
Alpha-1,4-transglycosylation Activity of GH57 Glycogen Branching Enzymes Is Higher in the Absence of a Flexible Loop with a Conserved Tyrosine Residue

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Enzyme Production and Purification
2.3. Enzyme Activity with Iodine Assay
2.4. Enzyme Reactions and Analysis with Reducing End Assay
2.5. Chain Length Distribution with Anion Exchange Chromatography
2.6. Statistical Analysis
3. Results and Discussion
3.1. Definition of Activity for Glycogen Branching Enzymes
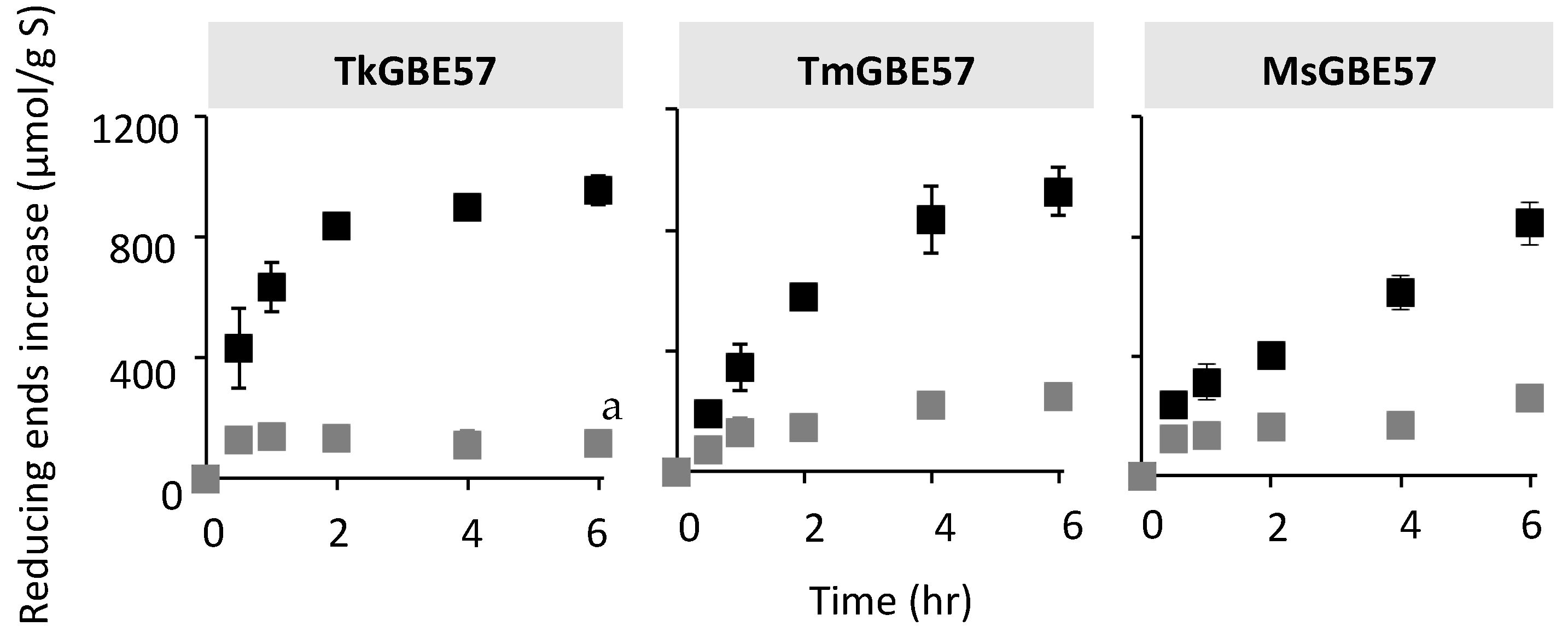
3.2. Activity of GH57 Glycogen Branching Enzymes on Linear Maltodextrin
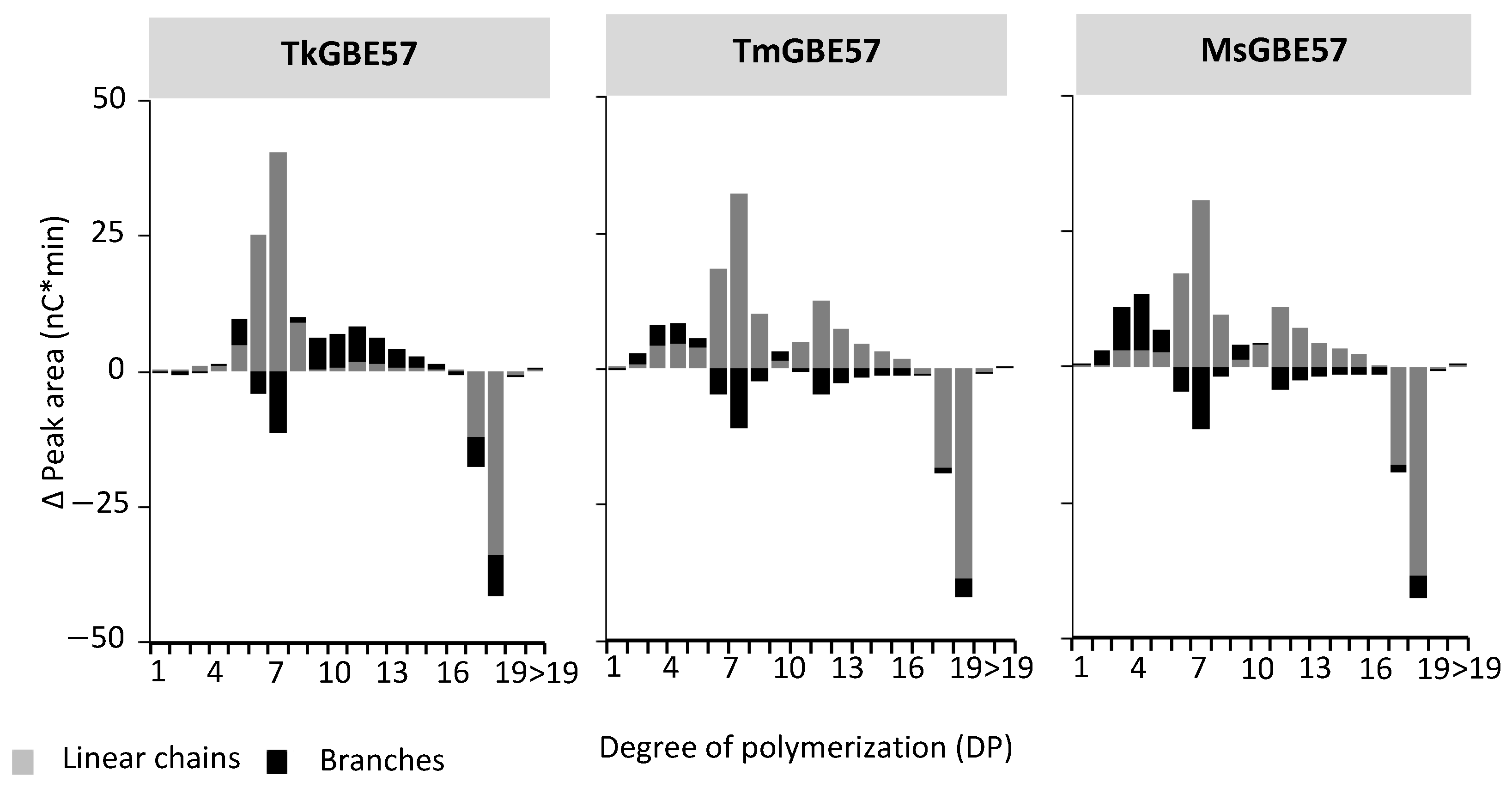
3.3. Chain Length Distribution of Modified Maltodextrin
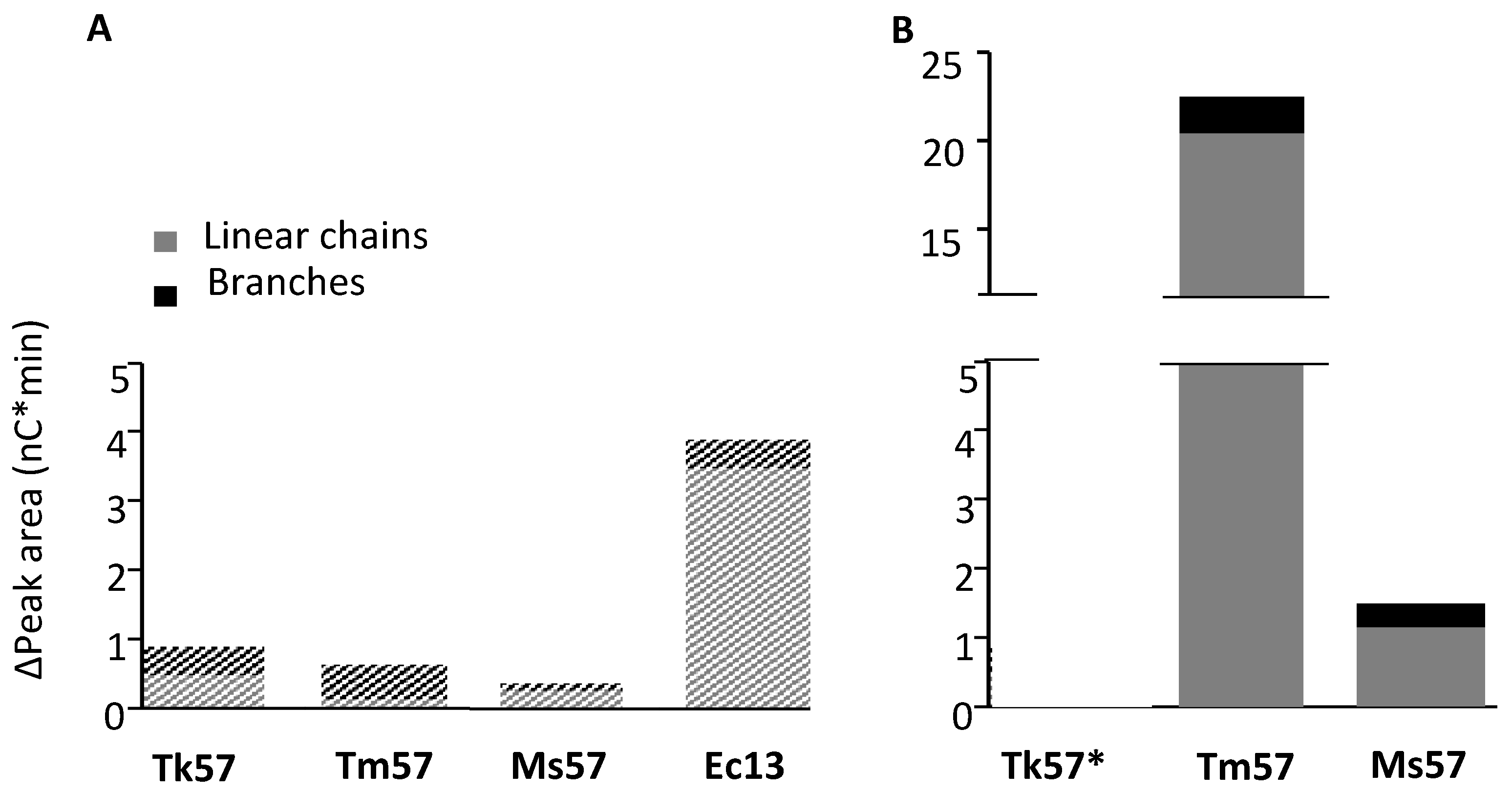
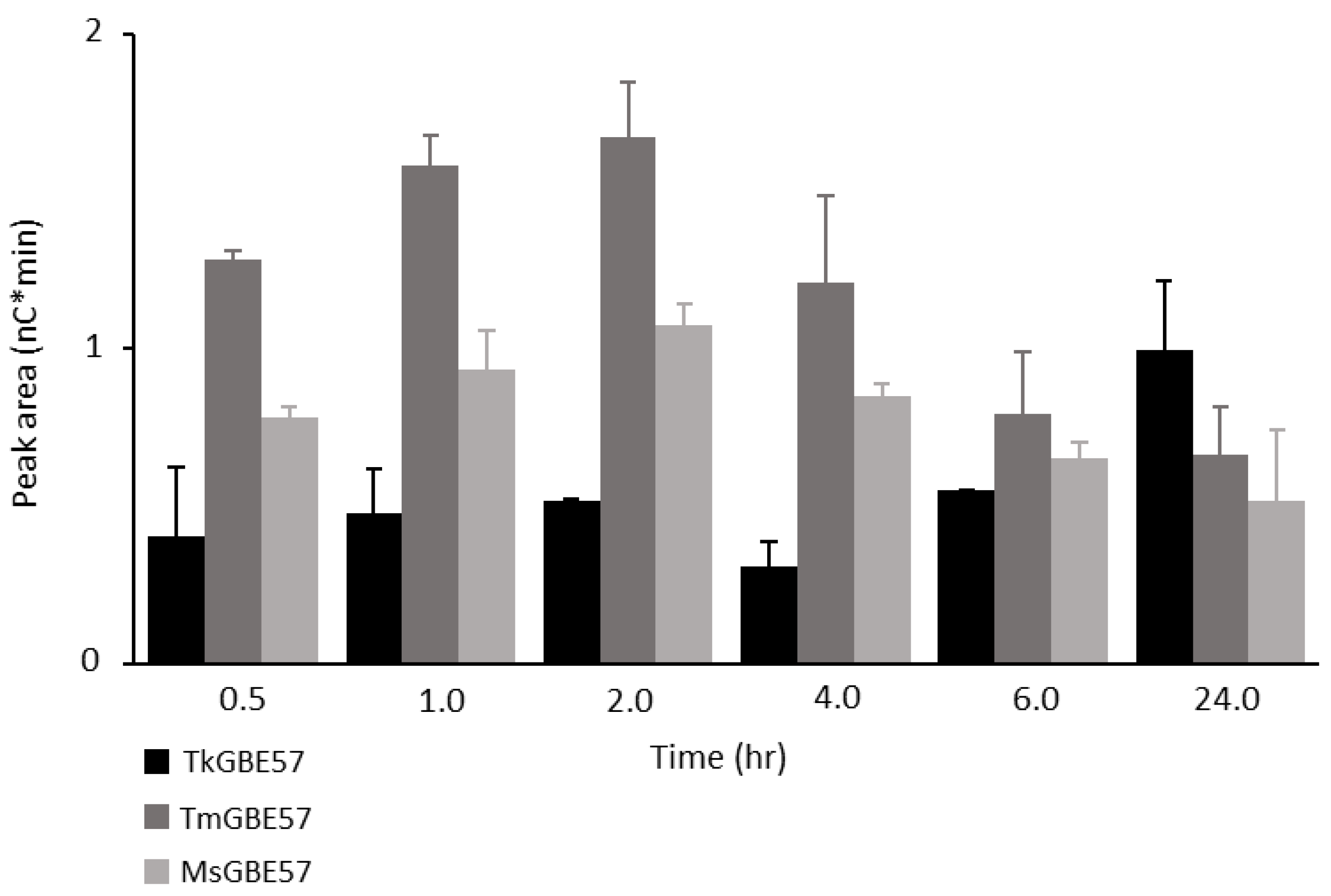
3.4. Alpha-1,4-transferase Activity of GH57 Glycogen Branching Enzymes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Ball, S.G.; Morell, M.K. From Bacterial Glycogen to Starch: Understanding the Biogenesis of the Plant Starch Granule. Annu. Rev. Plant Biol. 2003, 54, 207–233. [Google Scholar] [CrossRef]
- Roach, P.J.; Depaoli-Roach, A.A.; Hurley, T.D.; Tagliabracci, V.S. Glycogen and its metabolism: Some new developments and old themes. Biochem. J. 2012, 441, 763–787. [Google Scholar] [CrossRef] [Green Version]
- Cifuente, J.O.; Comino, N.; Trastoy, B.; D’Angelo, C.; Guerin, M.E. Structural basis of glycogen metabolism in bacteria. Biochem. J. 2019, 476, 2059–2092. [Google Scholar] [CrossRef] [PubMed]
- D’hulst, C.; Mérida, Á. The priming of storage glucan synthesis from bacteria to plants: Current knowledge and new developments. New Phytol. 2010, 188, 13–21. [Google Scholar] [CrossRef]
- Rashid, A.M.; Batey, S.F.; Syson, K.; Koliwer-Brandl, H.; Miah, F.; Barclay, J.E.; Findlay, K.C.; Nartowski, K.P.; Khimyak, Y.Z.; Kalscheuer, R. Assembly of α-Glucan by GlgE and GlgB in Mycobacteria and Streptomycetes. Biochemistry 2016, 55, 3270–3284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Wang, M.; Wise, M.J.; Liu, Q.; Yang, T.; Zhu, Z.; Li, C.; Tan, X.; Tang, D.; Wang, W. Recent progress in the structure of glycogen serving as a durable energy reserve in bacteria. World J. Microbiol. Biotechnol. 2020, 36, 14. [Google Scholar] [CrossRef] [PubMed]
- Takata, H.; Kajiura, H.; Furuyashiki, T.; Kakutani, R.; Kuriki, T. Fine structural properteis of natural and synthetic glycogens. Carbohydr. Res. 2009, 344, 654–659. [Google Scholar] [CrossRef]
- Yoo, S.-H.; Keppel, C.; Spalding, M.; Jane, J.-L. Effects of growth condition on the structure of glycogen produced in cyanobacterium Synechocystis sp. PCC6803. Int. J. Biol. Macromol. 2007, 40, 498–504. [Google Scholar] [CrossRef]
- Hayashi, M.; Suzuki, R.; Colleoni, C.; Ball, S.G.; Fujita, N.; Suzuki, E. Bound Substrate in the Structure of Cyanobacterial Branching Enzyme Supports a New Mechanistic Model. J. Biol. Chem. 2017, 292, 5465–5475. [Google Scholar] [CrossRef] [Green Version]
- Gavgani, H.N.; Fawaz, R.; Ehyaei, N.; Walls, D.; Pawlowski, K.; Fulgos, R.; Park, S.; Assar, Z.; Ghanbarpour, A.; Geiger, J.H. A structural explanation for the mechanism and specificity of plant branching enzymes I and IIb. J. Biol. Chem. 2022, 298, 101395. [Google Scholar] [CrossRef]
- Wang, L.; Wise, M.J. Glycogen with short average chain length enhances bacterial durability. Sci. Nat. 2011, 98, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Binderup, K.; Mikkelsen, R.; Preiss, J. Truncation of the Amino Terminus of Branching Enzyme Changes Its Chain Transfer Pattern. Arch. Biochem. Biophys. 2002, 397, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, J.; Chaen, K.; Vu, N.T.; Akasaka, T.; Shimada, H.; Nakashima, T.; Nishi, A.; Satoh, H.; Omori, T.; Kakuta, Y.; et al. Crystal structure of the branching enzyme I (BEI) from Oryza sativa L. with implications for catalysis and substrate binding. Glycobiology 2011, 21, 1108–1116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abad, M.C.; Binderup, K.; Rios-Steiner, J.; Arni, R.K.; Preiss, J.; Geiger, J.H. The X-ray Crystallographic Structure ofEscherichia coli Branching Enzyme. J. Biol. Chem. 2002, 277, 42164–42170. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, E.; Suzuki, R. Distribution of glucan-branching enzymes among prokaryotes. Cell. Mol. Life Sci. 2016, 73, 2643–2660. [Google Scholar] [CrossRef]
- Moller, M.S.; Henriksen, A.; Svensson, B. Structure and function of α-glucan debranching enzymes. Cell. Mol. Life Sci. 2016, 73, 2619–2641. [Google Scholar] [CrossRef] [Green Version]
- Conchou, L.; Martin, J.; Gonçalves, I.R.; Galisson, F.; Violot, S.; Guillière, F.; Aghajari, N.; Ballut, L. The Candida glabrata glycogen branching enzyme structure reveals unique features of branching enzymes of the Saccharomycetaceae phylum. Glycobiology 2022, 32, 343–355. [Google Scholar] [CrossRef]
- Suzuki, R.; Suzuki, E. Structure and function of branching enzymes in eukaryotes. Trends Glycosci. 2020, 32, E21–E30. [Google Scholar] [CrossRef] [Green Version]
- Drula, E.; Garron, M.-L.; Dogan, S.; Lombard, V.; Henrissat, B.; Terrapon, N. The carbohydrate-active enzyme database: Functions and literature. Nucleic Acids Res. 2021, 50, D571–D577. [Google Scholar] [CrossRef]
- Stam, M.R.; Danchin, E.G.; Rancurel, C.; Coutinho, P.M.; Henrissat, B. Dividing the large glycoside hydrolase family 13 into subfamilies: Towards improved functional annotations of -amylase-related proteins. Protein Eng. Des. Sel. 2006, 19, 555–562. [Google Scholar] [CrossRef] [Green Version]
- Palomo, M.; Pijning, T.; Booiman, T.; Dobrochowska, J.M.; van der Vlist, J.; Kralj, S.; Planas, A.; Loos, K.; Kamerling, J.P.; Dijkstra, B.W.; et al. Thermus thermophilus glycoside hydrolase family 57 branching enzyme crystal structure, mechanism of action, and products formed. J. Biol. Chem. 2011, 286, 3520–3530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Na, S.; Park, M.; Jo, I.; Cha, J.; Ha, N.-C. Structural basis for the transglycosylase activity of a GH57-type glycogen branching enzyme from Pyrococcus horikoshii. Biochem. Biophys. Res. Commun. 2017, 484, 850–856. [Google Scholar] [CrossRef]
- Santos, C.R.; Tonoli, C.C.; Trindade, D.M.; Betzel, C.; Takata, H.; Kuriki, T.; Kanai, T.; Imanaka, T.; Arni, R.K.; Murakami, M.T. Structural basis for branching-enzyme activity of glycoside hydrolase family 57: Structure and stability studies of a novel branching enzyme from the hyperthermophilic archaeon Thermococcus Kodakaraensis KOD1. Proteins 2011, 79, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Janeček, S.; Gabriško, M. Remarkable evolutionary relatedness among the enzymes and proteins from the α-amylase family. Cell. Mol. Life Sci. 2016, 73, 2707–2725. [Google Scholar] [CrossRef] [PubMed]
- Gaenssle, A.L.O.; Bax, H.H.M.; Jurak, E. GH13 Glycogen branching enzymes can adapt the substrate chain length to-wards their preferences via α-1, 4-transglycosylation. Enzyme Microb. Technol. 2021, 150, 109882. [Google Scholar] [CrossRef]
- Kuriki, T.; Imanaka, T. The concept of the α-amylase family: Structural similarity and common catalytic mechanism. J. Biosci. Bioeng. 1999, 87, 557–565. [Google Scholar] [CrossRef]
- Sarian, F.D.; Janeček, Š.; Pijning, T.; Nurachman, Z.; Radjasa, O.K.; Dijkhuizen, L.; Natalia, D.; van der Maarel, M.J.E.C. A new group of glycoside hydrolase family 13 α-amylases with an aberrant catalytic triad. Sci. Rep. 2017, 7, 44230. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.; Ryu, S.-I.; Lee, S.-B.; Moon, T.-W. Purification and characterization of branching specificity of a novel extra-cellular amylolytic enzyme from marine hyperthermophilic Rhodothermus marinus. J. Microbiol. Biotechnol. 2008, 18, 457–464. [Google Scholar]
- Zhang, X.; Leemhuis, H.; van der Maarel, M.J.E.C. Synthesis of highly branched α-glucans with different structures using GH13 and GH57 glycogen branching enzymes. Carbohydr. Polym. 2019, 216, 231–237. [Google Scholar] [CrossRef]
- Roussel, X.; Lancelon-Pin, C.; Viksø-Nielsen, A.; Rolland-Sabaté, A.; Grimaud, F.; Potocki-Véronèse, G.; Buléon, A.; Putaux, J.-L.; D’Hulst, C. Characterization of substrate and product specificity of the purified recombinant glycogen branching enzyme of Rhodothermus obamensis. Biochim. Et Biophys. Acta (BBA) Gen. Subj. 2013, 1830, 2167–2177. [Google Scholar] [CrossRef]
- Tran, P.L.; An, Y.; Jeong, G.-Y.; Ban, S.-Y.; Nguyen, P.C.; Woo, E.; You, S.; Park, J.-T. One-step synthesis of glycogen-type polysaccharides from maltooctaose and its structural characteristics. Carbohydr. Polym. 2022, 284, 119175. [Google Scholar] [CrossRef]
- Dickmanns, A.; Ballschmiter, M.; Liebl, W.; Ficner, R. Structure of the novel α-amylase AmyC from Thermotoga maritima. Acta Crystallogr. D 2006, 62, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Xiang, G.; Leemhuis, H.; van der Maarel, M.J.E.C. Structural elements determining the transglycosylating activity of glycoside hydrolase family 57 glycogen branching enzymes. Proteins 2022, 90, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Storms, R.; Tsang, A. A quantitative starch–iodine method for measuring alpha-amylase and glucoamylase activities. Anal. Biochem. 2006, 351, 146–148. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Leemhuis, H.; Janeček, Š.; Martinovičová, M.; Pijning, T.; van der Maarel, M.J.E.C. Identification of Thermotoga maritima MSB8 GH57 α-amylase AmyC as a glycogen-branching enzyme with high hydrolytic activity. Appl. Microbiol. Biotechnol. 2019, 103, 6141–6151. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Leemhuis, H.; van der Maarel, M.J.E.C. Characterization of the GH13 and GH57 glycogen branching enzymes from Petrotoga mobilis SJ95 and potential role in glycogen biosynthesis. PLoS ONE 2019, 14, e0219844. [Google Scholar] [CrossRef] [Green Version]
- Gaenssle, A.L.O.; van der Maarel, M.J.E.C.; Jurak, E. The influence of amylose content on the modification of starches by glycogen branching enzymes. Food Chem. 2022, 393, 133294. [Google Scholar] [CrossRef]
- Gaenssle, A.L.; van der Maarel, M.J.; Jurak, E. Reliability factor for identification of amylolytic enzyme activity in the optimized starch-iodine assay. Anal. Biochem. 2020, 597, 113696. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TmGBE57 | TkGBE57 | MsGBE57 | EcGBE13 | |
|---|---|---|---|---|
| Non-branching activity [mUNB/mg E] | 4.4 ± 1.0 | 13.0 ± 5.6 | 5.2 ± 0.3 | 120.1 ± 0.9 a |
| Branching activity [mUB/mg E] | 5.0 ± 0.6 | 92.4 ± 9.5 a | 5.6 ± 0.2 | 286.9 ± 42.5 b |
| Ratio B:NB * | 1.2 ± 0.2 | 8.1 ± 2.3 a | 1.1 ± 0.0 | 2.4 ± 0.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bax, H.H.M.; van der Maarel, M.J.E.C.; Jurak, E. Alpha-1,4-transglycosylation Activity of GH57 Glycogen Branching Enzymes Is Higher in the Absence of a Flexible Loop with a Conserved Tyrosine Residue. Polymers 2023, 15, 2777. https://doi.org/10.3390/polym15132777
Bax HHM, van der Maarel MJEC, Jurak E. Alpha-1,4-transglycosylation Activity of GH57 Glycogen Branching Enzymes Is Higher in the Absence of a Flexible Loop with a Conserved Tyrosine Residue. Polymers. 2023; 15(13):2777. https://doi.org/10.3390/polym15132777
Chicago/Turabian StyleBax, Hilda Hubertha Maria, Marc Jos Elise Cornelis van der Maarel, and Edita Jurak. 2023. "Alpha-1,4-transglycosylation Activity of GH57 Glycogen Branching Enzymes Is Higher in the Absence of a Flexible Loop with a Conserved Tyrosine Residue" Polymers 15, no. 13: 2777. https://doi.org/10.3390/polym15132777