
Anti-Melanogenesis and Anti-Photoaging Effects of the Sulfated Polysaccharides Isolated from the Brown Seaweed Padina boryana
 ,
,  , , , , , and
, , , , , and {kind=link}
{kind=link}
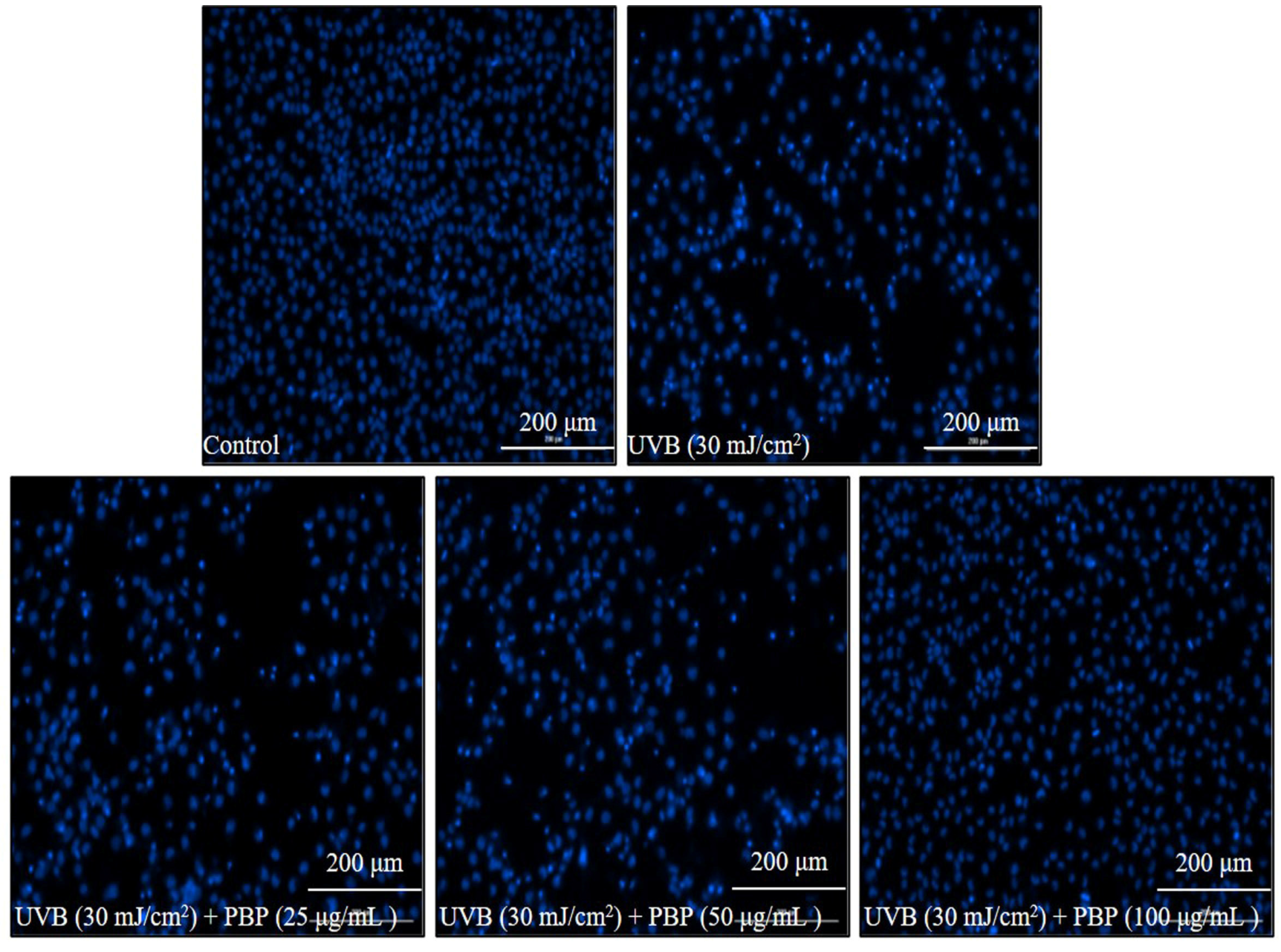{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Preparation and Characterization of Sulfated Polysaccharides from P. boryana
2.3. Evaluation of Tyrosinase and Collagenase Inhibitory Effect of PBP
2.4. Maintenance of Cell Lines
2.5. Determination of Cytotoxicity of PBP
2.6. Determination of the Anti-Melanogenesis Activity of PBP
2.7. Determination of the Photoprotective Effect of PBP on HaCaT Cells
2.8. Determination of the Photoprotective Effect of PBP in HDF Cells
2.9. Statistical Analysis
3. Results and Discussion
3.1. Anti-Melanogenesis Effect of PBP
3.2. Protective Effect of PBP on Photodamage Stimulated by UVB
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gu, Y.; Han, J.; Jiang, C.; Zhang, Y. Biomarkers, oxidative stress and autophagy in skin aging. Ageing Res. Rev. 2020, 59, 101036. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Qu, L.; Wan, S.; Li, Y.; Fan, D. Ginsenoside Rk1 Prevents UVB Irradiation-Mediated Oxidative Stress, Inflammatory Response, and Collagen Degradation via the PI3K/AKT/NF-κB Pathway In Vitro and In Vivo. J. Agric. Food Chem. 2022, 70, 15804–15817. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.-A.; Huang, Y.-T.; Hsiao, P.-F.; Chiu, L.-Y.; Chern, S.-R.; Wu, N.-L. Critical roles of irradiance in the regulation of UVB-induced inflammasome activation and skin inflammation in human skin keratinocytes. J. Photochem. Photobiol. B Biol. 2022, 226, 112373. [Google Scholar] [CrossRef]
- Zheng, X.; Feng, M.; Wan, J.; Shi, Y.; Xie, X.; Pan, W.; Hu, B.; Wang, Y.; Wen, H.; Wang, K.; et al. Anti-damage effect of theaflavin-3′-gallate from black tea on UVB-irradiated HaCaT cells by photoprotection and maintaining cell homeostasis. J. Photochem. Photobiol. B Biol. 2021, 224, 112304. [Google Scholar] [CrossRef]
- Cadet, J.; Douki, T. Formation of UV-induced DNA damage contributing to skin cancer development. Photochem. Photobiol. Sci. 2018, 17, 1816–1841. [Google Scholar] [CrossRef]
- Slominski, A.T.; Hardeland, R.; Zmijewski, M.A.; Slominski, R.M.; Reiter, R.J.; Paus, R. Melatonin: A Cutaneous Perspective on its Production, Metabolism, and Functions. J. Investig. Dermatol. 2018, 138, 490–499. [Google Scholar] [CrossRef]
- Emanuelli, M.; Sartini, D.; Molinelli, E.; Campagna, R.; Pozzi, V.; Salvolini, E.; Simonetti, O.; Campanati, A.; Offidani, A. The Double-Edged Sword of Oxidative Stress in Skin Damage and Melanoma: From Physiopathology to Therapeutical Approaches. Antioxidants 2022, 11, 612. [Google Scholar] [CrossRef]
- Wlaschek, M.; Maity, P.; Makrantonaki, E.; Scharffetter-Kochanek, K. Connective Tissue and Fibroblast Senescence in Skin Aging. J. Investig. Dermatol. 2021, 141, 985–992. [Google Scholar] [CrossRef] [PubMed]
- Fitsiou, E.; Pulido, T.; Campisi, J.; Alimirah, F.; Demaria, M. Cellular Senescence and the Senescence-Associated Secretory Phenotype as Drivers of Skin Photoaging. J. Investig. Dermatol. 2021, 141, 1119–1126. [Google Scholar] [CrossRef]
- Bae, J.-E.; Choi, H.; Shin, D.W.; Na, H.-W.; Park, N.Y.; Kim, J.B.; Jo, D.S.; Cho, M.J.; Lyu, J.H.; Chang, J.H.; et al. Fine particulate matter (PM2.5) inhibits ciliogenesis by increasing SPRR3 expression via c-Jun activation in RPE cells and skin keratinocytes. Sci. Rep. 2019, 9, 3994. [Google Scholar] [CrossRef]
- Wang, L.; Jayawardena, T.U.; Yang, H.-W.; Lee, H.-G.; Jeon, Y.-J. The Potential of Sulfated Polysaccharides Isolated from the Brown Seaweed Ecklonia maxima in Cosmetics: Antioxidant, Anti-melanogenesis, and Photoprotective Activities. Antioxidants 2020, 9, 724. [Google Scholar] [CrossRef]
- Wang, L.; Oh, J.-Y.; Lee, W.; Jeon, Y.-J. Fucoidan isolated from Hizikia fusiforme suppresses ultraviolet B-induced photodamage by down-regulating the expressions of matrix metalloproteinases and pro-inflammatory cytokines via inhibiting NF-κB, AP-1, and MAPK signaling pathways. Int. J. Biol. Macromol. 2021, 166, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, M.; Yang, K.; Tian, J. 6,6′-Bieckol induces apoptosis and suppresses TGF-β-induced epithelial-mesenchymal transition in non-small lung cancer cells. Chin. Herb. Med. 2022, 14, 254–262. [Google Scholar] [CrossRef]
- Park, E.; Yu, H.; Lim, J.-H.; Hee Choi, J.; Park, K.-J.; Lee, J. Seaweed metabolomics: A review on its nutrients, bioactive compounds and changes in climate change. Food Res. Int. 2023, 163, 112221. [Google Scholar] [CrossRef] [PubMed]
- Dinesh Kumar, S.; Satish, L.; Dhanya, N.; Malar Vizhi, J.; Nadukkattu Nayagi, N.; Gopala Krishnan, S.; Ganesan, M. Tank cultivation of edible seaweeds: An overview of the Indian perspective for opportunities and challenges. Biomass Convers. Biorefinery 2023, 1–11. [Google Scholar] [CrossRef]
- Akrong, M.O.; Anning, A.K.; Addico, G.N.D.; Hogarh, J.N.; Adu-Gyamfi, A.; de Graft-Johnson, K.A.A.; Ale, M.; Ampofo, J.A.; Meyer, A.S. Variations in seaweed-associated and planktonic bacterial communities along the coast of Ghana. Mar. Biol. Res. 2023, 19, 219–233. [Google Scholar] [CrossRef]
- Xie, C.; Lee, Z.J.; Ye, S.; Barrow, C.J.; Dunshea, F.R.; Suleria, H.A.R. A Review on Seaweeds and Seaweed-Derived Polysaccharides: Nutrition, Chemistry, Bioactivities, and Applications. Food Rev. Int. 2023, 1–36. [Google Scholar] [CrossRef]
- Aly, S.H.; Elissawy, A.M.; Salah, D.; Alfuhaid, N.A.; Zyaan, O.H.; Mohamed, H.I.; Singab, A.N.B.; Farag, S.M. Phytochemical Investigation of Three Cystoseira Species and Their Larvicidal Activity Supported with In Silico Studies. Mar. Drugs 2023, 21, 117. [Google Scholar] [CrossRef]
- Mohd Fauziee, N.A.; Chang, L.S.; Wan Mustapha, W.A.; Md Nor, A.R.; Lim, S.J. Functional polysaccharides of fucoidan, laminaran and alginate from Malaysian brown seaweeds (Sargassum polycystum, Turbinaria ornata and Padina boryana). Int. J. Biol. Macromol. 2021, 167, 1135–1145. [Google Scholar] [CrossRef] [PubMed]
- Khongthong, S.; Theapparat, Y.; Roekngam, N.; Tantisuwanno, C.; Otto, M.; Piewngam, P. Characterization and immunomodulatory activity of sulfated galactan from the red seaweed Gracilaria fisheri. Int. J. Biol. Macromol. 2021, 189, 705–714. [Google Scholar] [CrossRef]
- Rodrigues-Souza, I.; Pessatti, J.B.K.; da Silva, L.R.; de Lima Bellan, D.; de Souza, I.R.; Cestari, M.M.; de Assis, H.C.S.; Rocha, H.A.O.; Simas, F.F.; da Silva Trindade, E.; et al. Protective potential of sulfated polysaccharides from tropical seaweeds against alkylating- and oxidizing-induced genotoxicity. Int. J. Biol. Macromol. 2022, 211, 524–534. [Google Scholar] [CrossRef]
- Deniaud-Bouët, E.; Hardouin, K.; Potin, P.; Kloareg, B.; Hervé, C. A review about brown algal cell walls and fucose-containing sulfated polysaccharides: Cell wall context, biomedical properties and key research challenges. Carbohydr. Polym. 2017, 175, 395–408. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Li, Q.; Xu, X.; Li, G.; Tian, C.; Zhang, T. Molecular mechanisms of anti-cancer bioactivities of seaweed polysaccharides. Chin. Herb. Med. 2022, 14, 528–534. [Google Scholar] [CrossRef] [PubMed]
- Santinon, C.; Ochi, D.; Beppu, M.M.; Vieira, M.G.A. Chemical modifications in the structure of seaweed polysaccharides as a viable antimicrobial application: A current overview and future perspectives. Algal Res. 2022, 66, 102796. [Google Scholar] [CrossRef]
- Vishchuk, O.S.; Ermakova, S.P.; Zvyagintseva, T.N. Sulfated polysaccharides from brown seaweeds Saccharina japonica and Undaria pinnatifida: Isolation, structural characteristics, and antitumor activity. Carbohydr. Res. 2011, 346, 2769–2776. [Google Scholar] [CrossRef]
- Pereira, L.; Critchley, A.T. The COVID 19 novel coronavirus pandemic 2020: Seaweeds to the rescue? Why does substantial, supporting research about the antiviral properties of seaweed polysaccharides seem to go unrecognized by the pharmaceutical community in these desperate times? J. Appl. Phycol. 2020, 32, 1875–1877. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Jia, J.; Jiang, P.; Zheng, W.; Li, X.; Song, S.; Ai, C. Polysaccharides from edible brown seaweed Undaria pinnatifida are effective against high-fat diet-induced obesity in mice through the modulation of intestinal microecology. Food Funct. 2022, 13, 2581–2593. [Google Scholar] [CrossRef]
- Arunkumar, K.; Raja, R.; Kumar, V.B.S.; Joseph, A.; Shilpa, T.; Carvalho, I.S. Antioxidant and cytotoxic activities of sulfated polysaccharides from five different edible seaweeds. J. Food Meas. Charact. 2021, 15, 567–576. [Google Scholar] [CrossRef]
- Corino, C.; Di Giancamillo, A.; Modina, S.C.; Rossi, R. Prebiotic Effects of Seaweed Polysaccharides in Pigs. Animals 2021, 11, 1573. [Google Scholar] [CrossRef]
- Koh, H.S.A.; Lu, J.; Zhou, W. Structural Dependence of Sulfated Polysaccharide for Diabetes Management: Fucoidan from Undaria pinnatifida Inhibiting α-Glucosidase More Strongly than α-Amylase and Amyloglucosidase. Front. Pharmacol. 2020, 11, 831. [Google Scholar] [CrossRef]
- Wang, L.; Oh, J.-Y.; Kim, Y.-S.; Lee, H.-G.; Lee, J.-S.; Jeon, Y.-J. Anti-Photoaging and Anti-Melanogenesis Effects of Fucoidan Isolated from Hizikia fusiforme and Its Underlying Mechanisms. Mar. Drugs 2020, 18, 427. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Barbosa, J.; Palhares, L.C.G.F.; Silva, C.H.F.; Sabry, D.A.; Chavante, S.F.; Rocha, H.A.O. In Vitro Antitumor Potential of Sulfated Polysaccharides from Seaweed Caulerpa cupressoides var. flabellata. Mar. Biotechnol. 2021, 23, 77–89. [Google Scholar] [CrossRef]
- Chaves Filho, G.P.; Lima, M.E.G.B.; Rocha, H.A.d.O.; Moreira, S.M.G. Role of sulfated polysaccharides from seaweeds in bone regeneration: A systematic review. Carbohydr. Polym. 2022, 284, 119204. [Google Scholar] [CrossRef] [PubMed]
- Jayawardena, T.U.; Wang, L.; Sanjeewa, K.K.A.; Kang, S.I.; Lee, J.-S.; Jeon, Y.-J. Antioxidant Potential of Sulfated Polysaccharides from Padina boryana; Protective Effect against Oxidative Stress in In Vitro and In Vivo Zebrafish Model. Mar. Drugs 2020, 18, 212. [Google Scholar] [CrossRef] [PubMed]
- Wen, S.; An, R.; Li, D.; Cao, J.; Li, Z.; Zhang, W.; Chen, R.; Li, Q.; Lai, X.; Sun, L.; et al. Tea and Citrus maxima complex induces apoptosis of human liver cancer cells via PI3K/AKT/mTOR pathway in vitro. Chin. Herb. Med. 2022, 14, 449–458. [Google Scholar] [CrossRef]
- Guo, W.; Zhang, Q.; Du, Y.; Guo, J.; Zhao, T.; Bai, L.; An, X. Immunomodulatory activity of polysaccharides from Brassica rapa by activating Akt/NF-κB signaling. Chin. Herb. Med. 2022, 14, 90–96. [Google Scholar] [CrossRef]
- Zhou, S.; Zeng, H.; Huang, J.; Lei, L.; Tong, X.; Li, S.; Zhou, Y.; Guo, H.; Khan, M.; Luo, L.; et al. Epigenetic regulation of melanogenesis. Ageing Res. Rev. 2021, 69, 101349. [Google Scholar] [CrossRef]
- Yousef, S.; Papadimitropoulos, M.; Faris, M.; Hasan, H.; Hossain, A.; Colman, I.; Manuel, D.; Wells, G.A. Melanin levels in relation to vitamin D among first-generation immigrants from different ethnic groups and origins: A comparative national Canadian cross-sectional study. Front. Med. 2023, 9, 992554. [Google Scholar] [CrossRef]
- Yang, X.; Tang, C.; Zhao, Q.; Jia, Y.; Qin, Y.; Zhang, J. Melanin: A promising source of functional food ingredient. J. Funct. Foods 2023, 105, 105574. [Google Scholar] [CrossRef]
- Bellei, B.; Picardo, M. Premature cell senescence in human skin: Dual face in chronic acquired pigmentary disorders. Ageing Res. Rev. 2020, 57, 100981. [Google Scholar] [CrossRef]
- Jayawardena, T.U.; Sanjeewa, K.K.A.; Kim, H.-S.; Lee, H.G.; Wang, L.; Lee, D.-S.; Jeon, Y.-J. Padina boryana, a brown alga from the Maldives: Inhibition of α-MSH-stimulated melanogenesis via the activation of ERK in B16F10 cells. Fish. Aquat. Sci. 2020, 23, 8. [Google Scholar] [CrossRef]
- Wang, L.; Oh, J.Y.; Jayawardena, T.U.; Jeon, Y.-J.; Ryu, B. Anti-inflammatory and anti-melanogenesis activities of sulfated polysaccharides isolated from Hizikia fusiforme: Short communication. Int. J. Biol. Macromol. 2020, 142, 545–550. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, G.; D’Amico, A.G.; Amenta, A.; Saccone, S.; Federico, C.; Reibaldi, M.; Russo, A.; Bonfiglio, V.; Avitabile, T.; Longo, A.; et al. Protective effect of PACAP against ultraviolet B radiation-induced human corneal endothelial cell injury. Neuropeptides 2020, 79, 101978. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Huang, F.; McClements, D.J.; Xie, B.; Sun, Z.; Deng, Q. Oligomeric Procyanidin Nanoliposomes Prevent Melanogenesis and UV Radiation-Induced Skin Epithelial Cell (HFF-1) Damage. Molecules 2020, 25, 1458. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Jayawardena, T.U.; Kim, Y.-S.; Wang, K.; Fu, X.; Ahn, G.; Cha, S.-H.; Kim, J.G.; Lee, J.S.; Jeon, Y.-J. Anti-Melanogenesis and Anti-Photoaging Effects of the Sulfated Polysaccharides Isolated from the Brown Seaweed Padina boryana. Polymers 2023, 15, 3382. https://doi.org/10.3390/polym15163382
Wang L, Jayawardena TU, Kim Y-S, Wang K, Fu X, Ahn G, Cha S-H, Kim JG, Lee JS, Jeon Y-J. Anti-Melanogenesis and Anti-Photoaging Effects of the Sulfated Polysaccharides Isolated from the Brown Seaweed Padina boryana. Polymers. 2023; 15(16):3382. https://doi.org/10.3390/polym15163382
Chicago/Turabian StyleWang, Lei, Thilina U. Jayawardena, Young-Sang Kim, Kaiqiang Wang, Xiaoting Fu, Ginnae Ahn, Seon-Heui Cha, Jeong Gyun Kim, Jung Suck Lee, and You-Jin Jeon. 2023. "Anti-Melanogenesis and Anti-Photoaging Effects of the Sulfated Polysaccharides Isolated from the Brown Seaweed Padina boryana" Polymers 15, no. 16: 3382. https://doi.org/10.3390/polym15163382