP2 | Penicillin binding protein, murein DD-transpeptidase | Rod complex, cell elongation | [68] |
| gltA | Citrate synthase | Manipulate cell rigidity | [65] |
| idi | Isopentenyl diphosphate isomerase | Cell wall synthesis | [65] |
| mraY | Translocase 1, phosphor-N-acetylmuramoyl-pentapeptide transferase | Cell wall synthesis | [65] |
| pgi | Phosphoglucose isomerase | Cell wall synthesis | [65] |
| PBP5, PBP7 | DD-carboxypeptidases and DD-endopeptidases | Hydrolases | [61] |
| ampD, amiA, amiB | MurNAc-L-Ala amidases | Hydrolases | [61] |
| Organism | Gene Edited b | Characteristics Affected | Impact on Polyhydroxyalkanoate c Production | Reference |
|---|---|---|---|---|
| Escherichia coli JM109SGIK | sulA | Transformation of rod to filamentous cell with larger internal space | PHB accumulation showed 100% increase | [75] |
| E. coli JM109SGIK | sad, gabD, ispH folk, and sulA | Transformation of rod to filamentous cell with larger internal space | Copolymers of PHA [P(3HB-co-4HB)] were 10% higher (78% in cell dry weight, CDW). | [75] |
| E. coli JM109SG (ΔmreB/pTK-mreB/pBHR68) a | ftsZ, mreB, and sulA | Enlarged cell space due to reduced restriction on space. Larger volume to size ratio. | PHB d production was observed to increase from 5.72 g/L to (9.29 g/L, with a yield of 73.53% of CDW) e in a shake flask | [59] |
| E. coli | envC and nlpD | Switch from binary to multiple fission mode | PHB storing capacity enhanced from 51 to 70% | [74] |
| E. coli JM109 | ftsZ and mreB | Enlarged cell volume | Enhanced PHB accumulations (up to 80%) | [73] |
| E. coli JM109 | ftsW, dxs, glmU, idi, pgi, murA, murC, murD, murE, and mraY | Cell wall thickening | PHB accumulation of 93% in weakened cells and 25% in thickened cell walls | [65] |
| Pseudomonas mendocina NK-01 | ftsZ, mreB, sulA, minCD, and mreB | Modified bacterial shape and growth pattern | Increased mcl-PHA f yield by 45.62% and up to 60.87% | [78] |
| Halomonas bluephagenesis TD08 | minCD | Enlarged cells (1.4-fold longer than the parent) | PHB content enhanced from 69 to 82% | [57] |
| Halomonas campaniensis LS21 | ftsZ and mreB | Enlarged cell morphology | Increase in PHB yield accompanied by normal growth | [72] |
| H. bluephagenesis TDH4-minCD-ΔphaP1 | phaP1, phaP2, phaP3, and minCD | Bigger PHA granules and larger cell size | PHA granules up to 10 μm. PHA copolymer with 14% higher 4HB mol% | [16] |
3.1.3. Cytoskeletal Protein
3.2. Complementary Extraction Processes
4. Perspectives
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, J.; Liu, S.; Huang, J.; Cui, R.; Xu, Y.; Song, Z. Genetic engineering strategies for sustainable polyhydroxyalkanoate (PHA) production from carbon-rich wastes. Environ. Technol. Innov. 2023, 30, 103069. [Google Scholar] [CrossRef]
- Atanasova, N.; Stoitsova, S.; Paunova-Krasteva, T.; Kambourova, M. Plastic degradation by extremophilic bacteria. Int. J. Mol. Sci. 2021, 22, 5610. [Google Scholar] [CrossRef]
- Moses, N.E.; Erhianoh, C.; Anih, C.E. Modelling and simulation of waste plastic power plant: A theoretical framework. Am. J. Chem. Eng. 2018, 6, 94–98. [Google Scholar] [CrossRef]
- Mahmoud, Y.S.; Belhanche-Bensemra, N.; Safidine, Z. Impact of microcrystalline cellulose extracted from walnut and apricots shells on the biodegradability of poly(lactic acid). Front. Mater. 2022, 9, 1005387. [Google Scholar] [CrossRef]
- Lebreton, L.; Andrady, A. Future scenarios of global plastic waste generation and disposal. Palgrave Commun. 2019, 5, 6. [Google Scholar] [CrossRef]
- Plastic Waste Management | Plastics and the Environment Series. Available online: https://www.genevaenvironmentnetwork.org/resources/updates/plastic-waste-management/ (accessed on 27 December 2023).
- Lonca, G.; Lesage, P.; Majeau-Bettez, G.; Bernard, S.; Margni, M. Assessing scaling effects of circular economy strategies: A case study on plastic bottle closed-loop recycling in the USA PET market. Resour. Conserv. Recycl. 2020, 162, 105013. [Google Scholar] [CrossRef]
- Kim, M.O.; Park, J.K.; Han, T.H.; Seo, J.; Lee, H. Influence of polyethylene terephthalate powder on hydration of portland cement. Polymers 2012, 13, 2551. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.; Encarnação, T.; Tavares, R.; Todo Bom, T.; Mateus, A. Bioplastics: Innovation for green transition. Polymers 2023, 15, 517. [Google Scholar] [CrossRef] [PubMed]
- Haider, T.P.; Völker, C.; Kramm, J.; Landfester, K.; Wurm, F.R. Plastics of the future? The impact of biodegradable polymers on the environment and on society. Angew. Chem. Int. Edit. 2019, 58, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Lin, Y.; Wu, Q.; Wang, Y.; Chen, G.Q. Synthetic biology and genome-editing tools for improving PHA metabolic engineering. Trends Biotechnol. 2020, 38, 7. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Chen, J.-C.; Chen, G.-Q. Controlling microbial PHB synthesis via CRISPRi. Appl. Microbiol. Biot. 2017, 101, 5861–5867. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.Q.; Jiang, X.R. Next generation industry biotechnology based on extremophiles. Curr. Opin. Biotechnol. 2018, 50, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, P.; Wang, H.; Hajnal, I.; Wu, Q.; Guo, Y.; Chen, G.Q. Increasing oxygen availability for improving poly(3-hydroxybutyrate) production by Halomonas. Metab. Eng. 2018, 45, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Landick, R.; Raman, S. A regulatory NADH/NAD+ redox biosensor for bacteria. ACS Synth. Biol. 2019, 8, 264–273. [Google Scholar] [CrossRef] [PubMed]
- Shen, R.; Ning, Z.Y.; Lan, Y.X.; Chen, J.C.; Chen, G.Q. Manipulation of polyhydroxyalkanoate granular sizes in Halomonas bluephagenesis. Metab. Eng. 2019, 54, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Anjum, A.; Zuber, M.; Zia, K.M.; Noreen, A.; Anjum, M.N.; Tabasum, S. Microbial production of polyhydroxyalkanoates (PHAs) and its copolymers: A review of recent developments. Int. J. Biol. Macromol. 2016, 89, 161–174. [Google Scholar] [CrossRef]
- Costa, S.S.; Miranda, A.L.; de Morais, M.G.; Costa, J.A.V.; Druzian, J.I. Microalgae as source of polyhydroxyalkanoates (PHAs) —A review. Int. J. Biol. Macromol. 2019, 131, 536–547. [Google Scholar] [CrossRef]
- Singh, M.; Kumar, P.; Patel, S.K.S.; Kalia, V.C. Production of polyhydroxyalkanoate co-polymer by Bacillus thuringiensis. Indian J. Microbiol. 2013, 53, 77–83. [Google Scholar] [CrossRef]
- Singh, M.; Patel, S.K.S.; Kalia, V.C. Bacillus subtilis as potential producer for polyhydroxyalkanoates. Microb. Cell Fact. 2009, 8, 38. [Google Scholar] [CrossRef] [PubMed]
- Balaji, S.; Gopi, K.; Muthuvelan, B. A review on production of poly β hydroxybutyrates from cyanobacteria for the production of bioplastics. Algal Res. 2013, 2, 278–285. [Google Scholar] [CrossRef]
- Singh, G.; Kumari, A.; Mittal, A.; Goel, V.; Yadav, A.; Aggarwal, N.K. Cost effective production of poly-β-hydroxybutyrate by Bacillus subtilis NG05 using sugar industry waste water. J. Polym. Environ. 2013, 21, 441–449. [Google Scholar] [CrossRef]
- Singh, A.K.; Mallick, N. Advances in cyanobacterial polyhydroxyalkanoates production. FEMS Microbiol. Lett. 2017, 364, fnx189. [Google Scholar] [CrossRef]
- Singh, M.; Kumar, P.; Ray, S.; Kalia, V.C. Challenges and opportunities for customizing polyhydroxyalkanoates. Indian J. Microbiol. 2015, 55, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Peelman, N.; Ragaert, R.; De Meulenaer, B.; Adons, D.; Peeters, R.; Cardon, L.; Van Impe, F.; Devlieghere, F. Application of bioplastics for food packaging. Trends Fd. Sci. Technol. 2013, 32, 128–141. [Google Scholar] [CrossRef]
- Ray, S.; Kalia, V.C. Biomedical applications of polyhydroxyalkanoates. Indian J. Microbiol. 2017, 57, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Kalia, V.C.; Ray, S.; Patel, S.K.; Singh, M.; Singh, G.P. The dawn of novel biotechnological applications of polyhydroxyalkanoates. In Biotechnological Applications of Polyhydroxyalkanoates; Springer: Singapore, 2019; pp. 1–11. [Google Scholar] [CrossRef]
- Venkatachalam, H.; Palaniswamy, R. Bioplastic World: A Review. J. Adv. Sci. Res. 2020, 11, 43–53. [Google Scholar]
- Pretula, J.; Slomkowski, S.; Penczek, S. Polylactides-methods of synthesis and characterization. Adv. Drug Deliv. Rev. 2016, 107, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.A.; Waymouth, R.M. Ion pairing effects in the zwitterionic ring opening polymerization of δ-valerolactone. Polym. Chem. 2015, 6, 5212–5218. [Google Scholar] [CrossRef]
- Inkinen, S.; Hakkarainen, M.; Albertsson, A.C.; Södergård, A. From lactic acid to poly(lactic acid) (PLA): Characterization and analysis of PLA and Its precursors. Biomacromolecules 2011, 12, 523–532. [Google Scholar] [CrossRef]
- Vert, M. Degradable and bioresorbable polymers in surgery and in pharmacology: Beliefs and facts. J. Mater. Sci. Mater. Med. 2009, 20, 437–446. [Google Scholar] [CrossRef]
- Woodruff, M.A.; Hutmacher, D.W. The return of a forgotten polymer—Polycaprolactone in the 21st century. Prog. Polym. Sci. 2010, 35, 1217–1256. [Google Scholar] [CrossRef]
- Isroi, I.; Cifriadi, A.; Panji, T.; Wibowo, N.A.; Syamsu, K. Bioplastic production from cellulose of oil palm empty fruit bunch. IOP Conf. Ser. Earth Environ. Sci. 2017, 65, 012011. [Google Scholar] [CrossRef]
- El-malek, F.A.; Khairy, H.; Farag, A.; Omar, S. The sustainability of microbial bioplastics, production and applications. Int. J. Biol. Macromol. 2020, 157, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Luengo, J.M.; García, B.; Sandoval, A.; Naharro, G.; Olivera, E.R. Bioplastics from microorganisms. Curr. Opin. Microbiol. 2003, 6, 251–260. [Google Scholar] [CrossRef]
- Karpušenkaite, A.; Varžinskas, V. Bioplastics: Development, possibilities and difficulties. Environ. Res. Eng. Manag. 2014, 68, 69–78. [Google Scholar] [CrossRef]
- Lackner, M. Bioplastics. In Kirk-Othmer Encyclopedia of Chemical Technology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 1–41. [Google Scholar] [CrossRef]
- Atiwesh, G.; Mikhael, A.; Parrish, C.C.; Banoub, J.; Le, T.-A.T. Environmental impact of bioplastic use: A review. Heliyon. 2021, 7, e07918. [Google Scholar] [CrossRef]
- Nandakumar, A.; Chuah, J.A.; Sudesh, K. Bioplastics: A boon or bane? Renew. Sustain. Energy Rev. 2021, 147, 111237. [Google Scholar] [CrossRef]
- Nanda, S.; Patra, B.R.; Patel, R.; Bakos, J.; Dalai, A.K. Innovations in applications and prospects of bioplastics and biopolymers: A review. Environ. Chem. Lett. 2022, 20, 379–395. [Google Scholar] [CrossRef]
- Jerez, A.; Partal, P.; Martínez, I.; Gallegos, C.; Guerrero, A. Protein-based bioplastics: Effect of thermo-mechanical processing. Rheol. Acta. 2007, 46, 711–720. [Google Scholar] [CrossRef]
- Costa, A.A.; Gameiro, F.; Potêncio, A.; Silva, D.P.; Carreira, P.; Martinez, J.C.; Pascoal-Faria, P.; Mateus, A.; Mitchell, G.R. Evaluating the injection moulding of plastic parts using in situ Time-Resolved Small-Angle X-ray Scattering Techniques. Polymers 2022, 14, 4745. [Google Scholar] [CrossRef]
- Eisoldt, L.; Smith, A.; Scheibel, T. Decoding the secrets of spider silk. Materialstoday 2011, 14, 80–86. [Google Scholar] [CrossRef]
- Qiao, X.; Qian, Z.; Li, J.; Sun, H.; Han, Y.; Xia, X.; Zhou, J.; Wang, C.; Wang, Y.; Wang, C. Synthetic engineering of spider silk fiber as implantable optical waveguides for low-loss light guiding. ACS Appl. Mater. Interfaces 2017, 9, 14665–14676. [Google Scholar] [CrossRef] [PubMed]
- Johansson, J.; Rising, A. Doing what spiders cannot-a road map to supreme artificial silk fibers. ACS Nano 2021, 15, 1952–1959. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.Q.; Patel, M.K. Plastics derived from biological sources: Present and future: A technical and environmental review. Chem. Rev. 2012, 112, 2082–2099. [Google Scholar] [CrossRef]
- Laycock, B.; Halley, P.; Pratt, S.; Werker, A.; Lant, P. The chemomechanical properties of microbial polyhydroxyalkanoates. Prog. Polym. Sci. 2013, 38, 536–583. [Google Scholar] [CrossRef]
- Koutinas, A.A.; Vlysidis, A.; Pleissner, D.; Kopsahelis, N.; Lopez Garcia, I.; Kookos, I.K.; Papanikolaou, S.; Kwan, T.H.; Lin, C.S. Valorization of industrial waste and by-product streams via fermentation for the production of chemicals and biopolymers. Chem. Soc. Rev. 2014, 43, 2587–2627. [Google Scholar] [CrossRef]
- Chen, G.-Q.; Chen, X.-Y.; Wu, F.-Q.; Chen, J.-C. Polyhydroxyalkanoates (PHA) toward cost competitiveness and functionality. Adv. Ind. Eng. Polymer Res. 2020, 3, 1–7. [Google Scholar] [CrossRef]
- Ademakinwa, A.N.; Ayinla, Z.A.; Agboola, F.K. Strain improvement and statistical optimization as a combined strategy for improving fructosyltransferase production by Aureobasidium pullulans NAC8. J. Genet. Eng. Biotechnol. 2017, 15, 345–358. [Google Scholar] [CrossRef]
- Cheng, K.K.; Zhao, X.B.; Zeng, J.; Wu, R.C.; Xu, Y.Z.; Liu, D.H.; Zhang, J.A. Downstream processing of biotechnological produced succinic acid. Appl. Microbiol. Biotechnol. 2012, 95, 841–850. [Google Scholar] [CrossRef]
- Jungbauer, A. Continuous downstream processing of biopharmaceuticals. Trends Biotechnol. 2013, 31, 479–492. [Google Scholar] [CrossRef]
- Kreyenschulte, D.; Krull, R.; Margaritis, A. Recent advances in microbial biopolymer production and purification. Crit. Rev. Biotechnol. 2014, 34, 1–15. [Google Scholar] [CrossRef]
- Jiang, X.R.; Chen, G.Q. Morphology engineering of bacteria for bio-production. Biotechnol. Adv. 2016, 34, 435–440. [Google Scholar] [CrossRef] [PubMed]
- van Teeffelen, S.; Gitai, Z. Rotate into shape: MreB and bacterial morphogenesis. EMBO J. 2011, 30, 4856–4857. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.; Wu, Q.; Chen, J.C.; Chen, G.Q. Engineering Halomonas TD01 for the lowcost production of polyhydroxyalkanoates. Metab. Eng. 2014, 26, 34–47. [Google Scholar] [CrossRef] [PubMed]
- Bisson-Filho, A.W.; Discola, K.F.; Castellen, P.; Blasios, V.; Martins, A.; Sforca, M.L.; Garcia, W.; Zeri, A.C.; Erickson, H.P.; Dessen, A.; et al. FtsZ filament capping by MciZ, a developmental regulator of bacterial division. Proc. Natl. Acad. Sci. USA 2015, 112, E2130–E2138. [Google Scholar] [CrossRef]
- Jiang, X.R.; Wang, H.; Shen, R.; Chen, G.Q. Engineering the bacterial shapes for enhanced inclusion bodies accumulation. Metab. Eng. 2015, 29, 227–237. [Google Scholar] [CrossRef]
- Bernhardt, T.G.; de Boer, P.A.J. SlmA, a Nucleoid-associated, FtsZ binding protein required for blocking septal ring assembly over chromosomes in E. coli. Mol. Cell. 2005, 18, 555–564. [Google Scholar] [CrossRef]
- Van Heijenoort, J. Peptidoglycan hydrolases of Escherichia coli. Microbiol. Mol. Biol. Rev. 2011, 75, 636–663. [Google Scholar] [CrossRef]
- Shiomi, D.; Niki, H. A mutation in the promoter region of zipA, a component of the divisome, suppresses the shape defect of RodZ-deficient cells. MicrobiologyOpen 2013, 2, 798–810. [Google Scholar] [CrossRef]
- Shiomi, D.; Toyoda, A.; Aizu, T.; Ejima, F.; Fujiyama, A.; Shini, T.; Kohara, Y.; Niki, H. Mutations in cell elongation genes mreB, mrdA and mrdB suppress the shape defect of RodZ-deficient cells. Mol. Microbiol. 2013, 87, 1029–1044. [Google Scholar] [CrossRef]
- Loose, M.; Mitchison, T.J. The bacterial cell division proteins FtsA and FtsZ self-organize into dynamic cytoskeletal patterns. Nat. Cell Biol. 2014, 16, 38–46. [Google Scholar] [CrossRef]
- Zhang, X.C.; Guo, Y.; Liu, X.; Chen, X.G.; Wu, Q.; Chen, G.Q. Engineering cell wall synthesis mechanism for enhanced PHB accumulation in E. coli. Metab. Eng. 2018, 45, 32–42. [Google Scholar] [CrossRef]
- Turner, R.D.; Hurd, A.F.; Cadby, A.; Hobbs, J.K.; Foster, S.J. Cell wall elongation mode in Gram-negative bacteria is determined by peptidoglycan architecture. Nat. Commun. 2013, 4, 1496. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, J.M.; Pereira, A.R.; Reichmann, N.T.; Saraiva, B.M.; Fernandes, P.B.; Veiga, H.; Tavares, A.C.; Santos, M.; Ferreira, M.T.; Macario, V.; et al. Peptidoglycan synthesis drives an FtsZ-treadmilling-independent step of cytokinesis. Nature 2018, 554, 528–532. [Google Scholar] [CrossRef] [PubMed]
- van Teeffelen, S.; Renner, L.D. Recent advances in understanding how rod-like bacteria stably maintain their cell shapes. F1000Research 2018, 7, 241. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ling, C.; Chen, Y.; Jiang, X.; Chen, G.-Q. Microbial engineering for easy downstream processing. Biotechnol. Adv. 2019, 37, 107365. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.K.; Parveen, S.; SaiSree, L.; Reddy, M. Regulated proteolysis of a crosslink-specific peptidoglycan hydrolase contributes to bacterial morphogenesis. Proc. Natl. Acad. Sci. USA 2015, 112, 10956–10961. [Google Scholar] [CrossRef]
- Egan, A.J.; Vollmer, W. The physiology of bacterial cell division. Ann. N. Y. Acad. Sci. 2013, 1277, 8–28. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.R.; Yao, Z.H.; Chen, G.Q. Controlling cell volume for efficient PHB production by Halomonas. Metab. Eng. 2017, 44, 30–37. [Google Scholar] [CrossRef]
- Elhadi, D.; Lv, L.; Jiang, X.R.; Wu, H.; Chen, G.Q. CRISPRi engineering E. coli for morphology diversification. Metab. Eng. 2016, 38, 358e369. [Google Scholar] [CrossRef]
- Wu, H.; Chen, J.; Chen, G.Q. Engineering the growth pattern and cell morphology for enhanced PHB production by Escherichia coli. Appl. Microbiol. Biotechnol. 2016, 100, 9907–9916. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, H.; Jiang, X.; Chen, G.Q. Engineering Escherichia coli for enhanced production of poly(3-hydroxybutyrate-co-4-hydroxybutyrate) in larger cellular space. Metab. Eng. 2014, 25, 183–193. [Google Scholar] [CrossRef]
- Ghosal, D.; Trambaiolo, D.; Amos, L.A.; Lowe, J. MinCD cell division proteins form alternating copolymeric cytomotive filaments. Nat. Commun. 2014, 5, 5341. [Google Scholar] [CrossRef] [PubMed]
- Uehara, T.; Dinh, T.; Bernhardt, T.G. LytM-domain factors are required for daughter cell separation and rapid ampicillin-induced lysis in Escherichia coli. J. Bacteriol. 2009, 191, 5094–5107. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Gong, T.; Liu, X.; Fan, X.; Huang, R.; Ma, T.; Wang, S.; Gao, W.; Yang, C. Morphology engineering for enhanced production of medium-chain-length polyhydroxyalkanoates in Pseudomonas mendocina NK-01. Appl. Microbiol. Biotechnol. 2019, 103, 1713–1724. [Google Scholar] [CrossRef] [PubMed]
- Strahl, H.; Burmann, F.; Hamoen, L.W. The actin homologue MreB organizes the bacterial cell membrane. Nat. Commun. 2014, 5, 3442. [Google Scholar] [CrossRef] [PubMed]
- Morgenstein, R.M.; Bratton, B.P.; Nguyen, J.P.; Ouzounov, N.; Shaevitz, J.W.; Gitai, Z. RodZ links MreB to cell wall synthesis to mediate MreB rotation and robust morphogenesis. Proc. Natl. Acad. Sci. USA 2015, 112, 12510–12515. [Google Scholar] [CrossRef] [PubMed]
- Colavin, A.; Shi, H.; Huang, K.C. RodZ modulates geometric localization of the bacterial actin MreB to regulate cell shape. Nat. Commun. 2018, 9, 1280. [Google Scholar] [CrossRef]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A.; et al. Multiplex genome engineering using CRISPR/Cas systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [PubMed]
- Cress, B.F.; Jones, J.A. Rapid generation of CRISPR/dCas9-regulated, orthogonally repressible hybrid T7-lac promoters for modular, tuneable control of metabolic pathway fluxes in Escherichia coli. Nucleic Acids Res. 2016, 44, 4472–4485. [Google Scholar] [CrossRef]
- Peternel, S. Bacterial cell disruption: A crucial step in protein production. New Biotechnol. 2013, 30, 250–254. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Feng, X.; Xian, M.; Wang, Q.; Zhao, G. Inducible cell lysis systems in microbial production of bio-based chemicals. Appl. Microbiol. Biotechnol. 2013, 97, 7121–7129. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.I.; Lee, S.Y. Efficient and economical recovery of poly(3-hy-droxybutyrate) from recombinant Escherichia coli by simple digestion with chemicals. Biotechnol. Bioeng. 1999, 62, 546–553. [Google Scholar] [CrossRef]
- Zhang, X.; Pan, Z.; Fang, Q.; Zheng, J.; Hu, M.; Jiao, X. An auto-inducible Escherichia coli lysis system controlled by magnesium. J. Microbiol. Methods 2009, 79, 199–204. [Google Scholar] [CrossRef]
- Hajnal, I.; Chen, X.; Chen, G.Q. A novel cell autolysis system for cost-competitive downstream processing. Appl. Microbiol. Biotechnol. 2016, 100, 9103–9110. [Google Scholar] [CrossRef]
- Martínez, V.; García, P.; García, J.L.; Prieto, M.A. Controlled autolysis facilitates the polyhydroxyalkanoate recovery in Pseudomonas putida KT2440. Microb. Biotechnol. 2011, 4, 533–547. [Google Scholar] [CrossRef]
- Borrero-De Acuna, J.M.; Hidalgo-Dumont, C.; Pacheco, N.; Cabrera, A.; Poblete-Castro, I. A novel programmable lysozyme-based lysis system in Pseudomonas putida for biopolymer production. Sci. Rep. 2017, 7, 4373. [Google Scholar] [CrossRef]
- Poblete-Castro, I.; Aravena-Carrasco, C.; Orellana-Saez, M.; Pacheco, N.; Cabrera, A.; Borrero-de Acuña, J.M. Engineering the osmotic state of Pseudomonas putida KT2440 for efficient cell disruption and downstream processing of poly(3-Hydroxyalkanoates). Front Bioeng Biotechnol. 2020, 8, 161. [Google Scholar] [CrossRef]
- Morita, M.; Tanji, Y.; Mizoguchi, K.; Soejima, A.; Orito, Y.; Unno, H. Antibacterial activity of Bacillus amyloliquefaciens phage endolysin without holing conjugation. J. Biosci. Bioeng. 2001, 91, 469–473. [Google Scholar] [CrossRef]
- Hori, K.; Kaneko, M.; Tanji, Y.; Xing, X.H.; Unno, H. Construction of self disruptive Bacillus megaterium in response to substrate exhaustion for polyhydroxybutyrate production. Appl. Microbiol. Biotechnol. 2002, 59, 211–216. [Google Scholar] [CrossRef]
- Salzberg, L.I.; Helmann, J.D. An antibiotic inducible cell wall associated protein that protects Bacillus subtilis from autolysis. J. Bacteriol. 2007, 189, 4671–4680. [Google Scholar] [CrossRef]
- Chatsungnoen, T.; Chisti, Y. Continuous flocculation-sedimentation for harvesting Nannochloropsis salina biomass. J. Biotechnol. 2016, 222, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.G.; Hao, X.M.; Lin, Y.H.; Bai, F.W. Redox potential driven aeration during very-high-gravity ethanol fermentation by using flocculating yeast. Sci. Rep. 2016, 6, 25763. [Google Scholar] [CrossRef]
- Cheng, C.; Zhang, M.M.; Xue, C.; Bai, F.W.; Zhao, X.Q. Development of stress tolerant Saccharomyces cerevisiae strains by metabolic engineering: New aspects from cell flocculation and zinc supplementation. J. Biosci. Bioeng. 2017, 123, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Ling, C.; Qiao, G.Q.; Shuai, B.W.; Song, K.N.; Yao, W.X.; Jiang, X.R.; Chen, G.Q. Engineering self-flocculating Halomonas campaniensis for wastewaterless open and continuous fermentation. Biotechnol. Bioeng. 2018, 116, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Koller, M. Established and advanced approaches for recovery of microbial polyhydroxyalkanoate (PHA) biopolyesters from surrounding microbial biomass. EuroBiotech. J. 2020, 4, 113–126. [Google Scholar] [CrossRef]
- Rebocho, A.T.; Pereira, J.R.; Neves, L.A.; Alves, V.D.; Sevrin, C.; Grandfils, C.; Freitas, F.; Reis, M.A.M. Preparation and characterization of films based on a natural P(3HB)/mcl-PHA blend obtained through the co-culture of Cupriavidus necator and Pseudomonas citronellolis in apple pulp waste. Bioengineering 2020, 7, 34. [Google Scholar] [CrossRef]
- Ojha, N.; Das, N. Process optimization and characterization of polyhydroxyalkanoate copolymers produced by marine Pichia kudriavzevii VIT-NN02 using banana peels and chicken feather hydrolysate. Biocat. Agri. Biotechnol. 2020, 27, 101616. [Google Scholar] [CrossRef]
- Koller, M.; Bona, R.; Chiellini, E.; Braunegg, G. Extraction of short-chain- length poly-(I-hydroxyalkanoates) (scl-PHA) by the “anti-solvent” acetone under elevated temperature and pressure. Biotechnol. Lett. 2013, 35, 1023–1028. [Google Scholar] [CrossRef]
- Cerrone, F.; Radivojevic, J.; Nikodinovic-Runic, J.; Walsh, M.; Kenny, S.T.; Babu, R.; O’Connor, K.E. Novel sodium alkyl-1, 3-disulfates, anionic biosurfactants produced from microbial polyesters. Colloid Surface B 2019, 182, 110333. [Google Scholar] [CrossRef]
- Tang, S.; Baker, G.A.; Zhao, H. Ether- and alcohol-functionalized task-specific ionic liquids: Attractive properties and applications. Chem. Soc. Rev. 2012, 41, 4030–4066. [Google Scholar] [CrossRef]
- Fujita, K.; Kobayashi, D.; Nakamura, N.; Ohno, H. Direct dissolution of wet and saliferous marine microalgae by polar ionic liquids without heating. Enzyme Microb. Tech. 2013, 52, 199–202. [Google Scholar] [CrossRef]
- Kobayashi, D.; Fujita, K.; Nakamura, N.; Ohno, H. A simple recovery process for biodegradable plastics accumulated in cyanobacteria treated with ionic liquids. Appl. Microbiol. Biotechnol. 2015, 99, 1647–1653. [Google Scholar] [CrossRef]
- Kalia, V.C.; Patel, S.K.S.; Lee, J.-K. Exploiting polyhydroxyalkanoates for biomedical applications. Polymers 2023, 15, 1937. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez Gamero, J.E.; Favaro, L.; Pizzocchero, V.; Lomolino, G.; Basaglia, M.; Casella, S. Nuclease expression in efficient polyhydroxyalkanoates-producing bacteria could yield cost reduction during downstream processing. Bioresour. Technol. 2018, 261, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Ray, S.; Kalia, V.C. Polyhydroxyalkanoate production and degradation patterns in Bacillus species. Indian J. Microbiol. 2017, 57, 387–392. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalia, V.C.; Patel, S.K.S.; Karthikeyan, K.K.; Jeya, M.; Kim, I.-W.; Lee, J.-K. Manipulating Microbial Cell Morphology for the Sustainable Production of Biopolymers. Polymers 2024, 16, 410. https://doi.org/10.3390/polym16030410
Kalia VC, Patel SKS, Karthikeyan KK, Jeya M, Kim I-W, Lee J-K. Manipulating Microbial Cell Morphology for the Sustainable Production of Biopolymers. Polymers. 2024; 16(3):410. https://doi.org/10.3390/polym16030410
Chicago/Turabian StyleKalia, Vipin C., Sanjay K. S. Patel, Kugalur K. Karthikeyan, Marimuthu Jeya, In-Won Kim, and Jung-Kul Lee. 2024. "Manipulating Microbial Cell Morphology for the Sustainable Production of Biopolymers" Polymers 16, no. 3: 410. https://doi.org/10.3390/polym16030410