
Structural Characterization and Anti-Inflammatory Activity of a Novel Polysaccharide from Duhaldea nervosa
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Extraction and Purification of Polysaccharides from the D. nervosa Roots
2.3. Molecular Weight and Homogeneity Determination
2.4. Monosaccharide Composition Analysis
2.5. Methylation Analysis
2.6. Fourier-Transform Infrared (FT-IR) and UV–Visible (UV–Vis) Spectroscopic Scanning Analysis
2.7. Nuclear Magnetic Resonance Spectroscopy Analysis
2.8. Cell Culture and Viability Assay
2.9. Detection of the Levels of Inflammatory Biomarkers
2.10. Statistical Analysis
3. Results
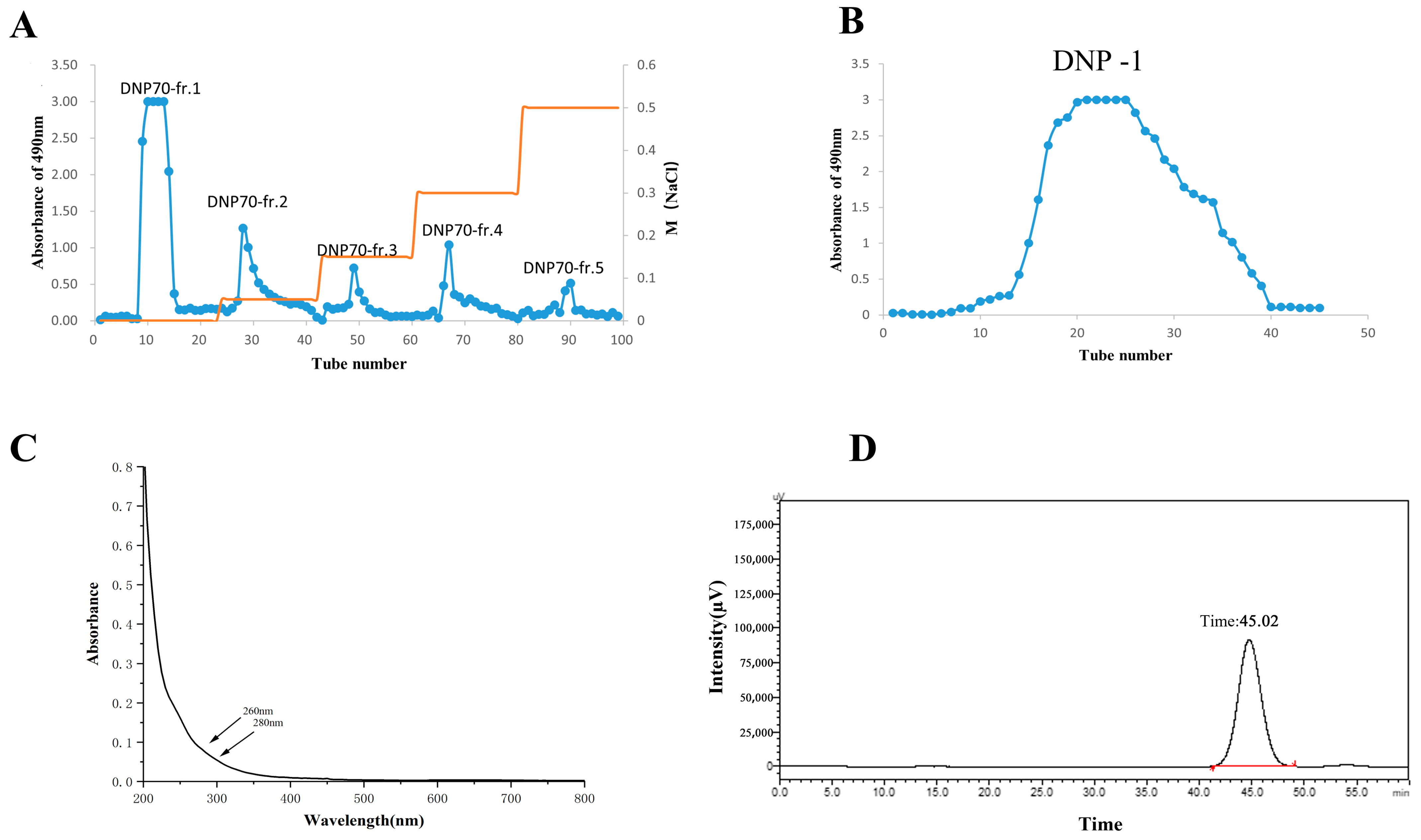
3.1. Extraction and Purification of Polysaccharides from the D. nervosa Roots
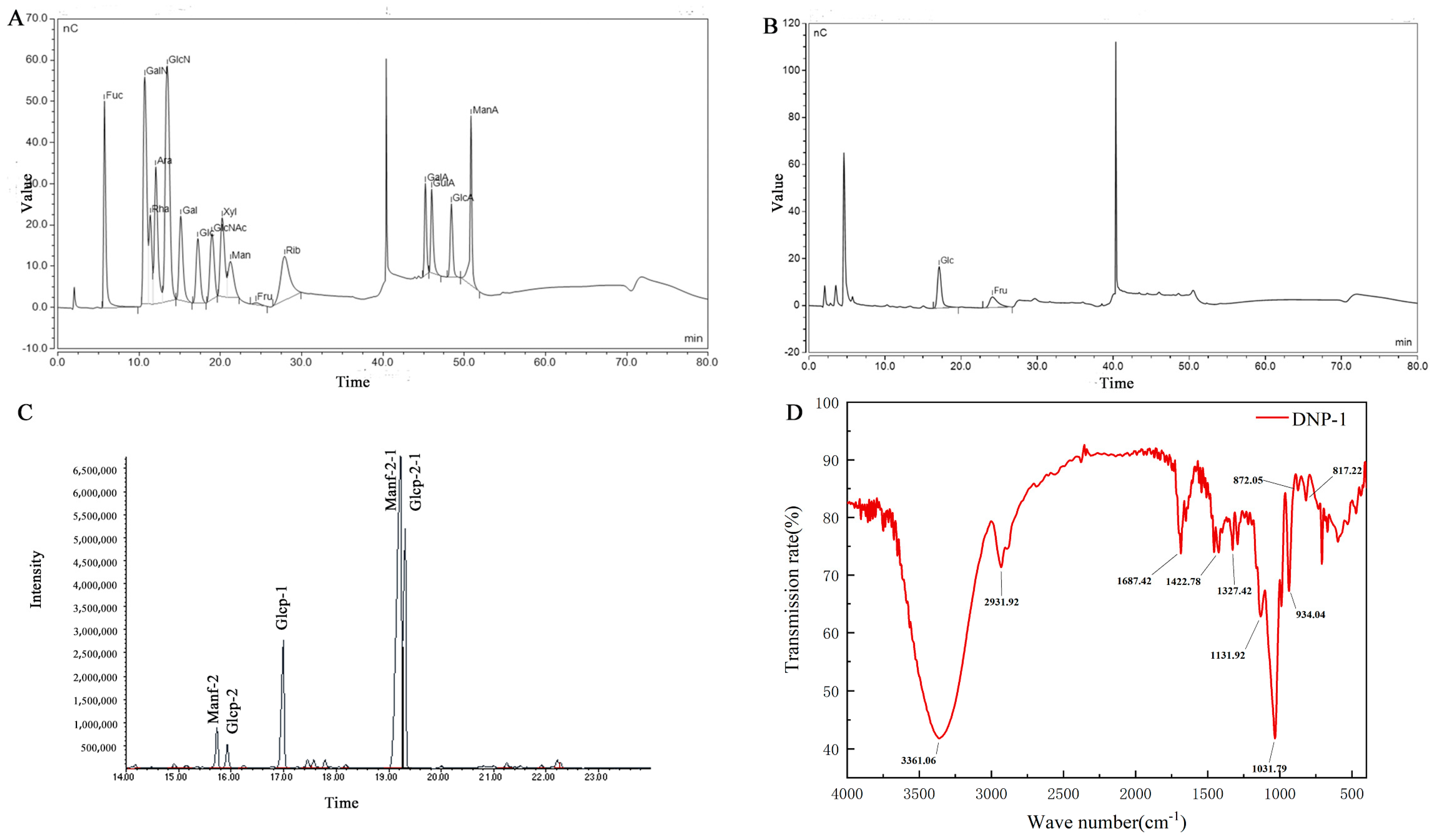
3.2. Molecular Weight and Monosaccharide Composition Analysis
3.3. Methylation Analysis
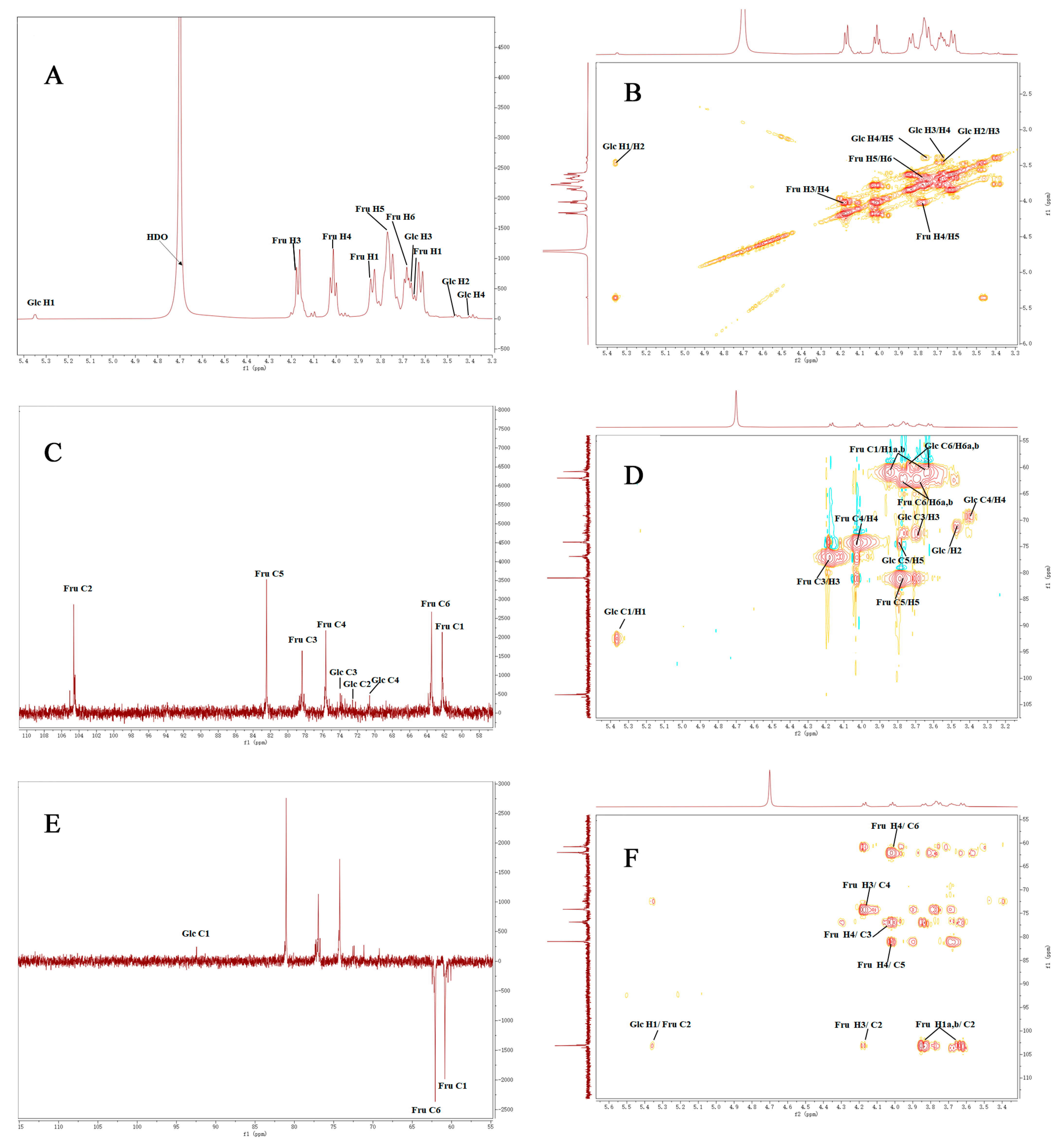
3.4. FT-IR Spectrum and NMR Analysis
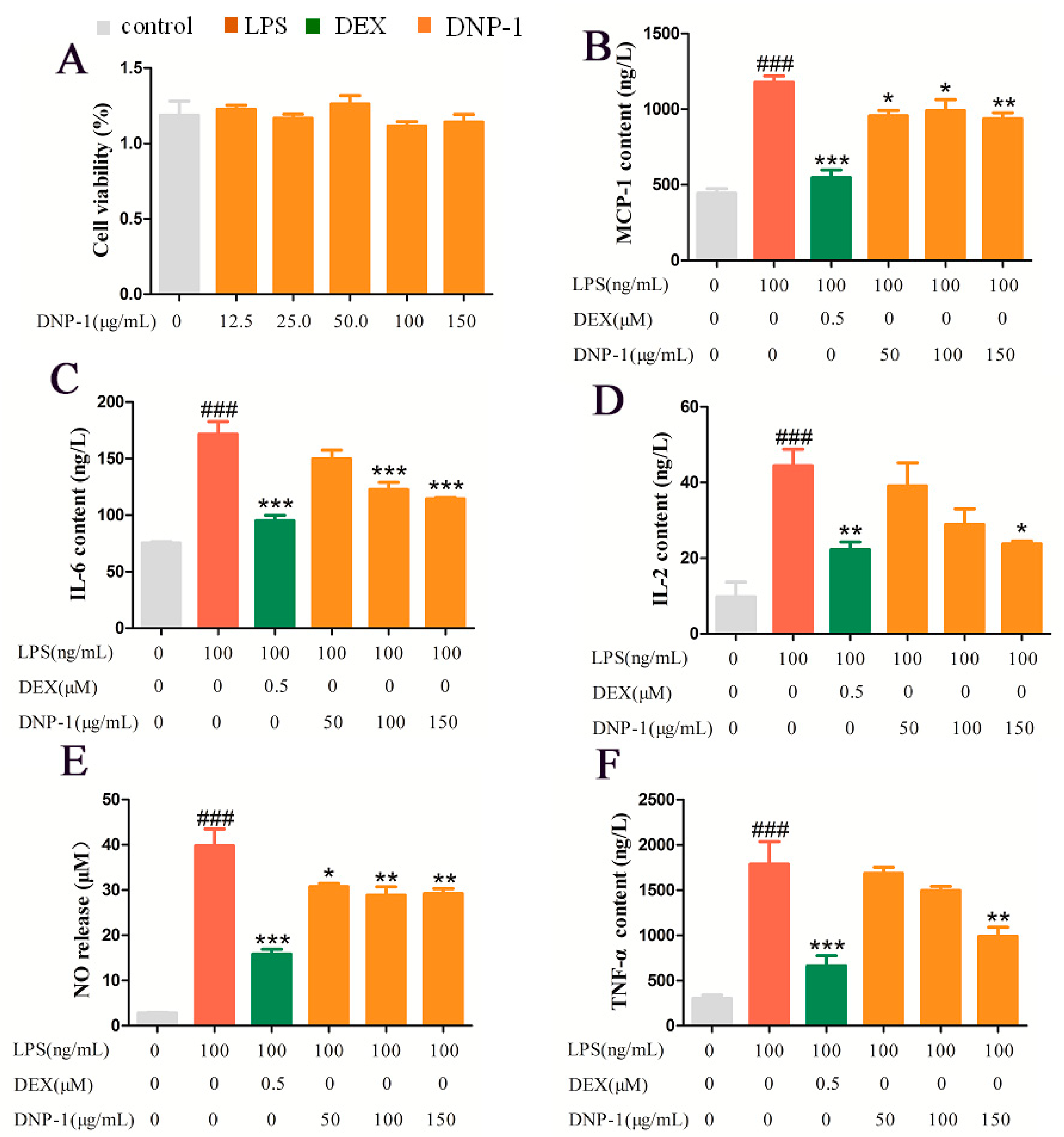
3.5. Effects of DNP−1 on RAW 264.7 Cell Viability
3.6. Effects of DNP−1 on LPS-Induced Pro-Inflammatory Cytokine Production in RAW264.7 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Duan, X.; Li, J.; Cui, J.; Wen, H.; Xin, X.; Aisa, H.A. A network pharmacology strategy combined with in vitro experiments to investigate the potential anti-inflammatory mechanism of Prunus cerasifera Ehrhart. J. Food Biochem. 2022, 46, e14396. [Google Scholar] [CrossRef]
- Guo, X.; Yu, X.; Zheng, B.; Zhang, L.; Zhang, F.; Zhang, Y.; Li, J.; Pu, G.; Zhang, L.; Wu, H. Network Pharmacology-Based Identification of Potential Targets of Lonicerae japonicae Flos Acting on Anti-Inflammatory Effects. Biomed. Res. Int. 2021, 2021, 5507003. [Google Scholar] [CrossRef]
- Marjoribanks, J.; Ayeleke, R.O.; Farquhar, C.; Proctor, M. Nonsteroidal anti-inflammatory drugs for dysmenorrhoea. Cochrane Database Syst. Rev. 2015, 2015, CD001751. [Google Scholar] [CrossRef] [PubMed]
- To, K.I.; Zhu, Z.X.; Wang, Y.N.; Li, G.A.; Sun, Y.M.; Li, Y.; Jin, Y.H. Integrative network pharmacology and experimental verification to reveal the anti-inflammatory mechanism of ginsenoside Rh4. Front. Pharmacol. 2022, 13, 953871. [Google Scholar] [CrossRef]
- Wang, J.; Chen, Q.; Sheng, R.; Li, P.; Liu, P.; Luo, J.; Zhong, Z.; Xu, S. Integration of transdermal chemistry and network pharmacology to decipher the mechanism of ShexiangZhuifeng analgesic plaster to treat rheumatoid arthritis. Phytomedicine 2023, 108, 154507. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Yilin, C.; Yousheng, C.; Yourun, L.; Shangwu, L.; Xuejun, G.; Tiangang, G.; Shixin, Z.; Ying, L.; Qiner, Y.; et al. ASTERACEAE (COMPOSITAE). In Flora of China; Science Press: Beijing, China, 2011; Volume 20–21, p. 548. [Google Scholar]
- Long, S. Clinical experience of “maoshoucai” for treating traumatic injury. J. Med. Pharm. Chin. Minor. 2004, 12, 231–232. [Google Scholar]
- Shen, J.; Li, Y.Z.; Yao, S.; Zhu, Z.W.; Wang, X.; Sun, H.H.; Ji, W.F. Effect of Duhaldea nervosa Extract on the Proliferation, Differentiation and Mineralization of Osteoblasts. Chin. J. Ethnomed. Ethnopharm. 2022, 31, 6. [Google Scholar]
- Guo, L.; Ma, R.; Sun, H.; Raza, A.; Tang, J.; Li, Z. Anti-Inflammatory Activities and Related Mechanism of Polysaccharides Isolated from Sargentodoxa cuneata. Chem. Biodivers. 2018, 15, e1800343. [Google Scholar] [CrossRef]
- Yan, L.; Cheng, X.R.; Zeng, Q.; Qin, J.J.; Zhang, W.D.; Jin, H.Z. Phytane and neoclerodane diterpenes from the aerial parts of Inula nervosa Wall. Biochem. Syst. Ecol. 2011, 39, 700–703. [Google Scholar] [CrossRef]
- Lan, Y.; Hui-Zi, J.; Li-Yue, N.; Jiang-Jiang, Q.; Jian-Jun, F.; Wei-Dong, Z. Chemical Constituents from Inula nervosa Wall. Nat. Prod. Res. Dev. 2011, 23, 258–261. [Google Scholar]
- Lianghong, L.; Jiayu, Z.; Binjie, Z.; Ying, G.; Liting, W.; Lei, C.; Wei, C. Rapid characterization of chlorogenic acids in Duhaldea nervosa based on ultra-high-performance liquid chromatography-linear trap quadropole-Orbitrap-mass spectrometry and mass spectral trees similarity filter technique. J. Sep. Sci. 2018, 41, 1764–1774. [Google Scholar] [CrossRef]
- Cai, W.; Li, K.-L.; Xiong, P.; Gong, K.-Y.; Zhu, L.; Yang, J.-B.; Wu, W.-H. A systematic strategy for rapid identification of chlorogenic acids derivatives in Duhaldea nervosa using UHPLC-Q-Exactive Orbitrap mass spectrometry. Arab. J. Chem. 2020, 13, 3751–3761. [Google Scholar] [CrossRef]
- Pinghuai, L.; Chunniu, W.; Xun, Y.; Yifei, H. Preliminary Test of Chemical Components and Content Determination of Total Flavonoids in Different Parts of Gonocaryum lobbianum. Med. Plant 2013, 4, 46–50. [Google Scholar]
- Tang, N.; Jiang, Y.; Zhu, J.; Zhou, L.; Cai, W.; Ning, Q. Preparation of Chitosan-based Whitening and Anti-inflammatory Films Using Angelica Dahurica Benth and Duhaldea nervosa. Shandong Chem. Ind. 2019, 48, 14–18. [Google Scholar] [CrossRef]
- Liu, Y.; Ye, Y.; Hu, X.; Wang, J. Structural characterization and anti-inflammatory activity of a polysaccharide from the lignified okra. Carbohydr. Polym. 2021, 265, 118081. [Google Scholar] [CrossRef] [PubMed]
- Suisui, J.; Huina, X.; Chunhui, Z.; Feng, Z.; Duo, L. Oyster polysaccharides relieve DSS-induced colitis via anti-inflammatory and maintaining the physiological hypoxia. Int. J. Biol. Macromol. 2023, 238, 124150. [Google Scholar] [CrossRef]
- ZhiZhi, Z.; Jie, Z.; Zhou, H.; WeiLian, B.; LiShuang, Z.; Yang, L.; Ji, D.; DaoFeng, C.; Yan, L. Structural characterization and anti-inflammatory activities of anti-complementary polysaccharides from Rhododendron principis. Planta Med. 2023. [Google Scholar] [CrossRef]
- Zhang, J.; Song, Z.; Li, Y.; Zhang, S.; Bao, J.; Wang, H.; Dong, C.; Ohizumi, Y.; Xu, J.; Guo, Y. Structural analysis and biological effects of a neutral polysaccharide from the fruits of Rosa laevigata. Carbohydr. Polym. 2021, 265, 118080. [Google Scholar] [CrossRef]
- An, Q.; Ye, X.; Han, Y.; Zhao, M.; Chen, S.; Liu, X.; Li, X.; Zhao, Z.; Zhang, Y.; Ouyang, K.; et al. Structure analysis of polysaccharides purified from Cyclocarya paliurus with DEAE-Cellulose and its antioxidant activity in RAW264.7 cells. Int. J. Biol. Macromol. 2020, 157, 604–615. [Google Scholar] [CrossRef]
- Gao, H.; Wen, J.J.; Hu, J.L.; Nie, Q.X.; Chen, H.H.; Xiong, T.; Nie, S.P.; Xie, M.Y. Polysaccharide from fermented Momordica charantia L. with Lactobacillus plantarum NCU116 ameliorates type 2 diabetes in rats. Carbohydr. Polym. 2018, 201, 624–633. [Google Scholar] [CrossRef]
- Ceole, L.F.; Companhoni, M.V.P.; Sanches Lopes, S.M.; de Oliveira, A.J.B.; Goncalves, R.A.C.; Dias Filho, B.P.; Nakamura, C.V.; Ueda-Nakamura, T. Anti-herpes activity of polysaccharide fractions from Stevia rebaudiana leaves. Nat. Prod. Res. 2020, 34, 1558–1562. [Google Scholar] [CrossRef]
- Wang, L.; Liu, H.M.; Qin, G.Y. Structure characterization and antioxidant activity of polysaccharides from Chinese quince seed meal. Food Chem. 2017, 234, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Sevag, M.G.; Lackman, D.B.; Smolens, J. The Isolation of the Components of Streptococcal Nucleoproteins in Serologically Active Form. J. Biol. Chem. 1938, 124, 425–436. [Google Scholar] [CrossRef]
- Liu, Y.; Xu, Z.; Zhang, W.; Duan, J.; Jiang, J.; Sun, D. Characterization of fractional polysaccharides from Gleditsia sinensis and Gleditsia microphylla gums. Molecules 2016, 21, 1745. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Yan, X.; Liang, J.; Li, S.; He, H.; Xiong, Q.; Lai, X.; Hou, S.; Huang, S. Comparison of different extraction methods for polysaccharides from Dendrobium officinale stem. Carbohydr. Polym. 2018, 198, 101–108. [Google Scholar] [CrossRef]
- Cao, J.-J.; Lv, Q.-Q.; Zhang, B.; Chen, H.-Q. Structural characterization and hepatoprotective activities of polysaccharides from the leaves of Toona sinensis (A. Juss) Roem. Carbohydr. Polym. 2019, 212, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Tim, M. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Biluca, F.C.; Silva, B.D.; Caon, T.; Mohr, E.T.B.; Vieira, G.N.; Gonzaga, L.V.; Vitali, L.; Micke, G.; Fett, R.; Dalmarco, E.M.; et al. Investigation of phenolic compounds, antioxidant and anti-inflammatory activities in stingless bee honey (Meliponinae). Food Res. Int. 2020, 129, 108756. [Google Scholar] [CrossRef]
- Pettolino, F.A.; Walsh, C.; Fincher, G.B.; Bacic, A. Determining the polysaccharide composition of plant cell walls. Nat. Protoc. 2012, 7, 1590–1607. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Zhang, J.; Gao, F.; Zhou, J.; Xiang, Z.; Zhou, C.; Wan, L.; Chen, J. Structure features and in vitro hypoglycemic activities of polysaccharides from different species of Maidong. Carbohydr. Polym. 2017, 173, 215–222. [Google Scholar] [CrossRef]
- Wang, C.; Hua, D.; Yan, C. Structural characterization and antioxidant activities of a novel fructan from Achyranthes bidentata Blume, a famous medicinal plant in China. Ind. Crops Prod. 2015, 70, 427–434. [Google Scholar] [CrossRef]
- Xu, J.; Chen, D.; Liu, C.; Wu, X.-Z.; Dong, C.-X.; Zhou, J. Structural characterization and anti-tumor effects of an inulin-type fructan from Atractylodes chinensis. Int. J. Biol. Macromol. 2016, 82, 765–771. [Google Scholar] [CrossRef]
- Chen, J.; Cheong, K.L.; Song, Z.; Shi, Y.; Huang, X. Structure and protective effect on UVB-induced keratinocyte damage of fructan from white garlic. Carbohydr. Polym. 2013, 92, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Wang, C.; Hou, X.; Yan, C. Structural characterization and osteoprotective effects of a polysaccharide purified from Achyranthes bidentata. Int. J. Biol. Macromol. 2019, 139, 1063–1073. [Google Scholar] [CrossRef]
- Kang, J.; Cui, S.W.; Phillips, G.O.; Chen, J.; Guo, Q.; Wang, Q. New Studies on Gum Ghatti (Anogeissus Latifolia) Part III: Structure Characterization of A Globular Polysaccharide Fraction By ID, 2D NMR Spectroscopy And Methylation Analysis. Food Hydrocoll. 2011, 25, 1999–2007. [Google Scholar] [CrossRef]
- Sun, Q.; Zhu, L.; Li, Y.; Cui, Y.; Jiang, S.; Tao, N.; Chen, H.; Zhao, Z.; Xu, J.; Dong, C. A novel inulin-type fructan from Asparagus cochinchinensis and its beneficial impact on human intestinal microbiota. Carbohydr. Polym. 2020, 247, 116761. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, P.; Zhang, X.; Li, X. Chemical structure elucidation of an inulin-type fructan isolated from Lobelia chinensis lour with anti-obesity activity on diet-induced mice. Carbohydr. Polym. 2020, 240, 116357. [Google Scholar] [CrossRef] [PubMed]
- Pontes, A.G.O.; Silva, K.L.; Fonseca, S.G.d.C.; Soares, A.A.; Feitosa, J.P.d.A.; Braz-Filho, R.; Romero, N.R.; Bandeira, M.A.M. Identification and determination of the inulin content in the roots of the Northeast Brazilian species Pombalia calceolaria L. Carbohydr. Polym. 2016, 149, 391–398. [Google Scholar] [CrossRef]
- Meng, Y.; Xu, Y.; Chang, C.; Qiu, Z.; Hu, J.; Wu, Y.; Zhang, B.; Zheng, G. Extraction, characterization and anti-inflammatory activities of an inulin-type fructan from Codonopsis pilosula. Int. J. Biol. Macromol. 2020, 163, 1677–1686. [Google Scholar] [CrossRef]
- Xin, S.; Chongzhen, S.; Xin, T.; Xiaosa, Z.; Duo, H.; Shan, L.; Rong, Q.; Xiaodan, H.; Yangwei, S.; Linhui, H.; et al. Anti-Inflammatory and Intestinal Microbiota Modulation Properties of Jinxiang Garlic (Allium sativum L.) Polysaccharides toward Dextran Sodium Sulfate-Induced Colitis. J. Agric. Food Chem. 2020, 68, 12295–12309. [Google Scholar] [CrossRef]
- Dong, C.X.; Zhang, L.J.; Xu, R.; Zhang, G.; Zhou, Y.B.; Han, X.Q.; Zhang, Y.; Sun, Y.X. Structural characterization and immunostimulating activity of a levan-type fructan from Curcuma kwangsiensis. Int. J. Biol. Macromol. 2015, 77, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Shaw, P.E.; Tatum, J.H.; Berry, R.E. Acid-catalyzed degradation of d-fructose. Carbohydr. Res. 1967, 5, 266–273. [Google Scholar] [CrossRef]
- Caleffi, E.R.; Krausova, G.; Hyrslova, I.; Paredes, L.L.; dos Santos, M.M.; Sassaki, G.L.; Goncalves, R.A.; de Oliveira, A.J. Isolation and prebiotic activity of inulin-type fructan extracted from Pfaffia glomerata (Spreng) Pedersen roots. Int. J. Biol. Macromol. 2015, 80, 392–399. [Google Scholar] [CrossRef]
- Vongchan, P.; Linhardt, R.J. Characterization of a new monoclonal anti-glypican-3 antibody specific to the hepatocellular carcinoma cell line, HepG2. World J. Hepatol. 2017, 9, 368–384. [Google Scholar] [CrossRef]
- Wahlstrom, R.; Rovio, S.; Suurnakki, A. Analysis of mono- and oligosaccharides in ionic liquid containing matrices. Carbohydr. Res. 2013, 373, 42–51. [Google Scholar] [CrossRef]
- Sun, Y.; Huo, J.; Zhong, S.; Zhu, J.; Li, Y.; Li, X. Chemical structure and anti-inflammatory activity of a branched polysaccharide isolated from Phellinus baumii. Carbohydr. Polym. 2021, 268, 118214. [Google Scholar] [CrossRef]
- Hou, C.; Chen, L.; Yang, L.; Ji, X. An insight into anti-inflammatory effects of natural polysaccharides. Int. J. Biol. Macromol. 2020, 153, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Epelman, S.; Lavine, K.J.; Randolph, G.J. Origin and functions of tissue macrophages. Immunity 2014, 41, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Davies, L.C.; Taylor, P.R. Tissue-resident macrophages: Then and now. Immunology 2015, 144, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.; Liu, W.; Song, X.; Qi, Y.; Zhang, C.; Gao, Z.; Zhang, J.; Jia, L. Antioxidant and anti-inflammation of enzymatic-hydrolysis residue polysaccharides by Lentinula edodes. Int. J. Biol. Macromol. 2018, 120, 811–822. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zhu, X.; Ma, J.; Zhang, M.; Wu, H. Structural Elucidation of a Novel Pectin-Polysaccharide from the Petal of Saussurea laniceps and the Mechanism of its Anti-HBV Activity. Carbohydr. Polym. 2019, 223, 115077. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Methylated Sugar | Mass Fragments (m/z) | Molar Ratios (%) | Deduced Linkage |
|---|---|---|---|
| 1,3,4,6-Me4 Manf/Glcf | 87, 101, 129, 145, 161 | 2.1 | Fruf-(2→ |
| 2,3,4,6-Me4-Glcp | 43, 71, 87, 101, 117, 129, 145, 161, 205 | 4.72 | Glcp-(1→ |
| 3,4,6-Me3-Manf/Glcf | 43, 71, 87, 99, 101, 129, 145, 161, 189 | 28.70 | →1)-Fruf-(2→ |
| Glycosyl Residues | C1/H1a, b | C2/H2 | C3/H3 | C4/H4 | C5/H5 | C6/H6a, b |
|---|---|---|---|---|---|---|
| α-D-Glcp-1 | 93.97/5.35 | 72.60/3.47 | 73.99/3.68 | 70.60/3.40 | 75.65/3.79 | 61.68/3.72,3.62 |
| β-D-Fruf-2,1 | 61.78/3.85,3.65 | 104.61/ns | 78.30/4.18 | 75.63/4.01 | 82.41/3.80 | 63.88/3.68,3.76 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Ma, X.; Shi, S.; He, S.; Li, J.; Wilson, G.; Cai, W.; Liu, L. Structural Characterization and Anti-Inflammatory Activity of a Novel Polysaccharide from Duhaldea nervosa. Polymers 2023, 15, 2081. https://doi.org/10.3390/polym15092081
Wang Z, Ma X, Shi S, He S, Li J, Wilson G, Cai W, Liu L. Structural Characterization and Anti-Inflammatory Activity of a Novel Polysaccharide from Duhaldea nervosa. Polymers. 2023; 15(9):2081. https://doi.org/10.3390/polym15092081
Chicago/Turabian StyleWang, Ziming, Xueqin Ma, Silin Shi, Shuo He, Jian Li, Gidion Wilson, Wei Cai, and Lianghong Liu. 2023. "Structural Characterization and Anti-Inflammatory Activity of a Novel Polysaccharide from Duhaldea nervosa" Polymers 15, no. 9: 2081. https://doi.org/10.3390/polym15092081