
Improving the Autofluorescence of Lophira alata Woody Cells via the Removal of Extractives
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
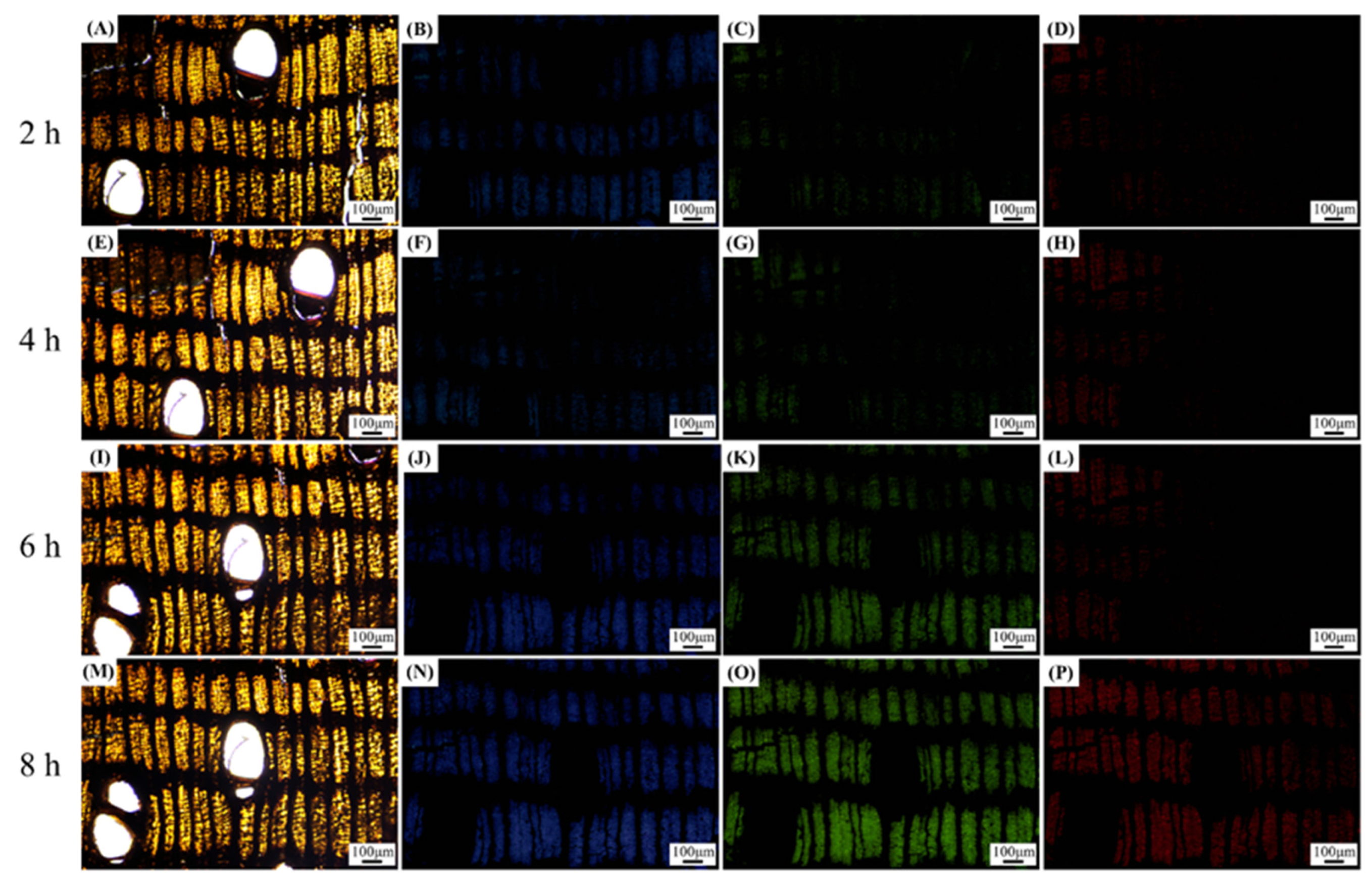
2.2. Autofluorescence Experiment
2.3. Diffuse Reflection Spectra and UV–Vis Absorption Spectra
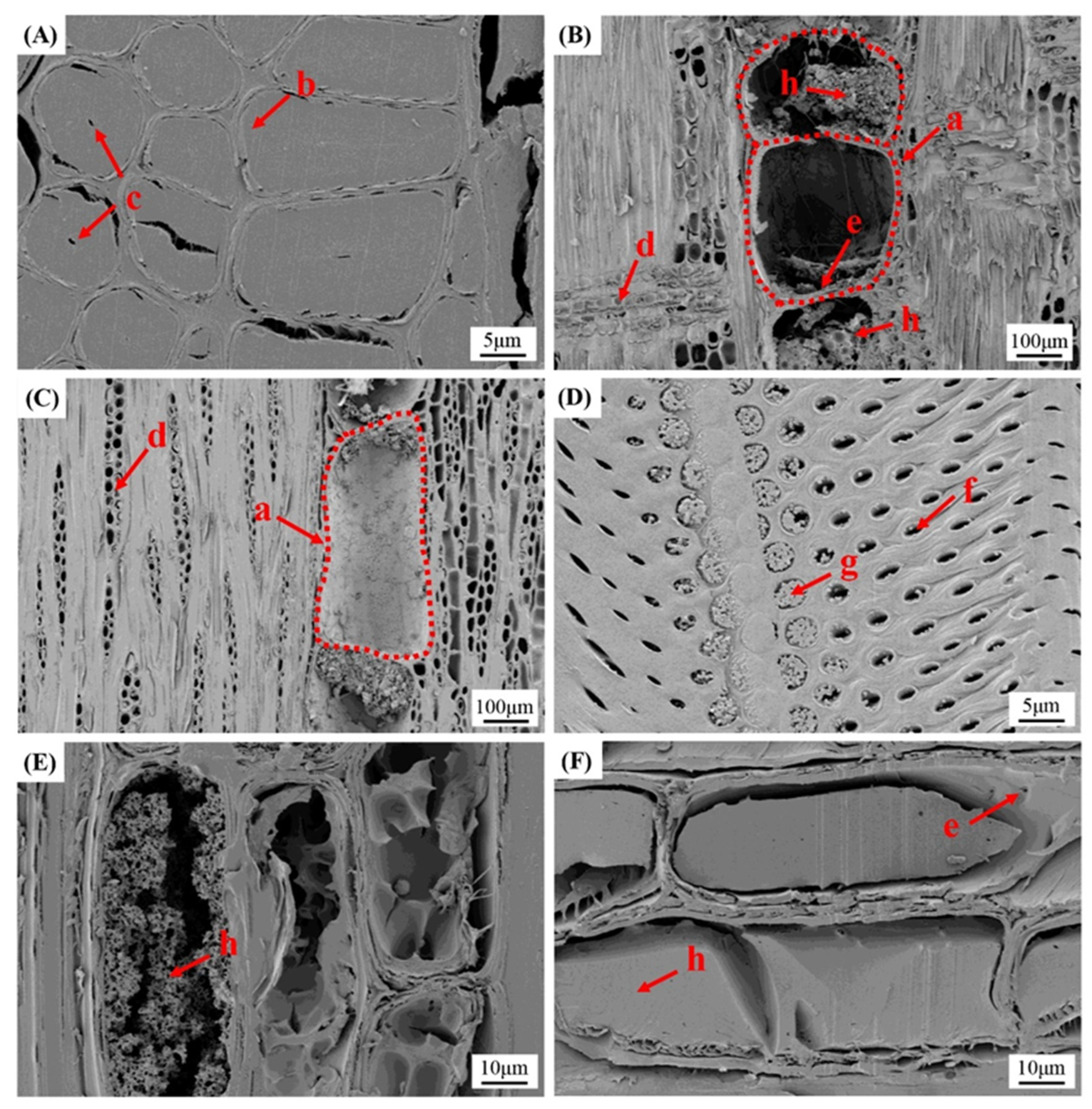
2.4. Macrostructural Observation
2.5. Microstructural and SEM Observation
3. Results
3.1. Autofluorescence Experiment
3.2. Diffuse Reflection Spectra and UV–Vis Absorption Spectra
3.3. Macroscopic Observation
3.4. Microscopic Observation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roshchina, V.V. Vital Autofluorescence: Application to the Study of Plant Living Cells. Int. J. Spectrosc. 2012, 2012, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Angyalossy-Alfonso, V.; Miller, R.B. Wood anatomy of the Brazilian species of Swartizia and considerations within the tribe Swartzieae. IAWA J. 2002, 23, 359–390. [Google Scholar] [CrossRef]
- Decou, R.; Serk, H.; Menard, D.; Pesquet, E. Analysis of Lignin Composition and Distribution Using Fluorescence Laser Confocal Microspectroscopy. Methods Mol. Biol. 2017, 1544, 233–247. [Google Scholar]
- Moran-Mirabal, J.M. The study of cell wall structure and cellulose–cellulase interactions through fluorescence microscopy. Cellulose 2013, 20, 2291–2309. [Google Scholar] [CrossRef]
- Djikanović, D.; Kalauzi, A.; Radotić, K.; Lapierre, C.; Jeremić, M. Deconvolution of lignin fluorescence spectra: A contribution to the comparative structural studies of lignins. Russ. J. Phys. Chem. A 2007, 81, 1425–1428. [Google Scholar]
- Donaldson, L.A.; Radotic, K. Fluorescence lifetime imaging of lignin autofluorescence in normal and compression wood. J. Microsc. 2013, 251, 178–187. [Google Scholar] [CrossRef]
- Donaldson, L. Softwood and Hardwood Lignin Fluorescence Spectra of Wood Cell Walls in Different Mounting Media. IAWA J. 2013, 34, 3–19. [Google Scholar] [CrossRef]
- Pegg, T.J.; Gladish, D.K.; Baker, R.L. Algae to angiosperms: Autofluorescence for rapid visualization of plant anatomy among diverse taxa. Appl. Plant. Sci. 2021, 9, e11437. [Google Scholar] [CrossRef] [PubMed]
- Godoy-Veiga, M.; Slotta, F.; Alecio, P.C.; Ceccantini, G.; Buckeridge, M.S.; Locosselli, G.M. Improved tree-ring visualization using autofluorescence. Dendrochronologia 2019, 55, 33–42. [Google Scholar] [CrossRef]
- Palanti, S.; Feci, E.; Anichini, M. Comparison between four tropical wood species for their resistance to marine borers (Teredo spp. and Limnoria spp.) in the Strait of Messina. Int. Biodeter. Biodegr. 2015, 104, 472–476. [Google Scholar] [CrossRef]
- Edoun, F.; Tchuente, B.R.T.; Dibacto, R.E.K.; Mouafo, H.T.; Tchuenchieu, A.K.; Ndzana, A.A.; Medoua, G. Phytochemical screening and antioxidant potential of aqueous extracts of Millettia laurenti, Lophira alata and Milicia excelsa, commonly used in the Cameroonian pharmacopoeia. Eur. J. Med. Plants 2020, 31, 11–23. [Google Scholar]
- Mouafo, H.T.; Tchuenchieu, A.D.K.; Nguedjo, M.W.; Edoun, F.L.E.; Tchuente, B.R.T.; Medoua, G.N. In vitro antimicrobial activity of Millettia laurentii De Wild and Lophira alata Banks ex CF Gaertn on selected foodborne pathogens associated to gastroenteritis. Heliyon 2021, 7, e06830. [Google Scholar] [CrossRef] [PubMed]
- Ajiboye, T.O.; Yakubu, M.T.; Oladiji, A.T. Cytotoxic, antimutagenic, and antioxidant activities of methanolic extract and chalcone dimers (lophirones B and C) derived from Lophira alata (Van Tiegh. Ex Keay) stem bark. Evid. Based Complement. Altern. Med. 2014, 19, 20–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tchinda, J.-B.S.; Ndikontar, M.K.; Belinga, A.D.F.; Mounguengui, S.; Njankouo, J.M.; Durmaçay, S.; Gerardin, P. Inhibition of fungi with wood extractives and natural durability of five Cameroonian wood species. Ind. Crop. Prod. 2018, 123, 183–191. [Google Scholar] [CrossRef]
- Kirker, G.; Blodgett, A.; Arango, R.; Lebow, P.; Clausen, C. The role of extractives in naturally durable wood species. Int. Biodeter. Biodegr. 2013, 82, 53–58. [Google Scholar] [CrossRef]
- Gasson, P.; Miller, R.; Stekel, D.J.; Whinder, F.; Ziemińska, K. Wood identification of Dalbergia nigra (CITES Appendix I) using quantitative wood anatomy, principal components analysis and naïve Bayes classification. Ann. Bot. 2010, 105, 45–56. [Google Scholar]
- Thomas, J.; Ingerfeld, M.; Nair, H.; Chauhan, S.S.; Collings, D.A. Pontamine fast scarlet 4B: A new fluorescent dye for visualising cell wall organisation in radiata pine tracheids. Wood Sci. Technol. 2012, 47, 59–75. [Google Scholar]
- Liu, G.; Chen, D.; Liu, R.; Yu, Z.; Jiang, J.; Liu, Y.; Hu, J.; Chang, S. Antifouling Wood Matrix with Natural Water Transfer and Microreaction Channels for Water Treatment. ACS Sustain. Chem. Eng. 2019, 7, 6782–6791. [Google Scholar] [CrossRef]
- ISO 3129; International Organization for Standardization. Wood: Sampling Methods and General Requirements for Physical and Mechanical Testing of Small Clear Wood Specimens. ISO: Geneva, Switzerland, 2019.
- Han, J.; Pan, F.; Molokeev, M.S.; Dai, J.; Peng, M.; Zhou, W.; Wang, J. Redefinition of Crystal Structure and Bi3+ Yellow Luminescence with Strong Near-ultraviolet Excitation in La3BWO9:Bi3+ Phosphor for White Light-emitting Diodes. ACS Appl. Mater. Interfaces. 2018, 10, 13660–13668. [Google Scholar] [CrossRef] [PubMed]
- Kishino, M.; Nakano, T. Artificial weathering of tropical woods. Part 1: Changes in wettability. Holzforschung 2004, 58, 552–557. [Google Scholar]
- Tolvaj, L.; Mitsui, K.; Varga, D. Validity limits of Kubelka-Munk theory for DRIFT spectra of photodegraded solid wood. Wood Sci. Technol. 2010, 45, 135–146. [Google Scholar] [CrossRef]
- Deng, Y.; Feng, X.; Zhou, M.; Qian, Y.; Yu, H.; Qiu, X. Investigation of aggregation and assembly of alkali lignin using iodine as a probe. Biomacromolecules 2011, 12, 1116–1125. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Jiang, G.; Xiao, M.; Zhou, J.; Yi, Z. Effects of heat treatment on water-soluble extractives and color changes of merbau heartwood. J. Wood Sci. 2012, 58, 465–469. [Google Scholar] [CrossRef]
- Lange, W.; Faix, O. Lignin-Polyphenol Interaction in Azobe (Lophira alata) heartwood. A Study on Milled Wood Lignin (MWL) and Klason Residues. Holzforschung 1999, 53, 519–524. [Google Scholar] [CrossRef]
- De Micco, V.; Balzano, A.; Wheeler, E.A.; Baas, P. Tyloses and gums: A review of structure, function and occurrence of vessel occlusions. Iawa J. 2016, 37, 186–205. [Google Scholar]
- Ding, Q.; Zhang, P. Revealing the autofluorescence properties of nanocellulose isolated from different raw materials by different methods. IOP Conf. Ser. Earth Environ. Sci. 2021, 639, 012037. [Google Scholar] [CrossRef]
- Li, Y.; Ren, J.; Sun, R.; Wang, X. Fluorescent Lignin Carbon Dots for Reversible Responses to High-Valence Metal Ions and Its Bioapplications. J. Biomed. Nanotechnol. 2018, 14, 1543–1555. [Google Scholar] [PubMed]
- Xie, F.; Gong, S.; Zhang, W.; Wu, J.; Wang, Z. Potential of lignin from Canna edulis ker residue in the inhibition of alpha-d-glucosidase: Kinetics and interaction mechanism merging with docking simulation. Int. J. Biol. Macromol. 2017, 95, 592–602. [Google Scholar]
- Simovski, C.R.; Mollaei, M.S.M.; Voroshilov, P.M. Fluorescence quenching by plasmonic nanoantennas. Phys. Rev. B 2020, 101, 245421. [Google Scholar] [CrossRef]
- Lin, T.; Liu, X.; Lou, Z.; Hou, Y.; Teng, F. Intermolecular-charge-transfer-induced fluorescence quenching in protic solvent. J. Mol. Struct. 2016, 1123, 49–54. [Google Scholar] [CrossRef]
- Li, W.; Chen, Z.; Yu, H.; Li, J.; Liu, S. Wood-Derived Carbon Materials and Light-Emitting Materials. Adv. Mater. 2021, 33, 2000596. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, L.A.; Grace, J.; Downes, G.M. Within-Tree Variation in Anatomical Properties of Compression Wood in Radiata Pine. IAWA J. 2004, 25, 253–271. [Google Scholar] [CrossRef] [Green Version]
- Xue, Y.; Qiu, X.; Ouyang, X. Insights into the effect of aggregation on lignin fluorescence and its application for microstructure analysis. Int. J. Biol. Macromol. 2020, 154, 981–988. [Google Scholar] [CrossRef]
- Shen, Q.; Xue, Y.; Zhang, Y.; Li, T.; Yang, T.; Li, S. Effect of microstructure-scale features on lignin fluorescence for preparation of high fluorescence efficiency lignin-based nanomaterials. Int. J. Biol. Macromol. 2022, 202, 520–528. [Google Scholar] [CrossRef] [PubMed]
- Iwata, M.; Gutierrez, A.; Marques, G.; Sabat, G.; Kersten, P.J.; Cullen, D.; Bhatnagar, J.M.; Yadav, J.; Lipzen, A.; Yoshinaga, Y.; et al. Omics analyses and biochemical study of Phlebiopsis gigantea elucidate its degradation strategy of wood extractives. Sci. Rep. 2021, 11, 12528. [Google Scholar] [PubMed]
- Xiong, F.; Han, Y.; Li, G.; Qin, T.; Wang, S.; Chu, F. Synthesis and characterization of renewable woody nanoparticles fluorescently labeled by pyrene. Ind. Crop. Prod. 2016, 83, 663–669. [Google Scholar] [CrossRef]
- Badiya, P.K.; Ramamurthy, S.S. Smartphone Plasmonics for Doxycycline Detection with Silver-Lignin Bio-spacer at Attomolar Sensitivity. Plasmonics 2017, 13, 955–960. [Google Scholar] [CrossRef]
- Ma, X.J.; Cao, S.L.; Yang, X.F.; Lin, L.; Chen, L.H.; Huang, L.L. Lignin removal and benzene-alcohol extraction effects on lignin measurements of the hydrothermal pretreated bamboo substrate. Bioresour. Technol. 2014, 151, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Costas, C.; Palanti, S.; Charpentier, J.-P.; Sanromán, M.Á.; Moldes, D. A Sustainable Treatment for Wood Preservation: Enzymatic Grafting of Wood Extractives. ACS Sustain. Chem. Eng. 2017, 5, 7557–7567. [Google Scholar]
- Guo, Y.; Cao, F.; Qiu, P.; Wang, Z. Studies of the effect of halide ions on the fluorescence of quinine sulfate. Luminescence 2019, 34, 450–455. [Google Scholar] [CrossRef]
- Musha, Y.; Goring, D. Distribution of syringyl and guaiacyl moieties in hardwoods as indicated by ultraviolet microscopy. Wood Sci. Technol. 1975, 9, 45–58. [Google Scholar] [CrossRef]
- Chang, T.-C.; Chang, H.-T.; Wu, C.-L.; Chang, S.-T. Influences of extractives on the photodegradation of wood. Polym. Degrad. Stabil. 2010, 95, 516–521. [Google Scholar] [CrossRef]
- Panda, M.; Chandel, T.I.; Kamil, M.; Khan, R.H. Fluorescence quenching of chloroquine by Cu2+ in micelles. J. Mol. Liq. 2020, 306, 112763. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measurements | Average | Maximum | Minimum | Standard Deviation | Variation Coefficient/% |
|---|---|---|---|---|---|
| Double wall thickness/µm | 14.6 | 16.8 | 12.3 | 1.0 | 7.2 |
| Cell lumen diameter/µm | 2.4 | 2.9 | 1.9 | 0.3 | 12.3 |
| Wall-to-lumen ratio | 5.9 | 7.6 | 5.1 | 0.7 | 11.7 |
| Cavity-to-diameter ratio | 0.14 | 0.16 | 0.12 | 0.01 | 9.63 |
| Measurements | Average | Maximum | Minimum | Standard Deviation | Variation Coefficient/% |
|---|---|---|---|---|---|
| Vessels/% | 10.2 | 13.3 | 8.6 | 0.02 | 15.9 |
| Fibers/% | 58.2 | 61.7 | 55.8 | 0.02 | 3.3 |
| Longitudinal parenchyma/% | 14.4 | 16.2 | 10.9 | 0.01 | 10.2 |
| Rays/% | 17.2 | 18.7 | 15.7 | 0.01 | 6.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Z.; Xu, D.; Hu, J.; Chang, S.; Liu, G.; Huang, Q.; Han, J.; Li, T.; Liu, Y.; Wang, X. Improving the Autofluorescence of Lophira alata Woody Cells via the Removal of Extractives. Polymers 2023, 15, 3269. https://doi.org/10.3390/polym15153269
Yu Z, Xu D, Hu J, Chang S, Liu G, Huang Q, Han J, Li T, Liu Y, Wang X. Improving the Autofluorescence of Lophira alata Woody Cells via the Removal of Extractives. Polymers. 2023; 15(15):3269. https://doi.org/10.3390/polym15153269
Chicago/Turabian StyleYu, Zhaoyang, Dongnian Xu, Jinbo Hu, Shanshan Chang, Gonggang Liu, Qiongtao Huang, Jin Han, Ting Li, Yuan Liu, and Xiaodong (Alice) Wang. 2023. "Improving the Autofluorescence of Lophira alata Woody Cells via the Removal of Extractives" Polymers 15, no. 15: 3269. https://doi.org/10.3390/polym15153269