
Inhibition of Carrageenan-Induced Acute Inflammation in Mice by the Microgramma vacciniifolia Frond Lectin (MvFL)

 , , and
, , and 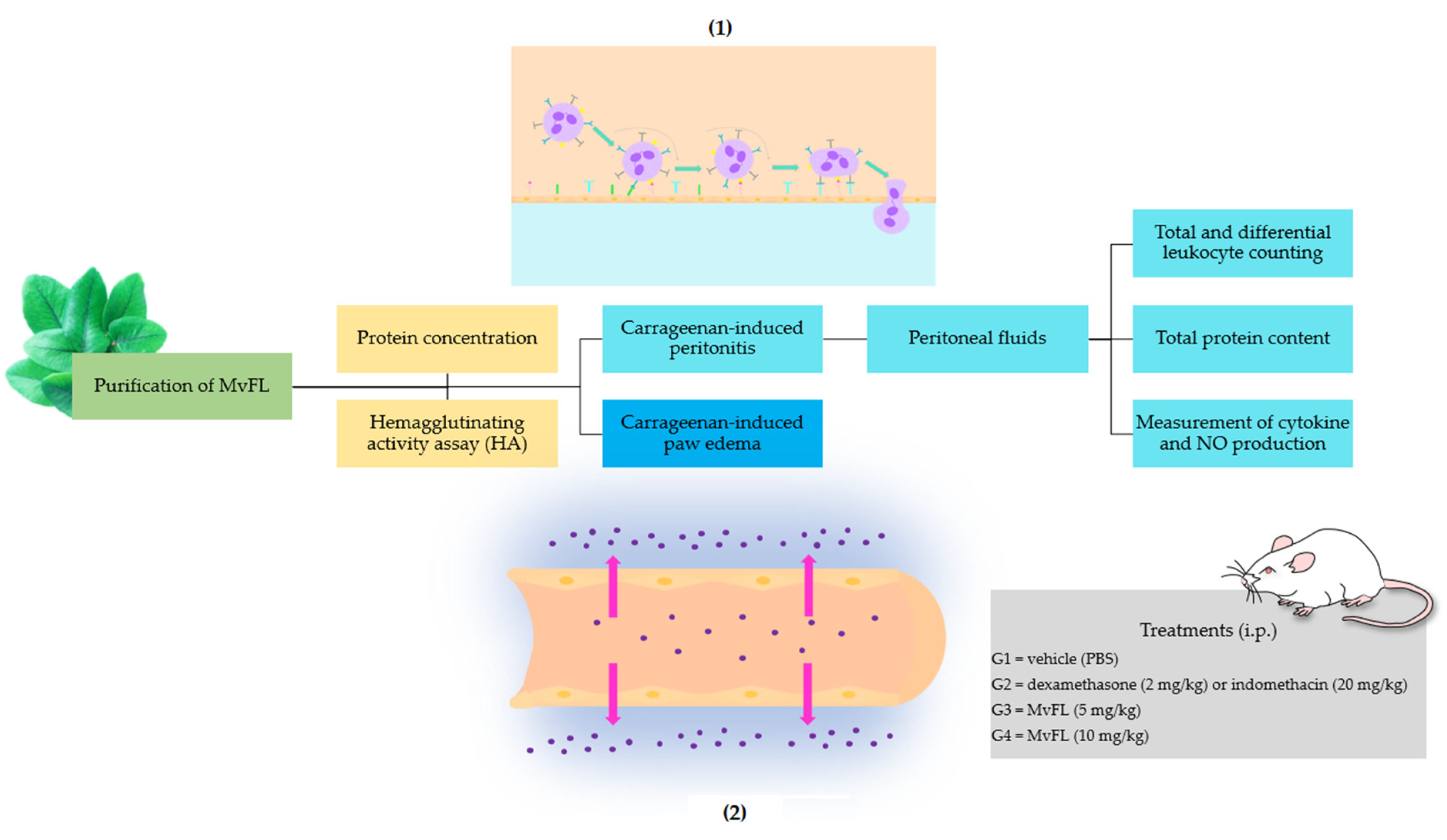{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Purification of MvFL
2.2. Protein Concentration
2.3. Hemagglutinating Activity Assay (HA)
2.4. Animals
2.5. Carrageenan-Induced Peritonitis
2.5.1. Induction of Inflammation and Treatments
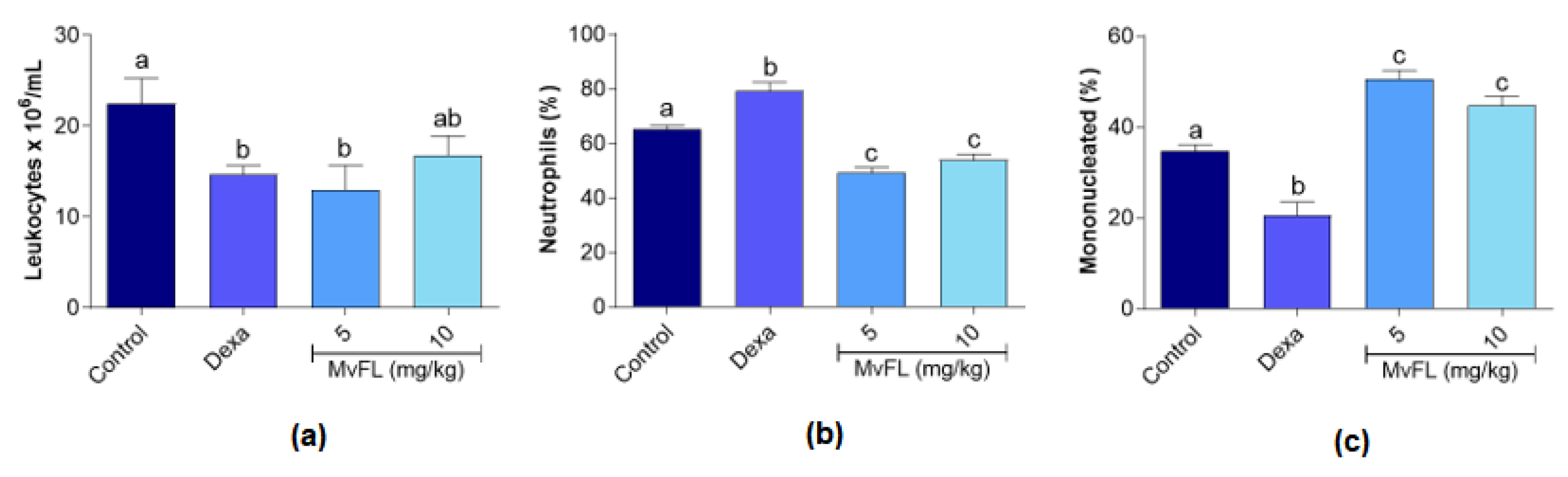
2.5.2. Leukocyte Counting
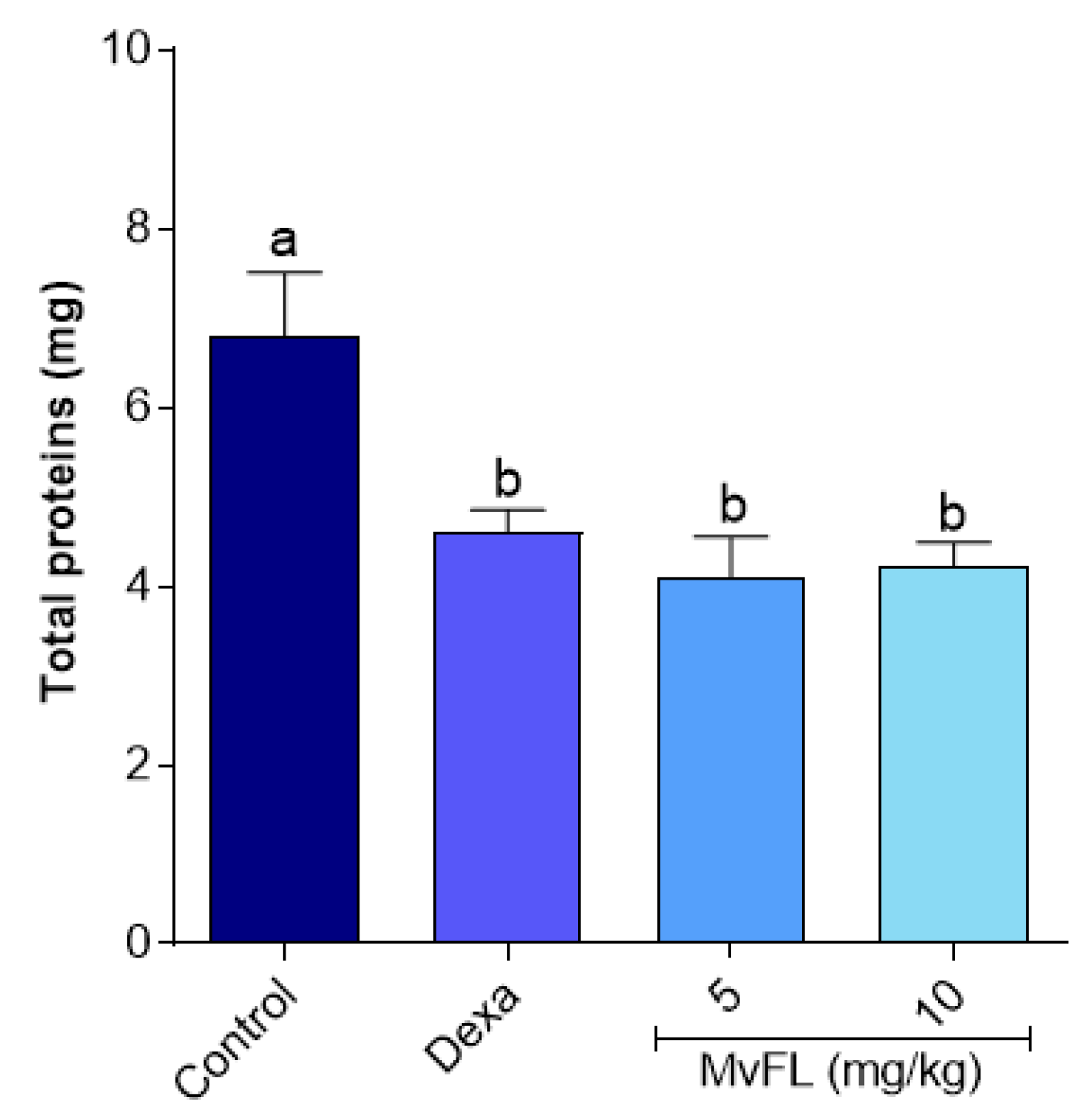
2.5.3. Total Protein Content
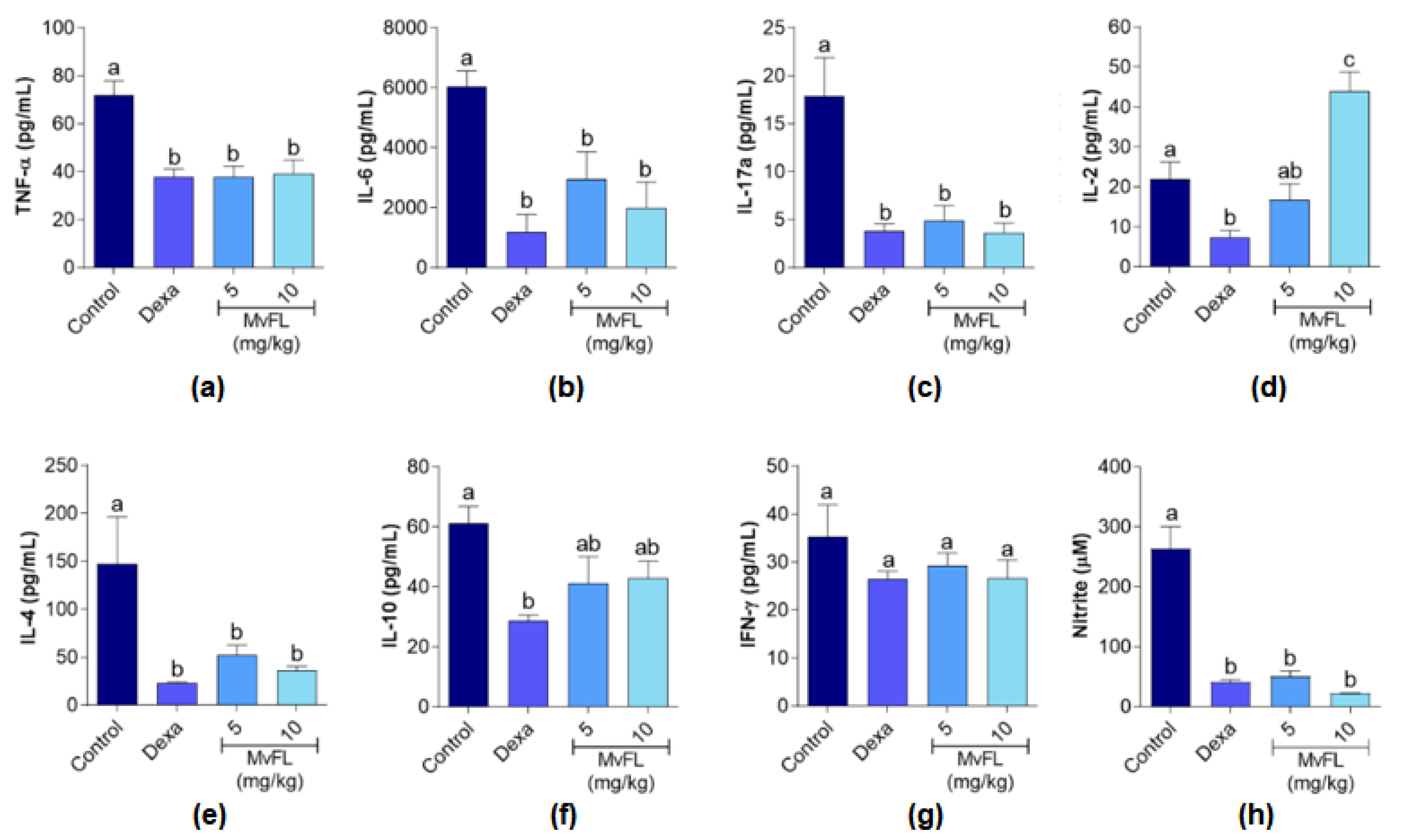
2.5.4. Measurement of Cytokine and NO Production
2.6. Carrageenan-Induced Paw Edema Assay
2.7. Statistical Analysis
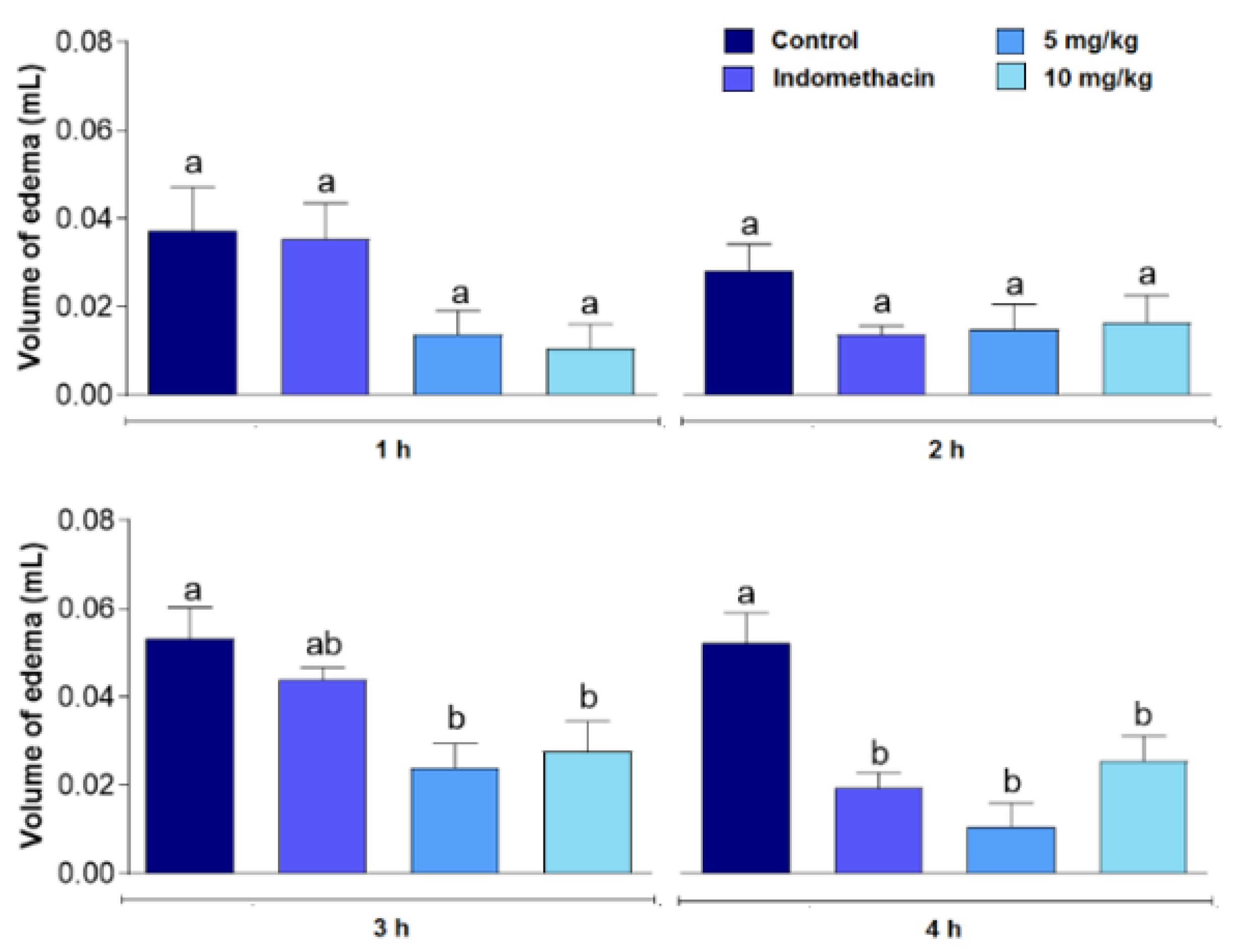
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Meizlish, M.L.; Franklin, R.A.; Zhou, X.; Medzhitov, R. Tissue Homeostasis and Inflammation. Annu. Rev. Immunol. 2021, 39, 557–581. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Shi, G. Editorial: Regulation of Inflammation in Chronic Disease. Front. Immunol. 2019, 10, 737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdulkhaleq, L.A.; Assi, M.A.; Abdullah, R.; Zamri-Saad, M.; Taufiq-Yap, Y.H.; Hezmee, M.N.M. The crucial roles of inflammatory mediators in inflammation: A review. Vet. World 2018, 11, 627–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouyahya, A.; Guaouguaou, F.E.; Omari, N.E.; Menyiy, N.E.; Balahbib, A.; El-Shazly, M.; Bakri, Y. Anti-inflammatory and analgesic properties of Moroccan medicinal plants: Phytochemistry, in vitro and in vivo investigations, mechanism insights, clinical evidences and perspectives. J. Pharm. Anal. 2022, 12, 35–57. [Google Scholar] [CrossRef]
- Oladokun, O.B.; Omotayo, O.; Omisore, N.; Osuko, O.A. Anti-nociceptive and anti-inflammatory activities of Tetracarpidium conophorum seed lectin. Sci. Afr. 2019, 3, e00073. [Google Scholar] [CrossRef]
- Patil, K.R.; Mahajan, U.B.; Unger, B.S.; Goyal, S.N.; Belemkar, S.; Surana, S.J.; Ojha, S.; Patil, C.R. Animal Models of Inflammation for Screening of Anti-inflammatory Drugs: Implications for the Discovery and Development of Phytopharmaceuticals. Int. J. Mol. Sci. 2019, 20, 4367. [Google Scholar] [CrossRef] [Green Version]
- Coelho, L.C.B.B.; Silva, P.M.S.; Lima, V.L.M.; Pontual, E.V.; Paiva, P.M.G.; Napoleão, T.H.; Correia, M.T.S. Lectins, interconnecting proteins with biotechnological/pharmacological and therapeutic applications. Evid. Based Complem. Altern. Med. 2017, 2017, 1594074. [Google Scholar] [CrossRef]
- Chettri, D.; Boro, M.; Sarkar, L.; Verma, A.K. Lectins: Biological significance to biotechnological application. Carbohydr. Res. 2021, 506, 108367. [Google Scholar] [CrossRef]
- Mishra, A.; Behura, A.; Mawatwal, S.; Kumar, A.; Naik, L.; Mohanty, S.S.; Manna, D.; Dokania, P.; Mishra, A.; Patra, S.K.; et al. Structure-function and application of plant lectins in disease biology and immunity. Food Chem. Toxicol. 2019, 134, 1108272. [Google Scholar] [CrossRef]
- Patriota, L.L.S.; Brito, J.S.; Ramos, D.B.M.; Procópio, T.F.; Paiva, P.M.G.; Pontual, E.V.; Melo, C.M.L.; Napoleão, T.H. Plant-derived lectins: A review of their status as alternatives to anticancer chemotherapy. In Horizons in Cancer Research, 1st ed.; Watanabe, H.S., Ed.; Nova Science Publishers Inc.: New York, NY, USA, 2019; Volume 73, pp. 171–205. [Google Scholar]
- Gautam, A.K.; Sharma, D.; Sharma, J.; Saini, K.C. Legume lectins: Potential use as a diagnostics and therapeutics against the cancer. Int. J. Biol. Macromol. 2020, 142, 474–483. [Google Scholar] [CrossRef]
- Silva, P.M.; Silva, B.R.; Silva, J.N.O.; Moura, M.C.; Soares, T.; Feitosa, A.P.S.; Brayner, F.A.; Alves, L.C.; Paiva, P.M.G.; Damborg, P.; et al. Punica granatum sarcotesta lectin (PgTeL) has antibacterial activity and synergistic effects with antibiotics against β-lactamase-producing Escherichia coli. Int. J. Biol. Macromol. 2019, 135, 931–939. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.D.F.; Silva, S.P.; Silva, P.M.; Vieira, A.M.; Araújo, L.C.C.; Lima, T.A.; Oliveira, A.P.S.; Carvalho, L.V.N.; Pitta, M.G.R.; Rêgo, M.J.B.M.; et al. Portulaca elatior root contains a trehalose-binding lectin with antibacterial and antifungal activities. Int. J. Biol. Macromol. 2019, 126, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Kreis, W. Plant lectins: Potential antineoplastic and antiviral drugs. Phytomedicine 2015, 22, S3. [Google Scholar] [CrossRef]
- Pires, A.F.; Bezerra, M.M.; Amorim, R.M.F.; Nascimento, F.L.F.; Marinho, M.M.; Moura, R.M.; Silva, M.T.L.; Correia, J.L.A.; Cavada, B.S.; Assreuy, A.M.S.; et al. Lectin purified from Lonchocarpus campestris seeds inhibits inflammatory nociception. Int. J. Biol. Macromol. 2019, 125, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Pires, A.F.; Marques, G.F.O.; Alencar, N.M.N.; Martins, M.G.Q.; Silva, M.T.L.; Nascimento, K.S.; Cavada, B.S.; Assreuy, A.M.S. Inhibitory effect of Lonchocarpus araripensis lectin in rat acute models of inflammation. An. Acad. Bras. Cienc. 2019, 91, e20180991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, B.; Sadaria, D.; Warrier, V.U.; Kirtonia, A.; Kant, R.; Awasthi, A.; Baligar, P.; Pal, J.K.; Yuba, E.; Sethi, G.; et al. Plant lectins and their usage in preparing targeted nanovaccines for cancer immunotherapy. Semin. Cancer Biol. 2020, 80, 87–106. [Google Scholar] [CrossRef]
- Bezerra, G.A.; Viertlmayr, R.; Moura, T.R.; Delatorre, P.; Rocha, B.A.M.; Nascimento, K.S.; Figueiredo, J.G.; Bezerra, I.G.; Teixeira, C.S.; Simões, R.C.; et al. Structural studies of an anti-inflammatory lectin from Canavalia boliviana seeds in complex with dimannosides. PLoS ONE 2014, 9, e97015. [Google Scholar] [CrossRef] [Green Version]
- Lacerda, R.R.; Moreira, I.C.; Nascimento, J.S.J.; Lacerda, A.C.S.; Cabral, N.L.; Lucetti, D.L.; Viana, G.S.B.; Felipe, C.F.B.; Pessoa, H.L.F.; Gadelha, C.A.A.; et al. Lectin isolated from Brazilian seeds of velvet bean (Mucuna pruriens (L.) DC. presents analgesic, anti-inflammatory and antihemolytic action. J. Med. Plants Res. 2015, 9, 231–242. [Google Scholar]
- Girão, D.K.F.B.; Cavada, B.S.; Pires, A.F.; Martins, T.V.; Franco, Á.X.; Morais, C.M.; Nascimento, K.S.; Delatorre, P.; Silva, H.C.; Nagano, C.S.; et al. The galactose-binding lectin isolated from Bauhinia bauhinioides Mart seeds inhibits neutrophil rolling and adhesion via primary cytokines. J. Mol. Recognit. 2015, 28, 285–292. [Google Scholar] [CrossRef]
- Butle, A.; Talmale, S.; Patil, M.B. Potential In vivo Immunomodulatory Effects of the most Active Lectin Isolated from Seeds of Zizyphus eonoplia. J. Clin. Cell. Immunol. 2016, 7, 1–6. [Google Scholar]
- Patriota, L.L.S.; Procópio, T.F.; Brito, J.S.; Sebag, V.; Oliveira, A.P.S.; Soares, A.K.A.; Moreira, L.R.; Lima, T.A.; Soares, T.; Silva, T.D.; et al. Microgramma vacciniifolia (Polypodiaceae) fronds contain a multifunctional lectin with immunomodulatory properties on human cells. Int. J. Biol. Macromol. 2017, 103, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Patriota, L.L.S.; Ramos, D.B.M.; Silva, Y.A.; Santos, A.C.L.A.; Araújo, M.T.M.F.; Brito, J.S.; Torres, D.J.L.; Oliveira, A.M.; Silva, D.C.N.; Lorena, V.M.B.; et al. Microgramma vacciniifolia frond lectin (MvFL) exhibits antitumor activity against sarcoma 180 in mice. Phytomed. Plus 2021, 1, 100013. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin-phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Bing, D.H.; Weyand, J.G.M.; Stavitsky, A.B. Hemagglutination with aldehyde-fixed erythrocytes for assay of antigens and antibodies. Proc. Soc. Exp. Biol. Med. 1967, 124, 1166–1170. [Google Scholar] [CrossRef] [PubMed]
- Lavor, E.M.; Leal, A.E.B.P.; Fernandes, A.W.C.; Ribeiro, F.P.R.A.; Barbosa, J.M.; Silva, M.G.; Teles, R.B.A.; Oliveira, L.F.S.; Silva, J.C.; Rolim, L.A.; et al. Ethanolic extract of the aerial parts of Passiflora cincinnata Mast. (Passifloraceae) reduces nociceptive and inflammatory events in mice. Phytomedicine 2018, 47, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.C.; Araújo, C.S.; Lima-Saraiva, S.R.G.; Oliveira-Junior, R.G.; Diniz, T.C.; Wanderley, C.W.S.; Palheta-Júnior, R.C.; Mendes, R.L.; Guimarães, A.G.; Quintans-Júnior, L.J.; et al. Antinociceptive and anti-inflammatory activities of the ethanolic extract of Annona vepretorum Mart. (Annonaceae) in rodents. BMC Complem. Alter. Med. 2015, 15, 197. [Google Scholar] [CrossRef] [Green Version]
- Melo, M.S.; Guimarães, A.G.; Santana, M.F.; Siqueira, R.S.; Lima, A.C.B.; Dias, A.S.; Santos, M.R.V.; Onofre, A.S.C.; Quintans, J.S.S.; Sousa, D.P.; et al. Anti-inflammatory and redox-protective activities of citronellal. Biol. Res. 2011, 44, 363–368. [Google Scholar] [CrossRef] [Green Version]
- Guimarães, A.G.; Xavier, M.A.; Santana, M.T.; Camargo, E.A.; Santos, C.A.; Brito, F.A.; Barreto, E.O.; Cavalcanti, S.C.; Antoniolli, A.R.; Oliveira, R.C.; et al. Carvacrol attenuates mechanical hypernociception and inflammatory response. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2012, 385, 253–263. [Google Scholar] [CrossRef]
- Ding, A.H.; Nathan, C.F.; Stuehr, D.J. Release of reactive nitrogen intermediates and reactive oxygen intermediates from mouse peritoneal macrophages. Comparison of activating cytokines and evidence for independent production. J. Immunol. 1988, 141, 2407–2412. [Google Scholar]
- Patriota, L.L.S.; Ramos, D.B.M.; Silva, M.G.; Santos, A.C.L.A.; Silva, Y.A.; Marinho, A.O.; Coelho, L.C.B.B.; Paiva, P.M.G.; Pontual, E.V.; Mendes, R.L.; et al. The trypsin inhibitor from Moringa oleifera flowers (MoFTI) inhibits acute inflammation in mice by reducing cytokine and nitric oxide levels. South Afr. J. Bot. 2021, 143, 474–481. [Google Scholar] [CrossRef]
- Campos, J.K.L.; Araíjo, C.S.F.; Araújo, T.F.S.; Santos, A.F.S.; Teixeira, J.A.; Lima, V.L.M.; Coelho, L.C.B.B. Anti-inflammatory and antinociceptive activities of Bauhinia monandra leaf lectin. Biochim. Open 2016, 2, 62–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pires, A.F.; Rodrigues, N.V.; Soares, P.M.; Ribeiro, R.A.; Aragão, K.S.; Marinho, M.M.; Silva, M.T.; Cavada, B.S.; Assreuy, A.M. A novel N-acetyl-glucosamine lectin of Lonchocarpus araripensis attenuates acute cellular inflammation in mice. Inflamm. Res. 2016, 1, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Assreuy, A.M.S.; Shibuya, M.D.; Martins, G.J.; Souza, M.L.P.; Cavada, B.S.; Moreira, R.A.; Oliveira, J.T.A.; Ribeiro, R.A.; Flores, C.A. Anti-inflammatory effect of glucose-mannose binding lectins isolated from Brazilian beans. Med. Inflamm. 1997, 6, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Sud, V.; Abboud, A.; Tohme, S.; Vodovotz, Y.; Simmons, R.L.; Tsung, A. IL-17A—A regulator in acute inflammation: Insights from in vitro, in vivo and in silico studies. Cytokine 2018, 139, 154344. [Google Scholar] [CrossRef]
- Xu, J.-G.; Jia, H.; Chen, S.; Xu, J.; Zhan, Y.; Yu, H.; Wang, W.; Kang, X.; Cui, X.; Feng, Y.; et al. Structural and functional insights into a novel pre-clinical-stage antibody targeting IL-17A for treatment of autoimmune diseases. Int. J. Biol. Macromol. 2022, 202, 529–538. [Google Scholar] [CrossRef]
- Ross, S.H.; Cantrell, D.A. Signaling and function of interleukin-2 in T lymphocytes. Annu. Rev. Immunol. 2018, 36, 411–433. [Google Scholar] [CrossRef]
- Burmeister, A.R.; Marriott, I. The interleukin-10 family of cytokines and their role in the CNS. Front. Cell. Neurosci. 2018, 12, 458. [Google Scholar] [CrossRef] [Green Version]
- Silva, H.C.; Bari, A.U.; Rocha, B.A.M.; Nascimento, K.S.; Ponte, E.L.; Pires, A.F.; Delatorre, P.; Teixeira, E.H.; Debray, H.; Assreuy, A.M.S.; et al. Purification and primary structure of a mannose/glucose-binding lectin from Parkia biglobosa Jacq. seeds with antinociceptive and anti-inflammatory properties. J. Mol. Recognit. 2013, 26, 470–478. [Google Scholar] [CrossRef]
- Napimoga, M.H.; Cavada, B.S.; Alencar, N.M.; Mota, M.L.; Bittencourt, F.S.; Alves-Filho, J.C.; Grespan, R.; Gonçalves, R.B.; Clemente-Napimoga, J.T.; Freitas, A.; et al. Lonchocarpus sericeus lectin decreases leukocyte migration and mechanical hypernociception by inhibiting cytokine and chemokines production. Int. Immunopharmacol. 2007, 7, 824–835. [Google Scholar] [CrossRef]
- Lind, M.; Hayes, A.; Caprnda, M.; Petrovic, D.; Rodrigo, L.; Kruzliak, P.; Zulli, A. Inducible nitric oxide synthase: Good or bad? Biomed. Pharmacother. 2017, 93, 370–375. [Google Scholar] [CrossRef]
- Predonzani, A.; Calì, B.; Agnellini, A.H.R.; Molon, B. Spotlights on immunological effects of reactive nitrogen species: When inflammation says nitric oxide. World J. Exp. Med. 2015, 5, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, V.; Abbas, A.K.; Aster, J.C. Robbins Basic Pathology, 10th ed.; Elsevier: Philadelphia, PA, USA, 2018. [Google Scholar]
- Karim, N.; Khan, I.; Khan, W.; Khan, I.; Khan, A.; Halim, S.A.; Khan, H.; Hussain, J.; Al-Harrasi, A. Anti-nociceptive and anti-inflammatory activities of asparacosin a involve selective cyclooxygenase 2 and inflammatory cytokines inhibition: An in-vitro, in-vivo, and in-silico approach. Front. Immunol. 2019, 10, 581. [Google Scholar] [CrossRef] [PubMed]
- Mansouri, M.T.; Hemmati, A.A.; Naghizadeh, B.; Mard, S.A.; Rezaie, A.; Ghorbanzadeh, B. A study of the mechanisms underlying the anti-inflammatory effect of ellagic acid in carrageenan-induced paw edema in rats. Indian J. Pharmacol. 2015, 47, 292–298. [Google Scholar] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Siqueira Patriota, L.L.; de Brito Marques Ramos, D.; e Silva, M.G.; dos Santos, A.C.L.A.; Silva, Y.A.; Paiva, P.M.G.; Pontual, E.V.; de Albuquerque, L.P.; Mendes, R.L.; Napoleão, T.H. Inhibition of Carrageenan-Induced Acute Inflammation in Mice by the Microgramma vacciniifolia Frond Lectin (MvFL). Polymers 2022, 14, 1609. https://doi.org/10.3390/polym14081609
de Siqueira Patriota LL, de Brito Marques Ramos D, e Silva MG, dos Santos ACLA, Silva YA, Paiva PMG, Pontual EV, de Albuquerque LP, Mendes RL, Napoleão TH. Inhibition of Carrageenan-Induced Acute Inflammation in Mice by the Microgramma vacciniifolia Frond Lectin (MvFL). Polymers. 2022; 14(8):1609. https://doi.org/10.3390/polym14081609
Chicago/Turabian Stylede Siqueira Patriota, Leydianne Leite, Dalila de Brito Marques Ramos, Mariana Gama e Silva, Angela Caroline Lima Amorim dos Santos, Yasmym Araújo Silva, Patrícia Maria Guedes Paiva, Emmanuel Viana Pontual, Lidiane Pereira de Albuquerque, Rosemairy Luciane Mendes, and Thiago Henrique Napoleão. 2022. "Inhibition of Carrageenan-Induced Acute Inflammation in Mice by the Microgramma vacciniifolia Frond Lectin (MvFL)" Polymers 14, no. 8: 1609. https://doi.org/10.3390/polym14081609