

Characterization of Cellulose-Degrading Bacteria Isolated from Silkworm Excrement and Optimization of Its Cellulase Production
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture Media and Sample Collection
2.2. Primary Screening of Cellulolytic Bacteria
2.3. Rescreening of Cellulolytic Bacteria
2.4. Quantification of Cellulase Activity
2.5. Identification of Cellulolytic Bacteria and Growth Curve
2.6. Optimization of Cellulase Production
2.7. Bioaugmentation of Silkworm Excrement with DC-11
2.8. Statistical Analysis
3. Results
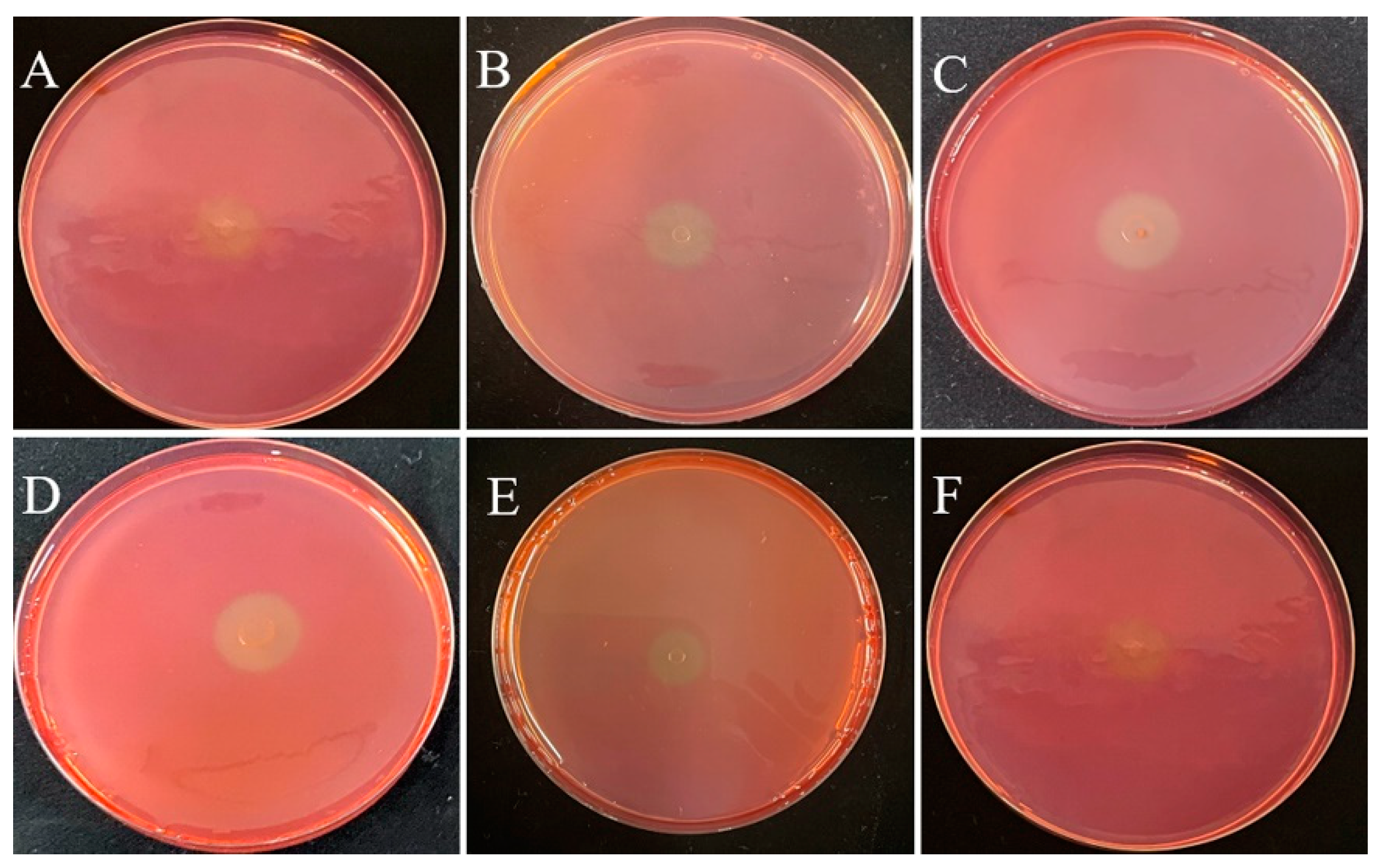
3.1. Isolation and Screening of Cellulolytic Bacteria
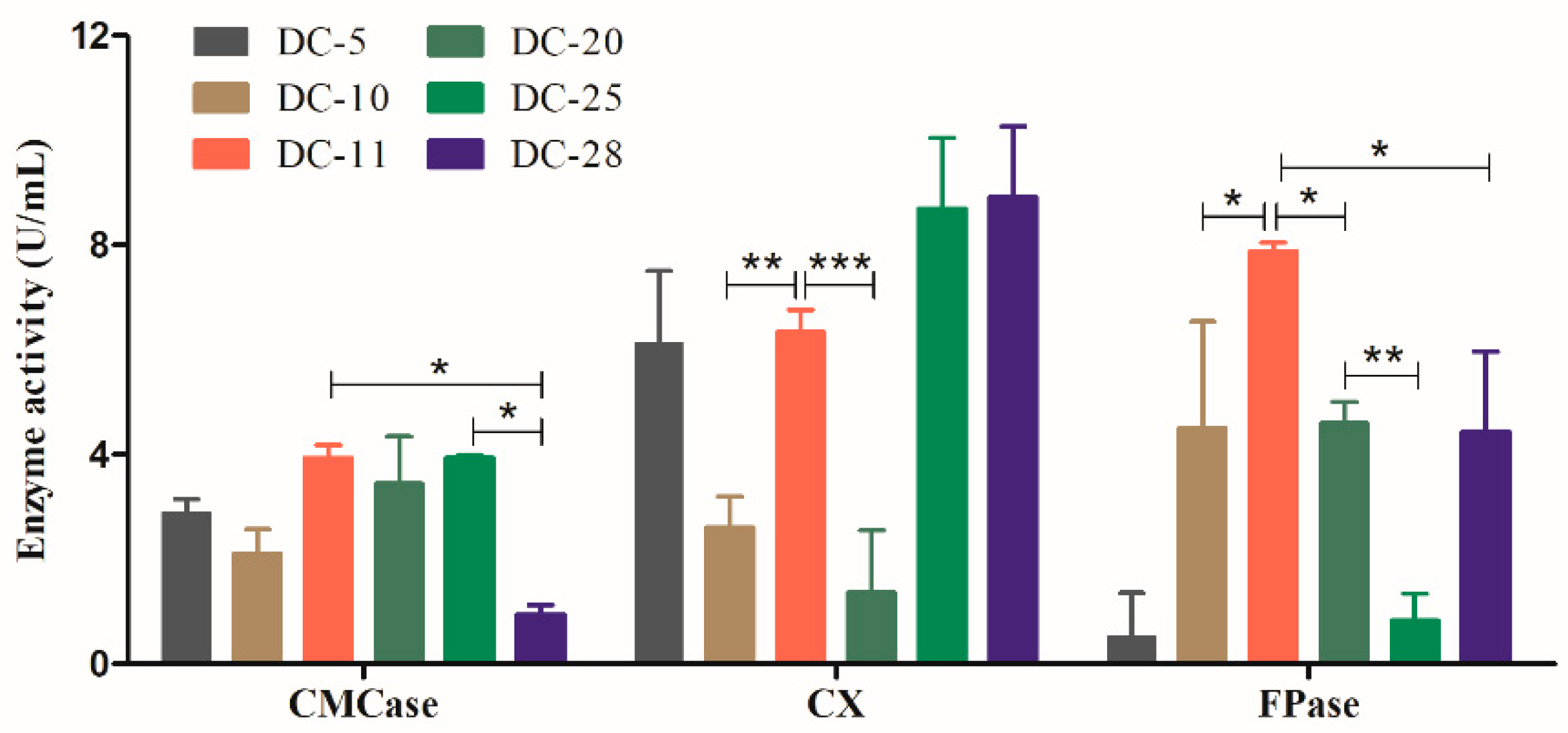
3.2. Quantification of Cellulase Activity for the Rescreening of Cellulolytic Bacteria
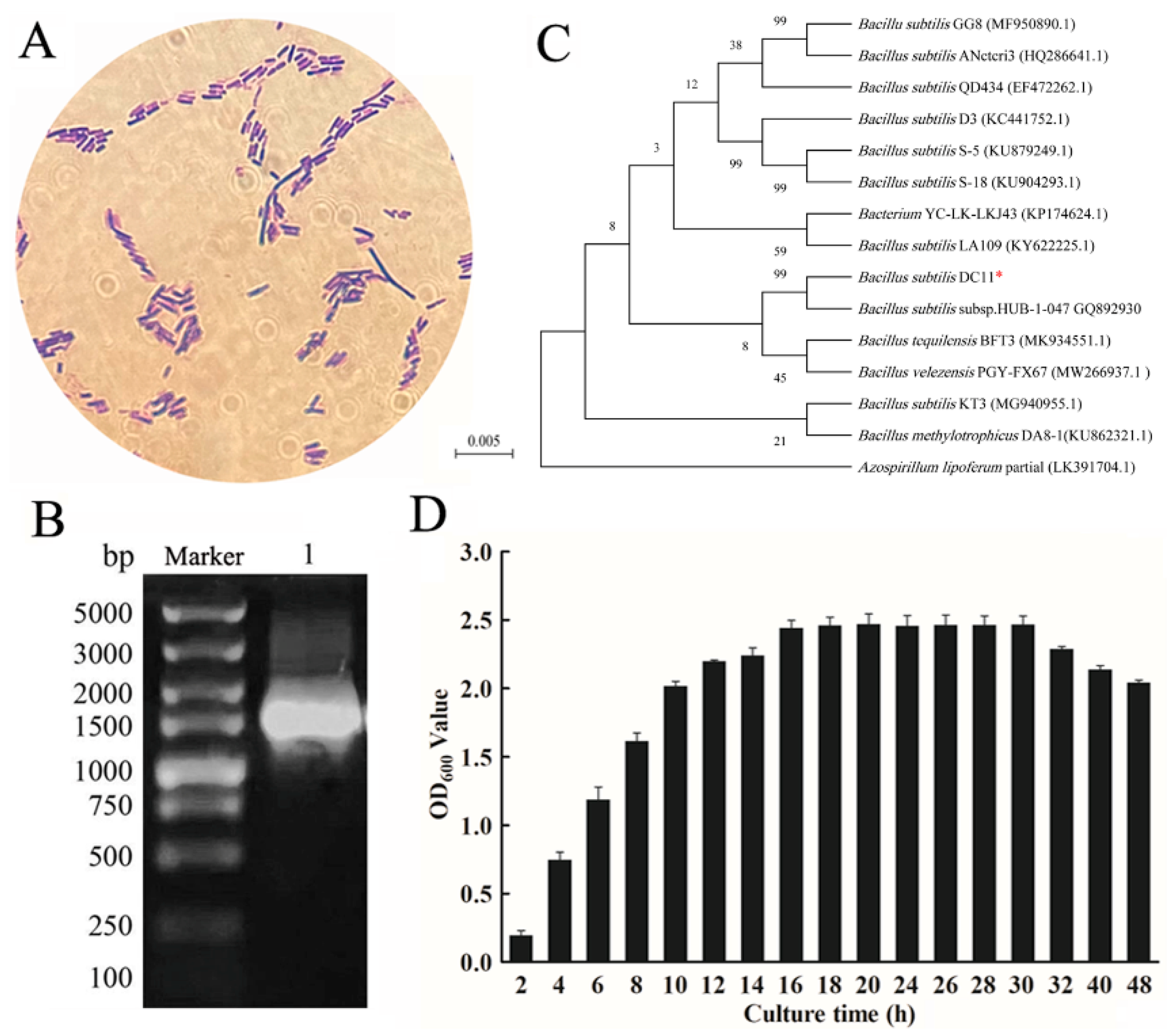
3.3. Identification of the Isolated Cellulolytic Bacteria
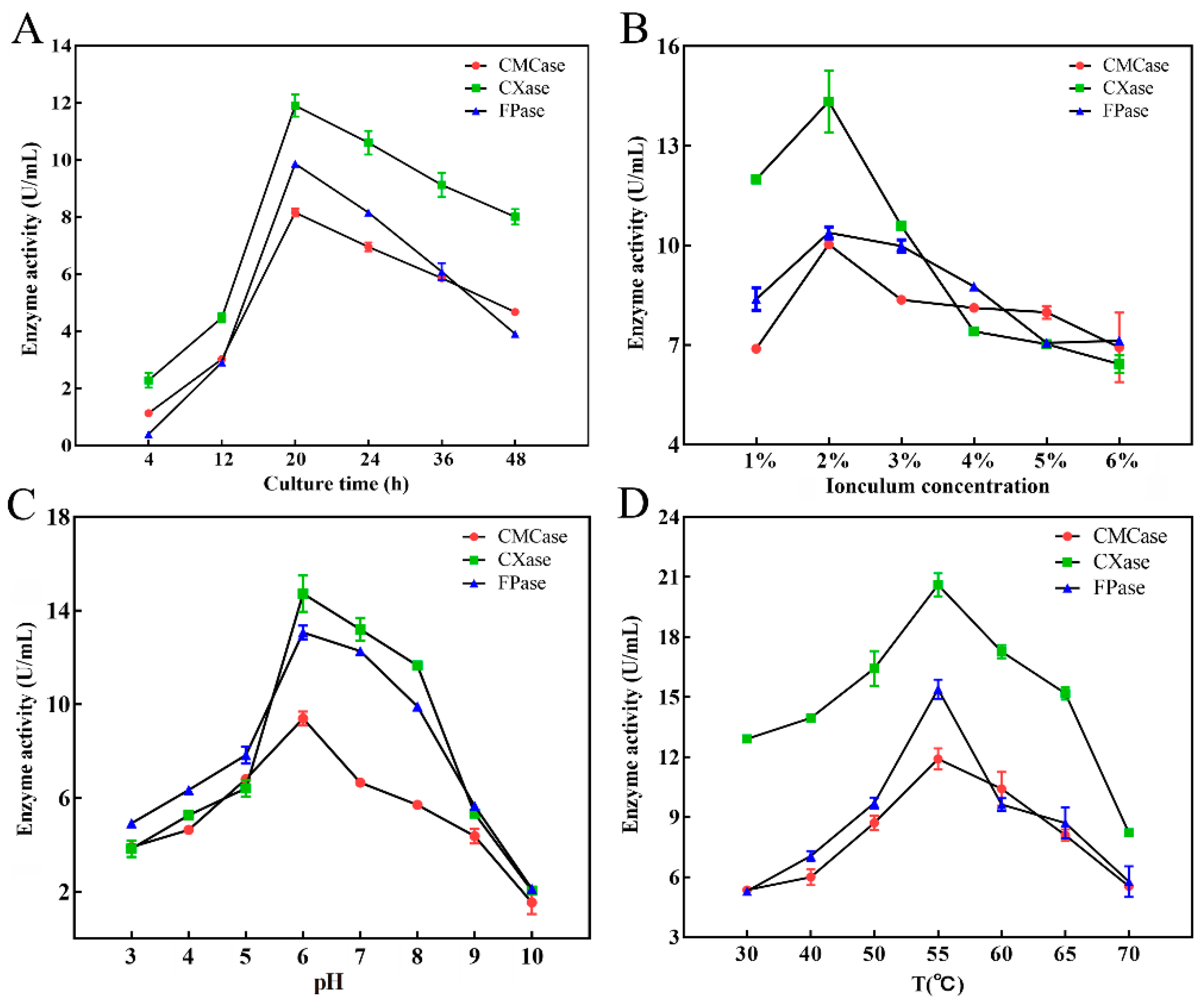
3.4. Optimization of Conditions for Cellulase Production by Strain DC-11
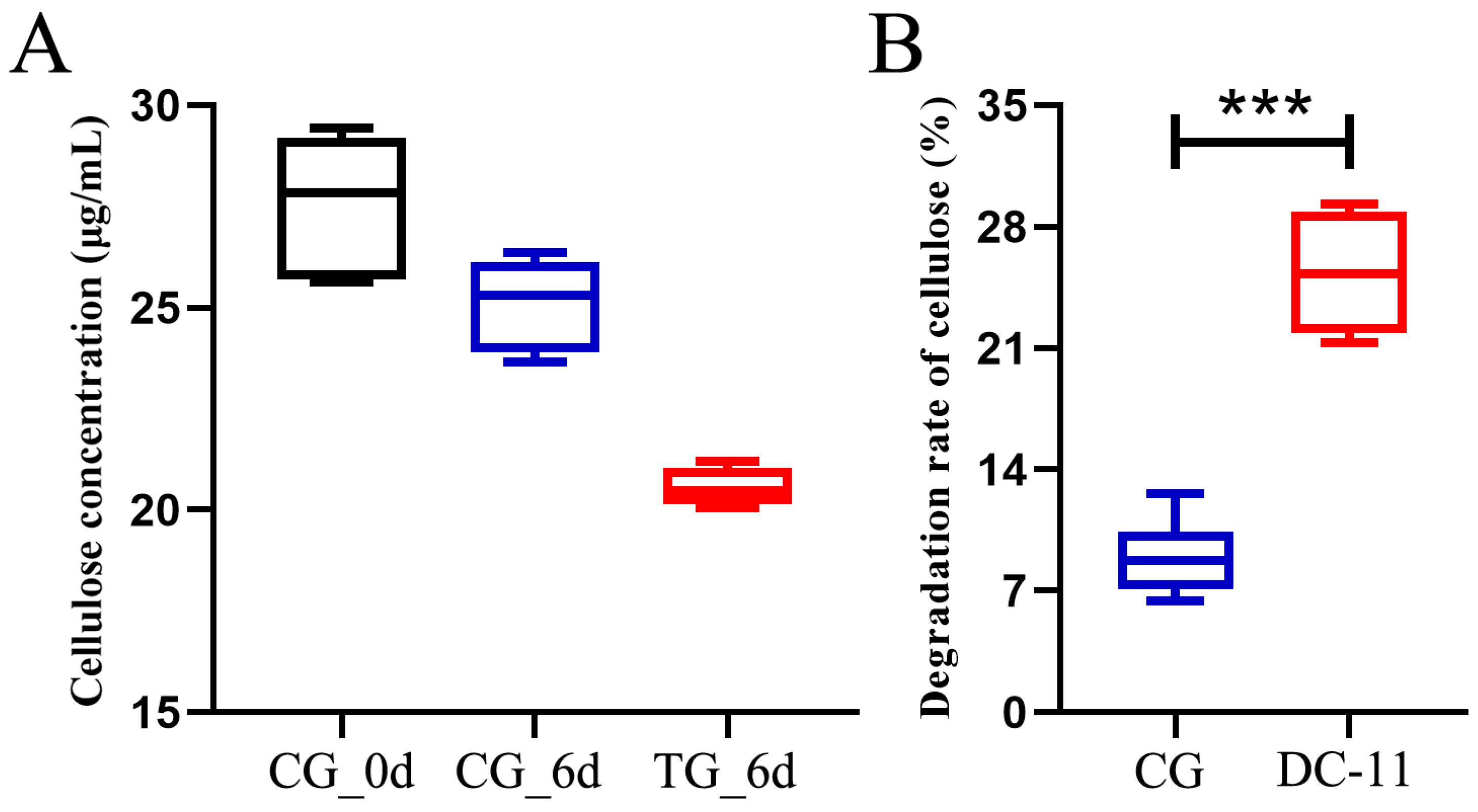
3.5. Bioaugmentation with DC-11 Enhanced the Degradation of Silkworm Excrement
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hussain, A.A.; Abdel-Salam, M.S.; Abo-Ghalia, H.H.; Hegazy, W.K.; Hafez, S.S. Optimization and molecular identification of novel cellulose degrading bacteria isolated from Egyptian environment. J. Genet. Eng. Biotechnol. 2017, 15, 77–85. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Wang, R.X.; Zhou, J.S.; Cheng, J.F.; Li, Y.H. An assessment of the genomics, comparative genomics and cellulose degradation potential of Mucilaginibacter polytrichastri strain RG4-7. Bioresour. Technol. 2020, 297, 122389. [Google Scholar] [CrossRef]
- Bajpai, P. Paper Bioprocessing. In Biermann’s Handbook of Pulp and Paper; Elsevier: Amsterdam, The Netherlands, 2018; pp. 187–205. [Google Scholar]
- Kumar, D.; Murthy, G.S. Development and validation of a stochastic molecular model of cellulose hydrolysis by action of multiple cellulase enzymes. Bioresour. Bioprocess. 2017, 4, 54. [Google Scholar] [CrossRef]
- Egwuatu, T.; Appeh, O. Isolation and characterization of filter paper degrading bacteria from the guts of coptotermes formosanus. J. Bioremediat. Biodegrad. 2018, 9, 20–26. [Google Scholar] [CrossRef]
- Harnvoravongchai, P.; Singwisut, R.; Ounjai, P.; Aroonnual, A.; Kosiyachinda, P.; Janvilisri, T.; Chankhamhaengdecha, S. Isolation and characterization of thermophilic cellulose and hemicellulose degrading bacterium, Thermoanaerobacterium sp. R63 from tropical dry deciduous forest soil. PLoS ONE 2020, 15, e0236518. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Yu, S.M. Flavobacterium nackdongense sp. nov., a cellulose-degrading bacterium isolated from sediment. Arch. Microbiol. 2020, 202, 591–595. [Google Scholar] [CrossRef] [PubMed]
- Paul, M.; Mohapatra, S.; Kumar Das Mohapatra, P.; Thatoi, H. Microbial cellulases—An update towards its surface chemistry, genetic engineering and recovery for its biotechnological potential. Bioresour. Technol. 2021, 340, 125710. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R. Functional characterization of cellulose-degrading AA9 lytic polysaccharide monooxygenases and their potential exploitation. Appl. Microbiol. Biotechnol. 2020, 104, 3229–3243. [Google Scholar] [CrossRef]
- Manzoor, N.; Cao, L.; Deng, D.; Liu, Z.; Jiang, Y.; Liu, Y. Cellulase extraction from cellulolytic bacteria promoting bioelectricity production by degrading cellulose. J. Electroanal. Chem. 2018, 829, 241–248. [Google Scholar] [CrossRef]
- Ariffin, H.; Hassan, M.A.; Shah, U.K.; Abdullah, N.; Ghazali, F.M.; Shirai, Y. Production of bacterial endoglucanase from pretreated oil palm empty fruit bunch by Bacillus pumilus EB3. J. Biosci. Bioeng. 2008, 106, 231–236. [Google Scholar] [CrossRef]
- Rastogi, G.; Bhalla, A.; Adhikari, A.; Bischoff, K.M.; Hughes, S.R.; Christopher, L.P.; Sani, R.K. Characterization of thermostable cellulases produced by Bacillus and Geobacillus strains. Bioresour. Technol. 2010, 101, 8798–8806. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Maiti, T.K.; Roy, R.N. Production, purification, and characterization of cellulase from Acinetobacter junii GAC 16.2, a novel cellulolytic gut isolate of Gryllotalpa africana, and its effects on cotton fiber and sawdust. Ann. Microbiol. 2020, 70, 28. [Google Scholar] [CrossRef]
- Gupta, P.; Samant, K.; Sahu, A. Isolation of cellulose-degrading bacteria and determination of their cellulolytic potential. Int. J. Microbiol. 2012, 2012, 578925. [Google Scholar] [CrossRef]
- McDonald, J.E.; Rooks, D.J.; McCarthy, A.J. Methods for the isolation of cellulose-degrading microorganisms. Methods Enzymol. 2012, 510, 349–374. [Google Scholar] [CrossRef] [PubMed]
- Bano, S.; Qader, S.A.; Aman, A.; Syed, M.N.; Durrani, K. High production of cellulose degrading endo-1,4-beta-D-glucanase using bagasse as a substrate from Bacillus subtilis KIBGE HAS. Carbohydr. Polym. 2013, 91, 300–304. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.S.; Fernando, S.; Ross, B.; Wu, J.; Qin, W. Endoglucanase (EG) activity assays. Methods Mol. Biol. 2018, 1796, 169–183. [Google Scholar] [CrossRef]
- Karthika, A.; Seenivasagan, R.; Kasimani, R.; Babalola, O.O.; Vasanthy, M. Cellulases of an alkalophilic Bacillus strain isolated from soil. Can. J. Microbiol. 1984, 30, 774–779. [Google Scholar]
- Li, F.; Xie, Y.; Gao, X.; Shan, M.; Sun, C.; Niu, Y.D.; Shan, A. Screening of cellulose degradation bacteria from Min pigs and optimization of its cellulase production. Electron. J. Biotechn. 2020, 48, 29–35. [Google Scholar] [CrossRef]
- Adams, A.S.; Jordan, M.S.; Adams, S.M.; Suen, G.; Goodwin, L.A.; Davenport, K.W.; Currie, C.R.; Raffa, K.F. Cellulose-degrading bacteria associated with the invasive woodwasp Sirex noctilio. ISME J. 2011, 5, 1323–1331. [Google Scholar] [CrossRef]
- Irfan, M.; Safdar, A.; Syed, Q.; Nadeem, M. Isolation and screening of cellulolytic bacteria from soil and optimization of cellulase production and activity. Turk. J. Biochem. 2012, 37, 287–293. [Google Scholar] [CrossRef]
- Islam, F.; Roy, N. Screening, purification and characterization of cellulase from cellulase producing bacteria in molasses. BMC Res. 2018, 11, 445. [Google Scholar] [CrossRef] [PubMed]
- Anu; Kumar, S.; Kumar, A.; Kumar, V.; Singh, B. Optimization of cellulase production by Bacillus subtilis subsp. subtilis JJBS300 and biocatalytic potential in saccharification of alkaline-pretreated rice straw. Prep. Biochem. Biotechnol. 2021, 51, 697–704. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Cheng, W.; Li, S.; Tang, X.; Wei, Z. The “quality” and “quantity” of microbial species drive the degradation of cellulose during composting. Bioresour. Technol. 2021, 320, 124425. [Google Scholar] [CrossRef] [PubMed]
- Mei, J.; Shen, X.; Gang, L.; Xu, H.; Wu, F.; Sheng, L. A novel lignin degradation bacteria-Bacillus amyloliquefaciens SL-7 used to degrade straw lignin efficiently. Bioresour. Technol. 2020, 310, 123445. [Google Scholar] [CrossRef]
- Jain, D.; Ravina; Bhojiya, A.; Chauhan, S.; Rajpurohit, D.; Mohanty, S.R. Polyphasic Characterization of Plant Growth Promoting Cellulose Degrading Bacteria Isolated from Organic Manures. Curr. Microbiol. 2021, 78, 739–748. [Google Scholar] [CrossRef]
- Singh, S.; Thavamani, P.; Megharaj, M.; Naidu, R. Multifarious activities of cellulose degrading bacteria from Koala (Phascolarctos cinereus) faeces. J. Anim. Sci. Technol. 2015, 57, 23. [Google Scholar] [CrossRef]
- Khosravi, F.; Khaleghi, M.; Naghavi, H. Screening and identification of cellulose-degrading bacteria from soil and leaves at Kerman province, Iran. Arch. Microbiol. 2021, 204, 88. [Google Scholar] [CrossRef]
- Tabassum, R.; Bashir, S.; Raza, A. Evaluation of cellulases and xylanases production from Bacillus spp. isolated from buffalo digestive system. Kafkas Univ. Vet. Fak. Derg. 2018, 25, 39–46. [Google Scholar] [CrossRef]
- Qaisar, S.; Zohra, R.R.; Aman, A.; Qader, S.A. Enhanced production of cellulose degrading CMCase by newly isolated strain of Aspergillus versicolor. Carbohydr. Polym. 2014, 104, 199–203. [Google Scholar] [CrossRef]
- Gautam, R.; Sharma, J. Optimization, purification of cellulase produced from Bacillus subtilis sp. inaquosorum under solid state fermentation and its potential applications in denim industry. Int. J. Sci. Res. 2014, 3, 1759–1763. [Google Scholar]
- Pant, G.; Prakash, A.; Pavani, J.V.P.; Bera, S.; Deviram, G.V.N.S.; Kumar, A.; Panchpuri, M.; Prasuna, R.G. Production, optimization and partial purification of protease from Bacillus subtilis. J. Taibah. Univ. Sci. 2018, 9, 50–55. [Google Scholar] [CrossRef]
- Kumari, P.; Sayas, T.; Bucki, P.; Brown-Miyara, S.; Kleiman, M. Real-Time Visualization of Cellulase Activity by Microorganisms on Surface. Int. J. Mol. Sci. 2020, 21, 6593. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Ma, L.; Ji, S.; Yang, W.; Cao, B. Isolation and characterization of Bacillus subtilis strain BY-3, a thermophilic and efficient cellulase-producing bacterium on untreated plant biomass. Lett. Appl. Microbiol. 2014, 59, 306–312. [Google Scholar] [CrossRef] [PubMed]
- Mokale Kognou, A.L.; Chio, C.; Khatiwada, J.R.; Shrestha, S.; Chen, X.; Han, S.; Li, H.; Jiang, Z.H.; Xu, C.C.; Qin, W. Characterization of cellulose-degrading bacteria isolated from soil and the optimization of their culture conditions for cellulase production. Appl. Biochem. Biotechnol. 2022, 194, 5060–5082. [Google Scholar] [CrossRef]
- Ekperigin, M.M. Preliminary studies of cellulase production by Acinetobacter anitratus and Branhamella sp. Afr. J. Biotechnol. 2007, 6, 28–33. [Google Scholar] [CrossRef]
- Asha, B.M.; Sakthivel, N. Production, purification and characterization of a new cellulase from Bacillus subtilis that exhibit halophilic, alkalophilic and solvent-tolerant properties. Ann. Microbiol. 2014, 64, 1839–1848. [Google Scholar] [CrossRef]
- Ahmed, A.; Bibi, A. Fungal Cellulase; Production and Applications: Minireview. LIFE Int. J. Health Life-Sci. 2018, 4, 19–36. [Google Scholar] [CrossRef]
- Pirzadah, T.; Garg, S.; Singh, J.; Vyas, A.; Kumar, M.; Gaur, N.; Bala, M.; Rehman, R.; Varma, A.; Kumar, V.; et al. Characterization of Actinomycetes and Trichoderma spp. for cellulase production utilizing crude substrates by response surface methodology. SpringerPlus. 2014, 3, 622. [Google Scholar] [CrossRef]
- Yadav, S.; Dubey, S.K. Cellulose degradation potential of Paenibacillus lautus strain BHU3 and its whole genome sequence. Bioresour. Technol. 2018, 262, 124–131. [Google Scholar] [CrossRef]
- Dehghanikhah, F.; Shakarami, J.; Asoodeh, A. Purification and Biochemical Characterization of Alkalophilic Cellulase from the Symbiotic Bacillus subtilis BC1 of the Leopard Moth, Zeuzera pyrina (L.) (Lepidoptera: Cossidae). Curr. Microbiol. 2020, 77, 1254–1261. [Google Scholar] [CrossRef]
- Atta, N.N.; El-Baz, A.A.; Said, N.; Abdel Daiem, M.M. Anaerobic co-digestion of wastewater activated sludge and rice straw in batch and semi continuous modes. J. Fundam Renew. Energy Appl. 2016, 6, 1–7. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate’s ID | d (mm) | D (mm) | Cellulolytic Index (D/d) |
|---|---|---|---|
| DC-5 | 0.40 | 1.30 | 3.250 |
| DC-10 | 0.20 | 0.50 | 2.500 |
| DC-11 | 0.50 | 1.75 | 3.500 |
| DC-20 | 0.50 | 1.30 | 2.670 |
| DC-25 | 0.30 | 0.70 | 2.300 |
| DC-28 | 0.40 | 1.20 | 3.000 |
| Physiological and Biochemical Test | Results |
|---|---|
| Hydrogen sulfide | + |
| Gelatin liquefaction | + |
| Contact enzyme assay | + |
| Methyl red | − |
| V-P | + |
| Indole test | + |
| Starch hydrolysis | + |
| Salt tolerance 3% NaCl test | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Zhang, M.; Zhang, Y.; Xu, X.; Zhao, Y.; Jiang, X.; Zhang, R.; Gui, Z. Characterization of Cellulose-Degrading Bacteria Isolated from Silkworm Excrement and Optimization of Its Cellulase Production. Polymers 2023, 15, 4142. https://doi.org/10.3390/polym15204142
Li H, Zhang M, Zhang Y, Xu X, Zhao Y, Jiang X, Zhang R, Gui Z. Characterization of Cellulose-Degrading Bacteria Isolated from Silkworm Excrement and Optimization of Its Cellulase Production. Polymers. 2023; 15(20):4142. https://doi.org/10.3390/polym15204142
Chicago/Turabian StyleLi, Hao, Minqi Zhang, Yuanhao Zhang, Xueming Xu, Ying Zhao, Xueping Jiang, Ran Zhang, and Zhongzheng Gui. 2023. "Characterization of Cellulose-Degrading Bacteria Isolated from Silkworm Excrement and Optimization of Its Cellulase Production" Polymers 15, no. 20: 4142. https://doi.org/10.3390/polym15204142