
Comparative Studies on the Structure and Biological Activities of Two New Polysaccharides from Tricholoma sinoportentosum (TS-P) and Termitomyces albuminosus (TA-P)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Extraction, Separation and Purification of Polysaccharide
2.2. Polysaccharide Molecular Weight Analysis
2.3. Fourier Transform Infrared (FT-IR) Spectroscopy Analysis
2.4. Methylation Analysis, GC-MS and Nuclear Magnetic Resonance (NMR) Assay
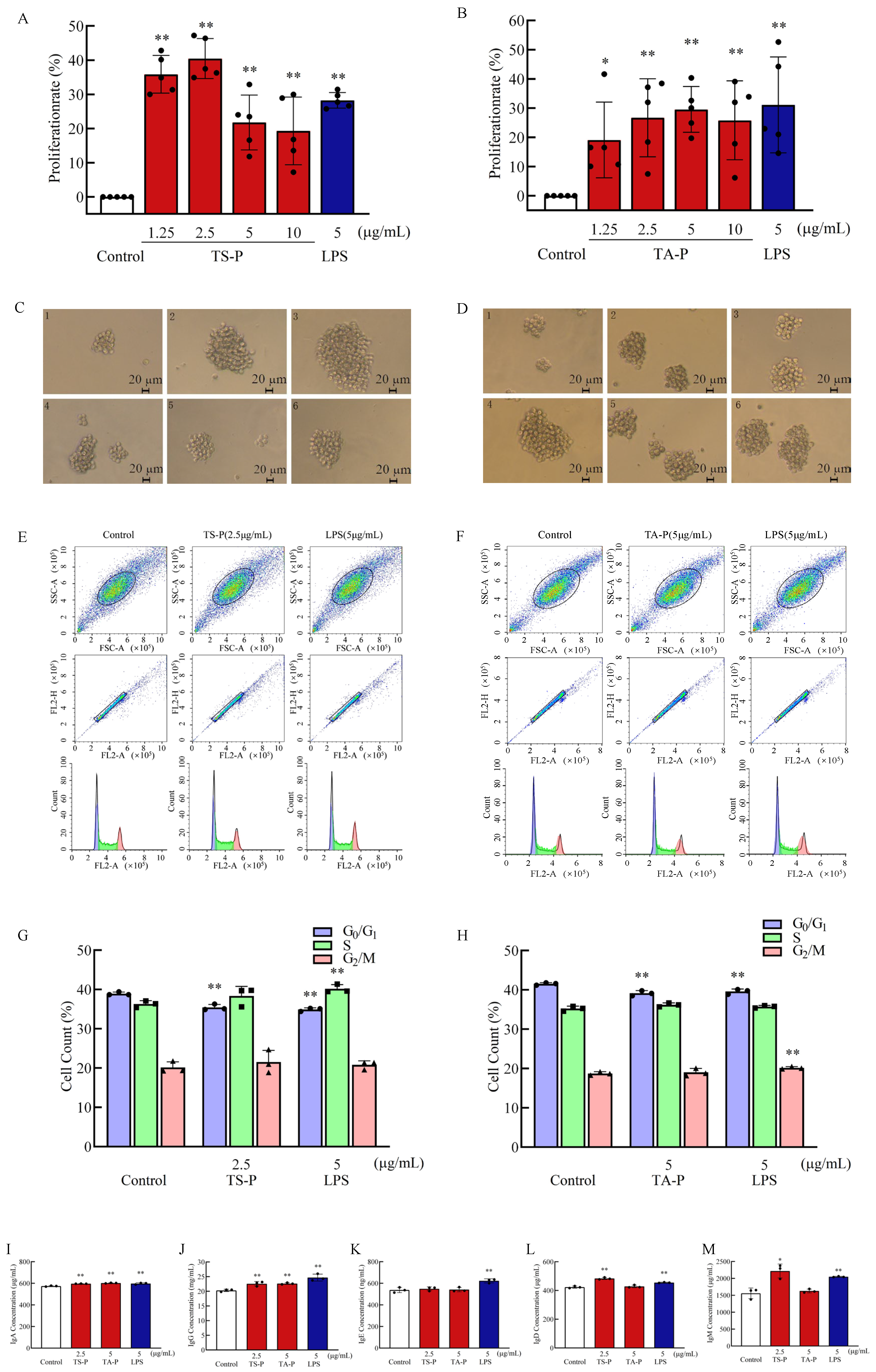
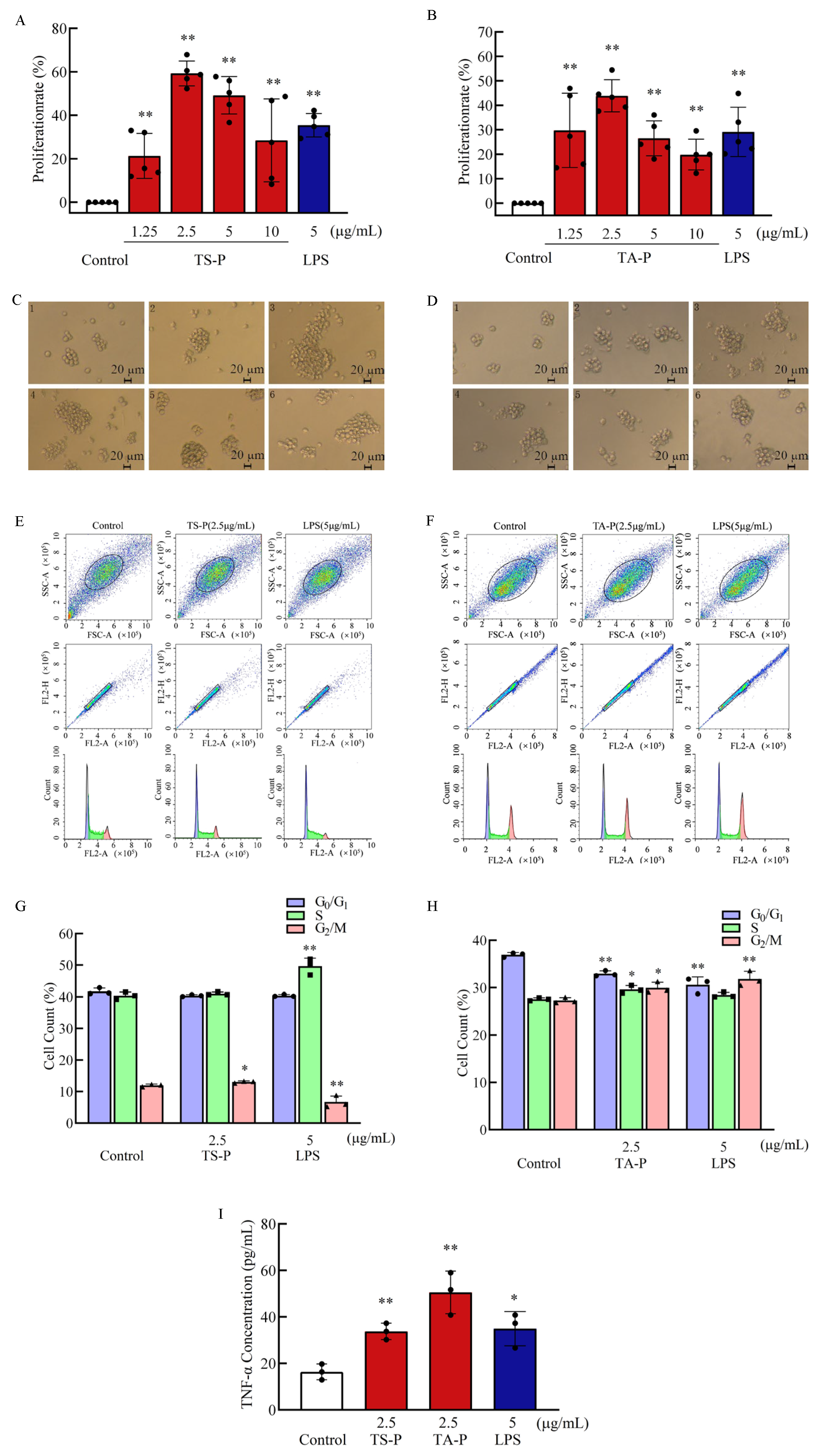
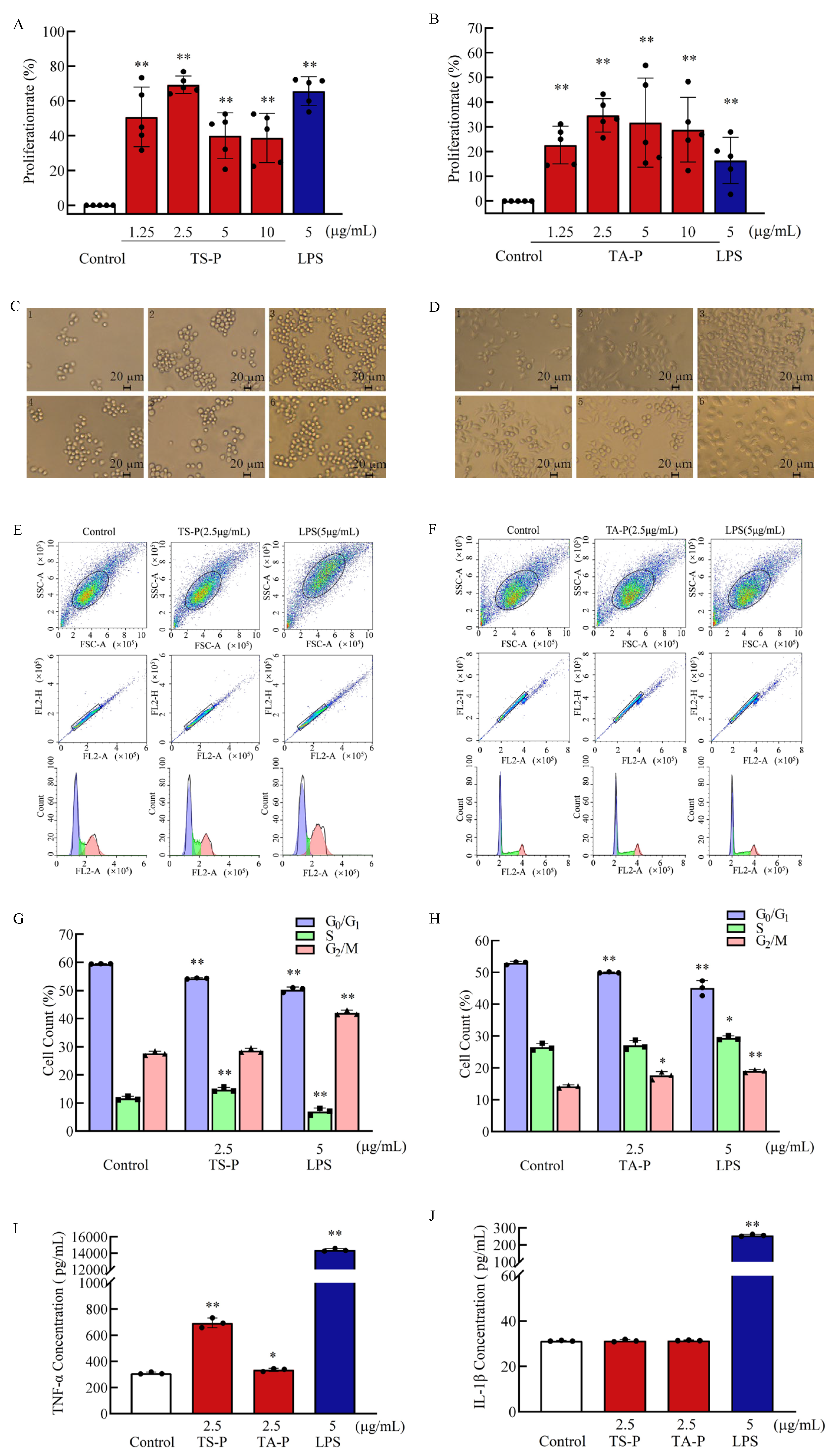
2.5. Effects of Polysaccharide on T Cell, B Cell and RAW264.7 Cell Proliferation and Cell Cycle
2.6. Effects of Polysaccharide on the Release of Cytokines in T Cells, B Cells and RAW264.7 Cells
2.7. Statistical Analysis
3. Results and Discussion
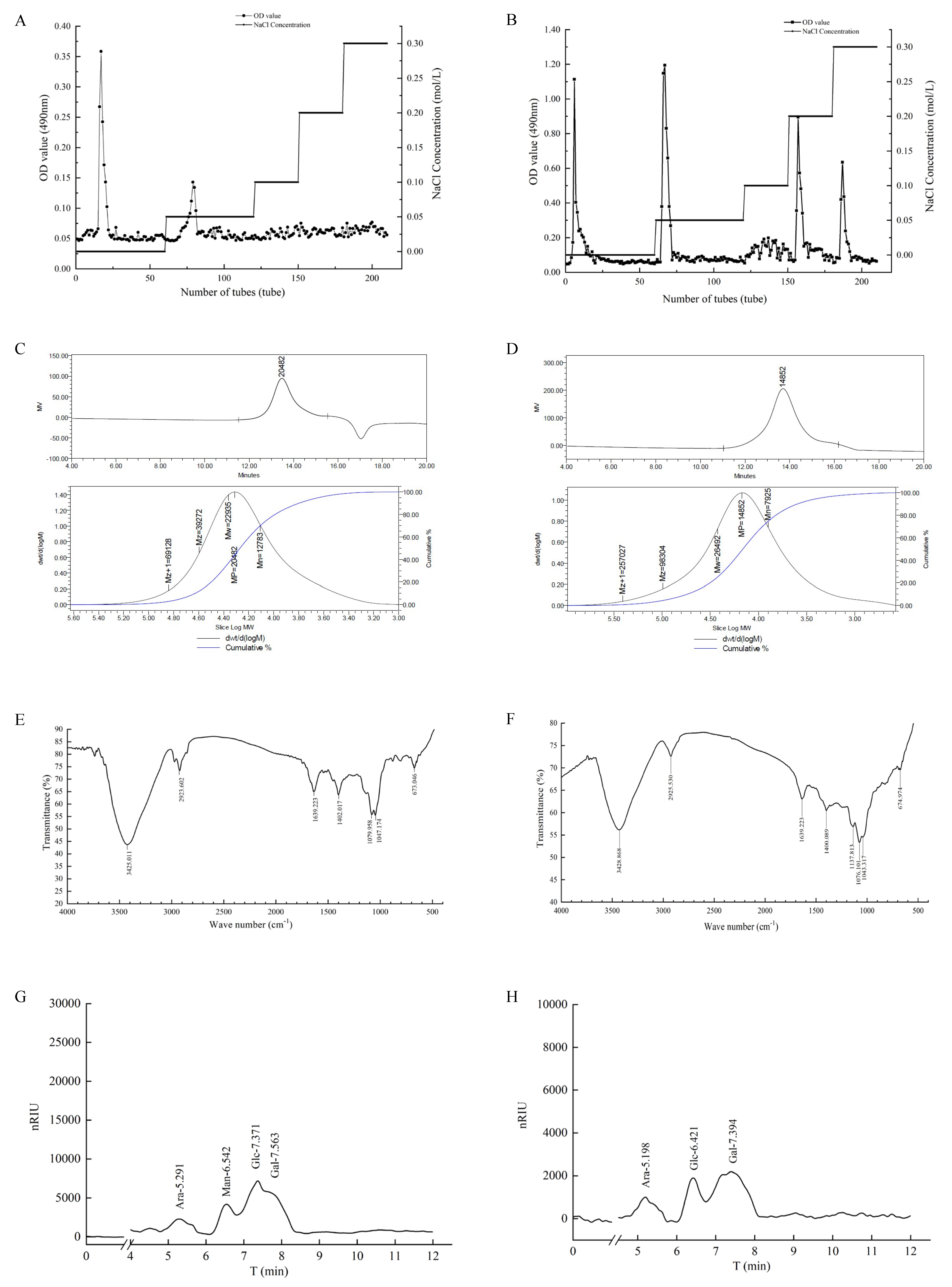
3.1. Elution Curve of TS-P and TA-P
3.2. Molecular Weight of TS-P and TA-P
3.3. FT-IR Analysis of TS-P and TA-P
3.4. Monosaccharide Composition Analysis of TS-P and TA-P
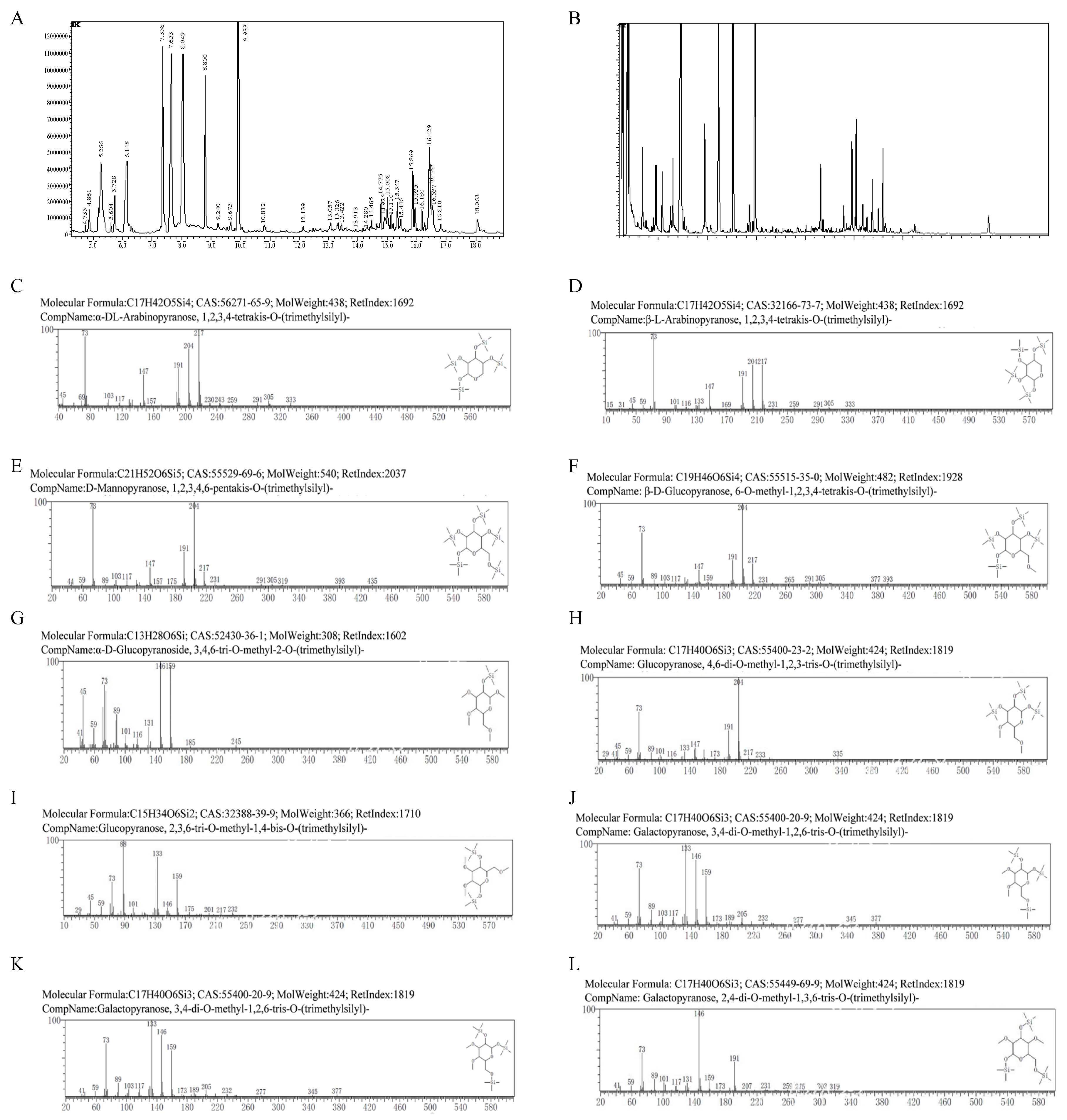
3.5. GC-MS Analysis of TS-P and TA-P
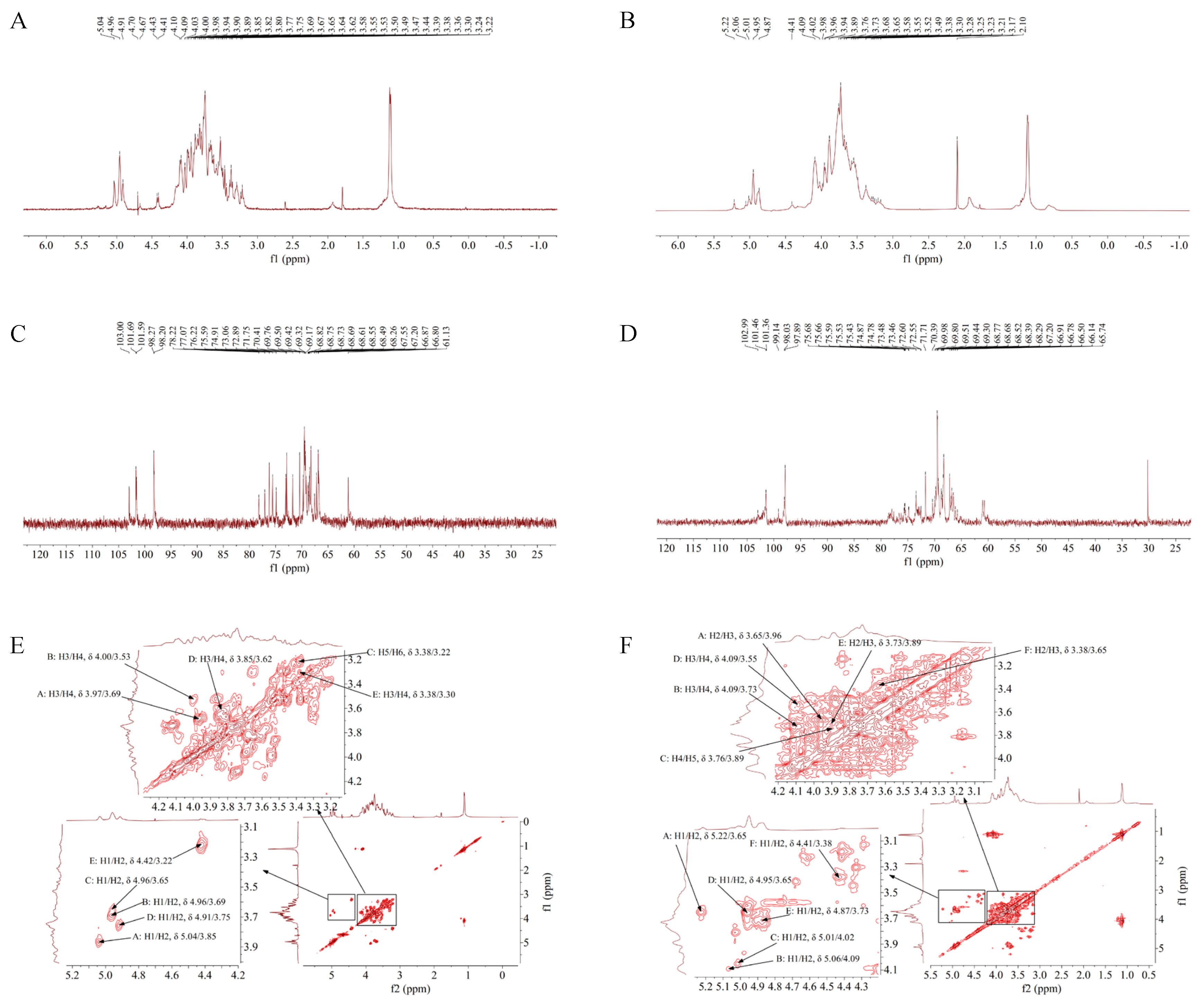
3.6. 1H-NMR Analysis of TS-P and TA-P
3.7. 13C-NMR Analysis of TS-P and TA-P
3.8. 1H-1H COSY Analysis of TS-P and TA-P
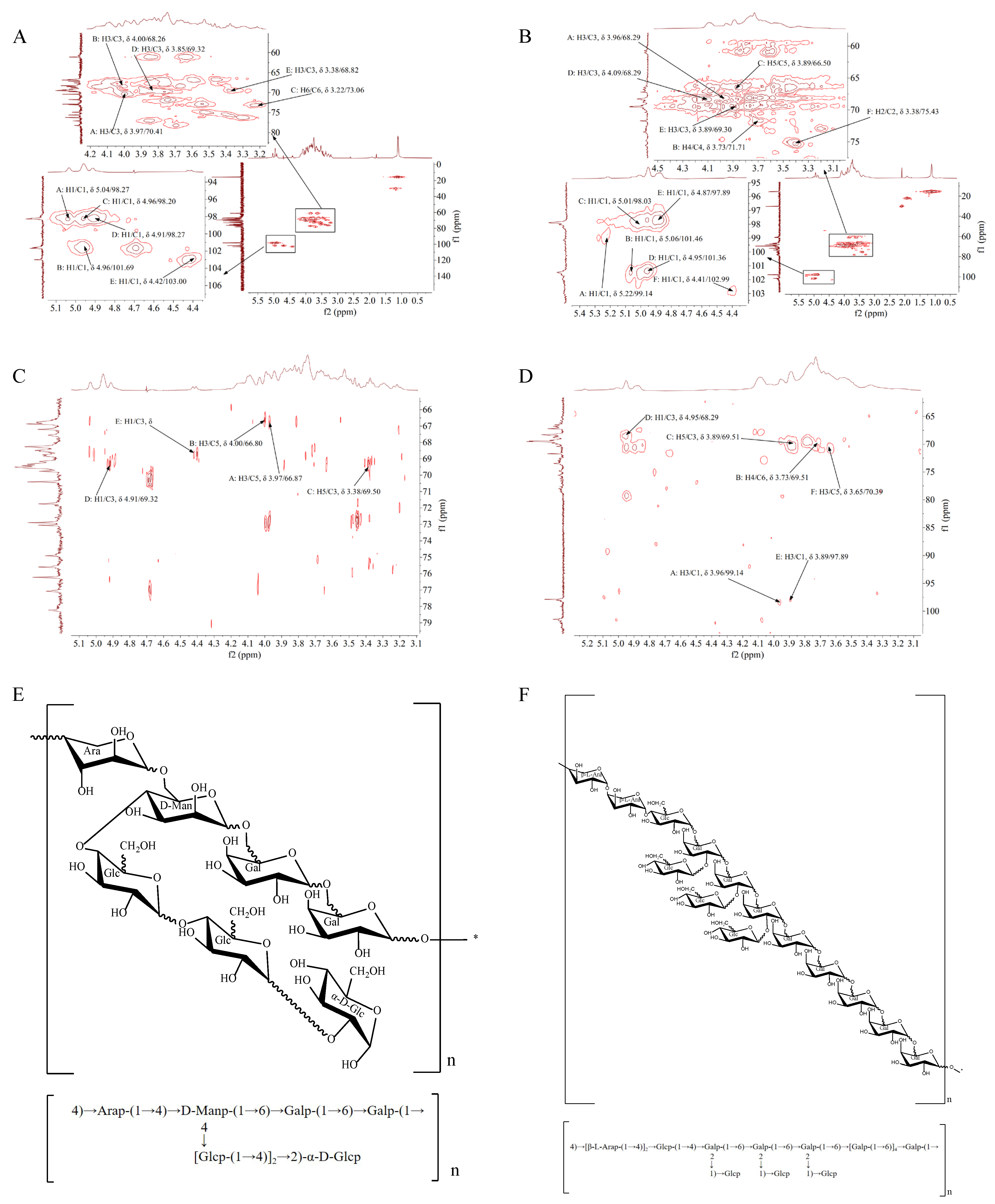
3.9. HMQC Analysis of TS-P and TA-P
3.10. HMBC Analysis of TS-P and TA-P
3.11. Effect of TS-P and TA-P on B Cell Activity In Vitro
3.12. Effect of TS-P and TA-P on T Cells Activity In Vitro
3.13. Effect of TS-P and TA-P on RAW264.7 Cells Activity In Vitro
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Izquierdo, M.A.; Scheffer, G.L.; Flens, M.J.; Giaccone, G.; Scheper, R.J. Broad distribution of the multidrug resistance-related vault lung resistance protein in normal human tissues and tumors. Am. J. Pathol. 1996, 148, 877–887. [Google Scholar]
- Dez, V.A.; Alvarez, A. Compositional and nutritional studies on two wild edible mushrooms from north west Spain. Food Chem. 2001, 75, 417–422. [Google Scholar] [CrossRef]
- Dogasaki, C.; Seki, W.; Kakuno, Y.; Kawakami, Y. Bioactivities and chemical properties of components obtained from coprinus comatus of an edible mushroom. J. Azabu. Univ. 2006, 13, 236–240. [Google Scholar]
- Mukhopadhyay, M. Bioactivities of some medicinal mushrooms: A modern perspective. J. Mycopath. Res. 2008, 1, 13–21. [Google Scholar]
- Polizeli, M.; Rizzatti, A.; Monti, R.; Terenzi, H.F.; Jorge, J.A.; Amorim, D.S. Xylanases from fungi: Properties and industrial applications. Appl. Microbio. Biotech. 2005, 67, 577–591. [Google Scholar] [CrossRef]
- Chihara, G. Inhibition of mouse srcoma 180 by polysaccharides from lentinus edodes (berk.) Sing. Nature 1969, 222, 687–688. [Google Scholar] [CrossRef]
- Yoshida, O.; Nakashima, H.; Yoshida, T.; Kaneko, Y.; Yamamoto, I.; Matsuzaki, K.; Uryu, T.; Yamamoto, N. Sulfation of the immunomodulating polysaccharide lentinan: A novel strategy for antivirals to human immunodeficiency virus (HIV). Biochem. Pharmacol. 1988, 37, 2887–2891. [Google Scholar] [CrossRef]
- Djordjevic, B.; Škugor, S.; Jørgensen, S.M.; Øverland, M.; Mydland, L.T.; Krasnov, A. Modulation of splenic immune responses to bacterial lipopolysaccharide in rainbow trout (Oncorhynchus mykiss) fed lentinan, a beta-glucan from mushroom Lentinula edodes. Fish Shellfish. Immunol. 2009, 26, 201–209. [Google Scholar] [CrossRef]
- Grenha, A. Editorial: Exploring the Role of Polysaccharides in Drug Delivery. Curr. Pharm. Des. 2015, 21, 4773–4774. [Google Scholar] [CrossRef]
- Brdaric, E.; Sokovic, S.; Djokic, J.; Nikolic, M. Protective effect of an exopolysaccharide produced by lactiplantibacillus plantarum bgan8 against cadmium-induced toxicity in caco-2 cells. Front. Microbio. 2021, 12, 759378. [Google Scholar] [CrossRef]
- Yao, X.Y.; Niu, W.R.; Cui, X.M.; Liu, Y.; Yang, X.Y.; Liu, J.K. Chemical Constituents from Cultures of the Tricholoma portrntosum. J. Chinese Med. Mater. 2017, 40, 1322–1325. [Google Scholar]
- Barros, L.; Baptista, P.; Correia, D.M.; Casal, S.; Oliveira, B.; Ferreira, I.C.F.R. Fatty acid and sugar compositions, and nutritional value of five wild edible mushrooms from northeast portugal. Food Chem. 2007, 105, 140–145. [Google Scholar] [CrossRef]
- Iwona, M.; Anna, W.M. Evaluation of Polish wild Mushrooms as Beta-Glucan Sources. Int. J. Environ. Res. Public Health 2020, 17, 7299. [Google Scholar]
- Shi, M.X.; Zhu, L.I.; Yang, K.Y.; Shao, J.M.; Ying-Xia, H.E. Analysis on the Nutritional Components in Different Parts of Termitomyces albuminosus. Food Res. Dev. 2012, 33, 182–183. [Google Scholar]
- Zhang, L.Z.; Chen, J. Analysis and determination of components in the fruit body of Collybia albuminosa. Sci. Tech. Food Indus. 2012, 33, 358–361. [Google Scholar]
- Zhao, L.F.; Zhang, J. Mycelium polysaccharides from Termitomyces albuminosus attenuate CCl4-Induced chronic liver injury via inhibiting TGFβ1/Smad3 and NF-κB signal pathways. Int. J. Mol. Sci. 2019, 20, 4872–4893. [Google Scholar] [CrossRef]
- Huajie, Z.; Juan, L.; Jianjun, Z.; Xiuxiu, W.; Min, L.; Chen, Z.; Le, J. Hepatoprotective and in vitro antioxidant effects of native depolymerised-exopolysaccharides derived from Termitomyces albuminosus. Sci. Rep. 2017, 7, 3910–3923. [Google Scholar]
- Jia, L.; Zhang, J.; Liu, L.; Zhao, Y.; Che, H. The antihyperlipidemic activities of enzymatic and acidic intracellular polysaccharides by Termitomyces albuminosus. Carbohy. Polym. Sci. Technol. Asp. Ind. Important Polysacch. 2016, 151, 1227–1234. [Google Scholar]
- Zhao, Y.X.; Tao, M.X.; Wen-Juan, L.U.; Cheng, G.Y.; Xing, J.I. Protective Effect of Polysaccharides from Termitomyces albuminosus on Alcoholic Liver Injury in Mice. Food Sci. 2014, 35, 260–265. [Google Scholar]
- Rong, H.E.; Luo, X.; Zhang, S.; Zhang, W. The Research Status of the Nutritional and Medicinal Value and Products of Termitomyces aurantiacus. Farm. Prod. Proces. 2019, 7, 74–77. [Google Scholar]
- Terbojevich, M.; Cosani, A.; Focher, B.; Marsano, E. High-performance gel-permeation chromatography of chitosan samples. Carbohydr. Res. 1993, 250, 301–314. [Google Scholar] [CrossRef]
- Chowdhury, S.R.; Basak, R.K.; Sen, R.; Adhikari, B. Production of extracellular polysaccharide by bacillus megaterium rb-05 using jute as substrate. Biores. Tech. 2011, 102, 6629–6632. [Google Scholar] [CrossRef]
- Scamparini, A.; Mariuzzo, D.; Fujihara, H.; Jacobusi, R.; Vendruscolo, C. Structural studies of cv-70 polysaccharide. Int. J. Biol. Macromol. 1997, 21, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Sudhamani, S.R.; Tharanathan, R.N.; Prasad, M.S. Isolation and characterization of an extracellular polysaccharide from pseudomonas caryophylli cfr 1705. Carbohydrate. Polym. 2004, 56, 423–427. [Google Scholar] [CrossRef]
- Pizzoferrato, L.; Manzi, P.; Bertocchi, F.; Fanelli, C.; Paci, M. Solid-state (13)c cp mas nmr spectroscopy of mushrooms gives directly the ratio between proteins and polysaccharides. J. Agri. Food Chem. 2000, 48, 5484–5488. [Google Scholar] [CrossRef]
- Brunger, A.T.; Adams, A.; Clore, B.; Delano, C.; Gros, D.P. Crystallography & nmr system: A new software suite for macromolecular structure determination. Acta. Crystallogr. 1998, 54 Pt 5, 905–921. [Google Scholar]
- Abeygunawardana, C.; Williams, T.C.; Sumner, J.S.; Hennessey, J.J. Development and validation of an NMR-based identity assay for bacterial polysaccharides. Analy. Biochem. 2000, 279, 226–240. [Google Scholar] [CrossRef]
- Katritzky, A.R.; Akhmedov, N.G.; Yang, H.; Hall, C.D. 1H and 13C NMR spectra of n-substituted morpholines. Mag. Reson. Chem. 2011, 43, 673–675. [Google Scholar] [CrossRef]
- Vuister, G.W.; Ruff-Cabello, J.; Zijl, P. Gradient-enhanced multiple-quantum filter (ge-mqf). a simple way to obtain single-scan phase-sensitive hmqc spectra. J. Mag. Reson. 1992, 100, 215–220. [Google Scholar] [CrossRef]
- Mélissa, D.; Canlet, C.; Vercauteren, J.; Andre, F.; Paris, A. Homeostatic signature of anabolic steroids in cattle using 1H-13C HMBC nmr metabonomics. J. Proteome. Res. 2005, 4, 1493–1502. [Google Scholar]
- Reddy, K.; Mohan, G.K.; Satla, S.; Gaikwad, S. Natural polysaccharides: Versatile excipients for controlled drug delivery systems. Asian J. Pharm. Sci. 2011, 6, 275–286. [Google Scholar]
- Xiao, J.B.; Jiang, H. A review on the structure-function relationship aspect of polysaccharides from tea materials. Crit. Rev. Food Sci. Nutr. 2015, 55, 930–938. [Google Scholar] [CrossRef]
- Supatra, K.; SangGuan, Y. Molecular characteristics of sulfated polysaccharides from Monostroma nitidum and their in vitro anticancer and immunomodulatory activities. Int. J. Biol. Macromol. 2011, 48, 311–318. [Google Scholar]
- Bernasconi, N.L.; Traggiai, E.; Lanzavecchia, A. Maintenance of serological memory by polyclonal activation of human memory b cells. Science 2002, 298, 2199–2202. [Google Scholar] [CrossRef]
- Elledge, S.J. Cell cycle checkpoints: Preventing an iden-tity crisis. Science 1997, 274, 1664–1672. [Google Scholar] [CrossRef]
- Raison, C.L.; Capuron, L.; Miller, A.H. Cytokines sing the blues: Inflammation and the pathogenesis of depression. Trends Immunol. 2006, 27, 24–31. [Google Scholar] [CrossRef]
- Aringer, M.; Smolen, J.S. Cytokine expression in lupus kidneys. Lupus 2005, 14, 13–18. [Google Scholar] [CrossRef]
- Park, H.; Li, Z.; Yang, X.O.; Chang, S.H.; Nurieva, R.; Wang, Y.H.; Wang, Y.; Hood, L.; Zhu, Z.; Tian, Q.; et al. A distinct lineage of cd4 t cells regulates tissue inflammation by producing interleukin 17. Nat. Immunol. 2005, 6, 1133–1141. [Google Scholar] [CrossRef]
- Mantovani, A.; Sozzani, S.; Allavena, P.; Locati, M.; Sica, A. Macrophage polarization: Tumor-associated macrophages as a paradigm for polarized m2 mononuclear phagocytes. Trends Immunol. 2002, 23, 549–555. [Google Scholar] [CrossRef]
- Zhang, J.J.; Li, Y.; Zhou, T.; Xu, D.P.; Pei, Z.; Li, S.; Li, H.B. Bioactivities and health benefits of mushrooms mainly from China. Molecules 2016, 21, 938–954. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, Y.; Zhang, L.; Tian, Q. Mushroom polysaccharide lentinan for treating different types of cancers: A review of 12 years clinical studies in China. Prog. Mol. Bio. Transl. Sci. 2019, 163, 297–328. [Google Scholar]
- Changyan, Z.; Yanru, Q.; Qingjiu, T.; Wei, J.; Yanfang, L. Purification and characterization of a novel small-molecule polysaccharide from the Maitake medicinal mushroom Grifola frondosa (higher Basidiomycetes). Int. J. Med. Mushrooms 2013, 15, 145. [Google Scholar]
- Ding, X.; Hou, Y.L.; Zhu, Y.X.; Wang, P.P.; Fu, L.; Zhu, H.Q.; Zhang, N. Structure elucidation, anticancer and antioxidant activities of a novel polysaccharide from Gomphus clavatus Gray. Oncol. Rep. 2015, 33, 3162–3170. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Methylated Product | Linkage | Unit (m/z) |
|---|---|---|
| 1,2,3,4-tetrakis-O-trimethylsilyl-β-DL-Arap | 1,4- | 45 59 73 103 117 147 169 191 204 217 230 243 259 291 305 333 |
| 1,2,3,4,6-pentakis-O-trimethylsilyl-D-Manp | 1,4,6- | 44 59 73 89 103 117 147 157 175 191 204 217 231 291 305 319 393 435 |
| 3,4,6-tri-O-methyl-2-O-trimethylsilyl-α-D-Glcp | 2- | 41 45 59 73 89 101 116 131 146 159 185 245 |
| 2,3,6-tri-O-methyl-1,4-bis-O-trimethylsilyl-Glcp | 1,4- | 29 45 59 73 88 101 133 146 159 175 201 217 232 |
| 3,4-di-O-methyl-1,2,6-tris-O-trimethylsilyl-Galp | 1,6- | 41 59 73 89 103 117 133 146 159 173 189 205 232 277 345 377 |
| Methylated Product | Linkage | Unit (m/z) |
|---|---|---|
| 1,2,3,4-tetrakis-O-trimethylsilyl-β-L-Arap | 1,4- | 15 31 45 59 73 101 116 133 147 169 191 204 217 231 259 291 305 333 |
| 6-O-methyl-1,2,3,4-tetrakis-O-trimethylsilyl-β-D-Glcp | 1,4- | 44 59 73 89 103 117 147 159 191 204 217 231 265 291 305 377 393 |
| 4,6-di-O-methyl-1,2,3-tris-O-trimethylsilyl-Glcp | 1- | 29 41 45 59 73 89 101 116 133 147 173 191 204 217 233 335 |
| 3,4-di-O-methyl-1,2,6-tris-O-trimethylsilyl-Galp | 1,2,6- | 41 59 73 89 103 117 133 146 159 173 189 205 232 277 345 377 |
| 2,4-di-O-methyl-1,3,6-tris-O-trimethylsilyl-Galp | 1,6- | 41 59 73 89 101 117 131 146 159 173 191 207 231 259 275 303 319 |
| Glycosyl Residues | Chemical Shifts (ppm) | |||||
|---|---|---|---|---|---|---|
| H1/C1 | H2/C2 | H3/C3 | H4/C4 | H5/C5 | H6/C6 | |
| →2)-α-D-Glcp (A) | 5.04/98.27 | 3.85/69.50 | 3.97/70.41 | 3.69/69.76 | 3.75/66.87 | 3.53/74.91 |
| (1→4,6)-D-Manp (B) | 4.96/101.69 | 3.69/71.75 | 4.00/68.26 | 3.53/74.91 | 3.85/66.80 | 3.65/68.26 |
| (1→6)-Galp (C) | 4.96/98.20 | 3.65/68.49 | 3.82/69.50 | 3.30/76.22 | 3.38/68.82 | 3.22/73.06 |
| (1→4)-Glcp (D) | 4.91/98.27 | 3.75/69.76 | 3.85/69.32 | 3.62/68.49 | 3.75/69.42 | 3.55/74.91 |
| (1→4)-Arap (E) | 4.42/103.00 | 3.22/72.89 | 3.38/68.82 | 3.30/69.50 | 3.65/69.17 | -- |
| Glycosyl Residues | Chemical Shifts (ppm) | |||||
|---|---|---|---|---|---|---|
| H1/C1 | H2/C2 | H3/C3 | H4/C4 | H5/C5 | H6/C6 | |
| (1→2,6)-Galp (A) | 5.22/99.14 | 3.65/68.39 | 3.96/68.29 | 3.55/69.44 | 3.89/69.51 | 3.73/71.71 |
| (1→2,6)-Galp (B) | 5.06/101.46 | 4.09/69.44 | 4.09/68.39 | 3.73/71.71 | 3.89/66.50 | 3.52/69.51 |
| (1→4)-Glcp (C) | 5.01/98.03 | 4.02/68.77 | 4.02/69.51 | 3.76/69.30 | 3.89/66.50 | 3.65/68.29 |
| (1→6)-Galp (D) | 4.95/101.36 | 3.65/68.39 | 4.09/68.29 | 3.55/69.51 | 3.94/66.14 | 3.49/66.78 |
| →1)-Glcp (E) | 4.87/97.89 | 3.73/69.44 | 3.89/69.30 | 3.58/68.29 | 3.73/69.44 | 3.49/67.20 |
| (1→4)-β-L-Arap (F) | 4.41/102.99 | 3.38/75.43 | 3.65/73.48 | 3.25/69.51 | 3.49/70.39 | -- |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Yang, T.; Huang, Q.; Li, B.; Ding, X.; Hou, Y. Comparative Studies on the Structure and Biological Activities of Two New Polysaccharides from Tricholoma sinoportentosum (TS-P) and Termitomyces albuminosus (TA-P). Polymers 2023, 15, 2227. https://doi.org/10.3390/polym15092227
Chen X, Yang T, Huang Q, Li B, Ding X, Hou Y. Comparative Studies on the Structure and Biological Activities of Two New Polysaccharides from Tricholoma sinoportentosum (TS-P) and Termitomyces albuminosus (TA-P). Polymers. 2023; 15(9):2227. https://doi.org/10.3390/polym15092227
Chicago/Turabian StyleChen, Xi, Tong Yang, Qinghua Huang, Biao Li, Xiang Ding, and Yiling Hou. 2023. "Comparative Studies on the Structure and Biological Activities of Two New Polysaccharides from Tricholoma sinoportentosum (TS-P) and Termitomyces albuminosus (TA-P)" Polymers 15, no. 9: 2227. https://doi.org/10.3390/polym15092227