
Finding of Novel Galactose Utilizing Halomonas sp. YK44 for Polyhydroxybutyrate (PHB) Production

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemical Reagents
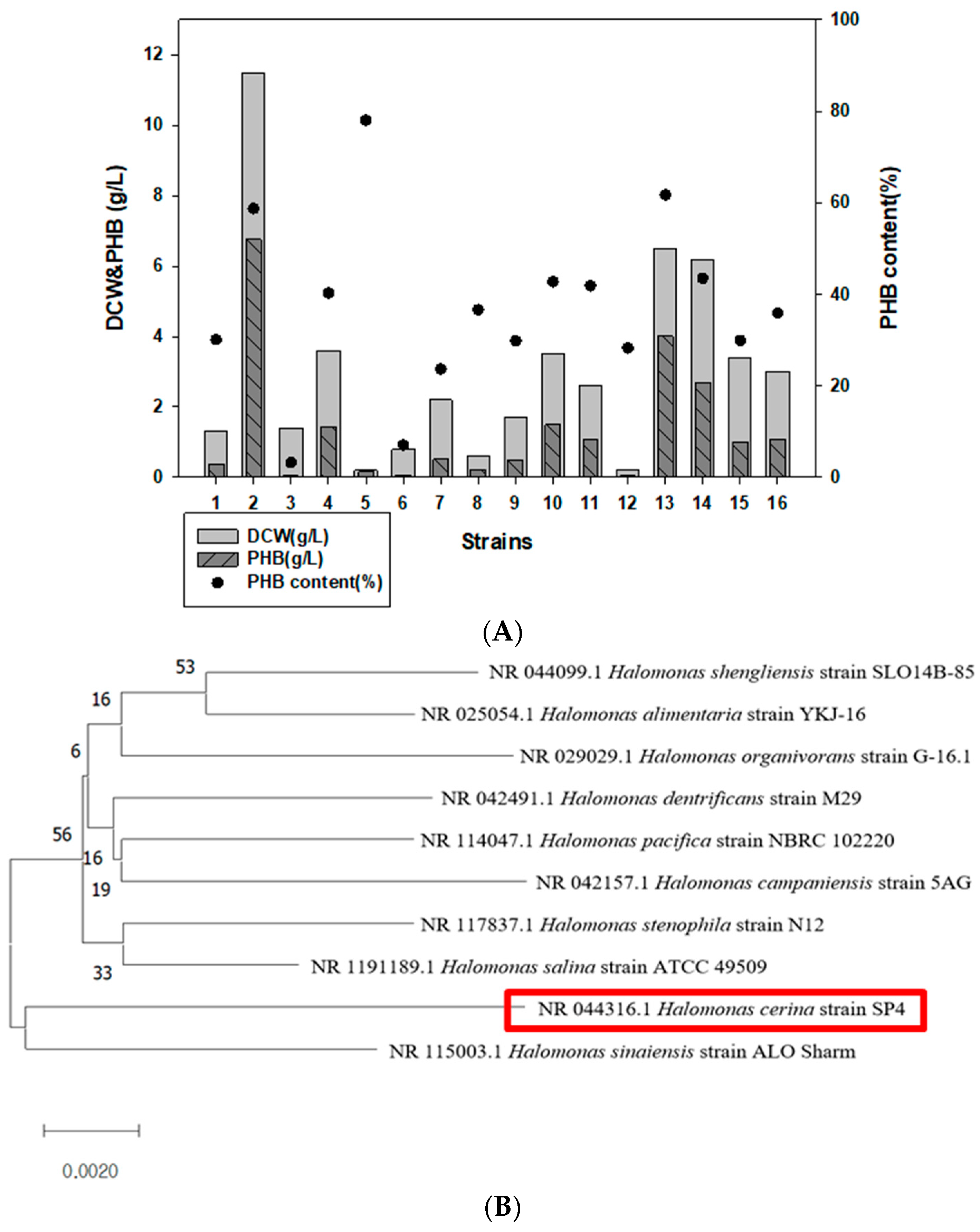
2.2. Sample Collection, Strain Isolation, and Phylogenetic Analysis
2.3. Plate Assay for Strain Characterization
2.4. Culture Conditions for PHA Synthesis
2.5. Analytical Methods
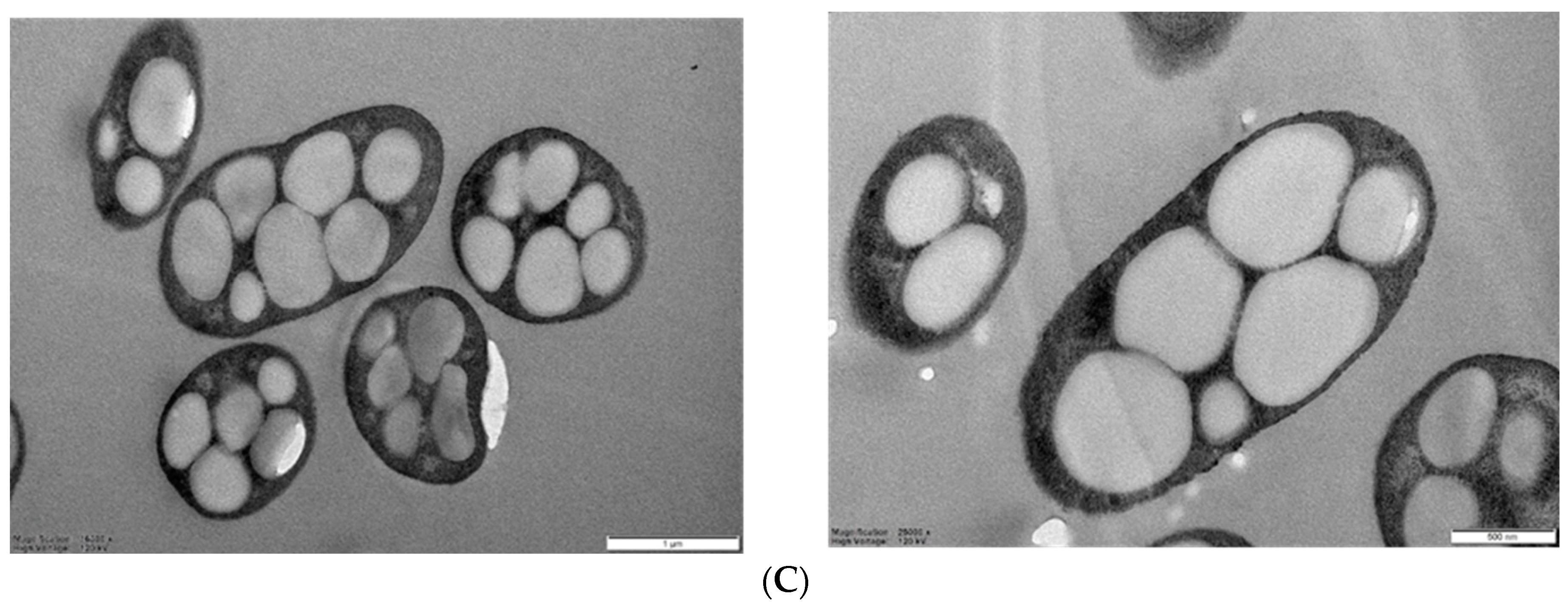
2.6. TEM Analysis
2.7. Polymer Extraction and Characterization
2.8. PHB Production in E. spinosum Hydrolysate
2.9. PHB Production Reusing Non-Sterilized and Sterilized Medium for Comparison
3. Results
3.1. Isolation and Characterization of Marine Strains for PHA Production
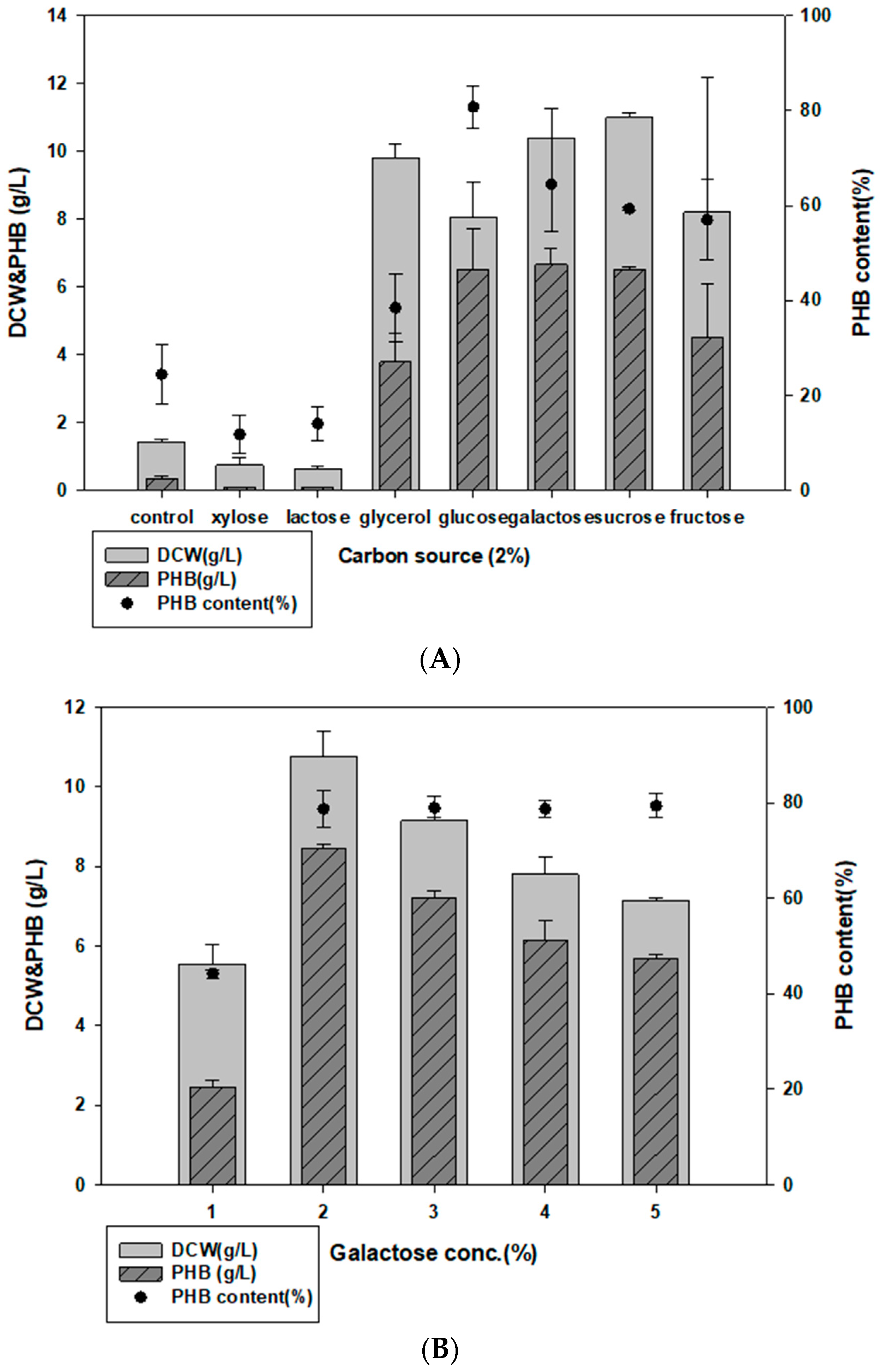
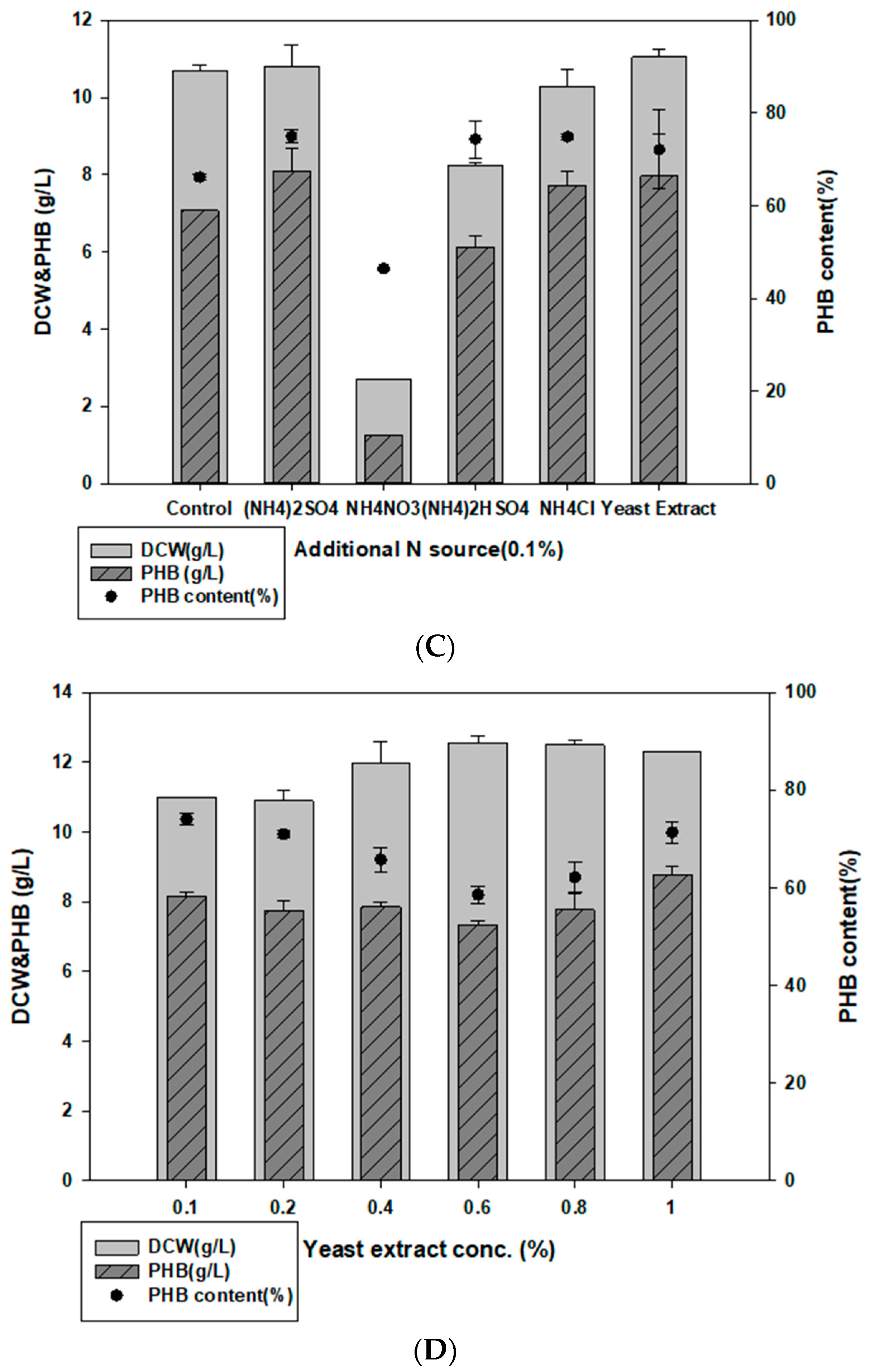
3.2. Evaluating Carbon and Nitrogen Sources for PHB Production
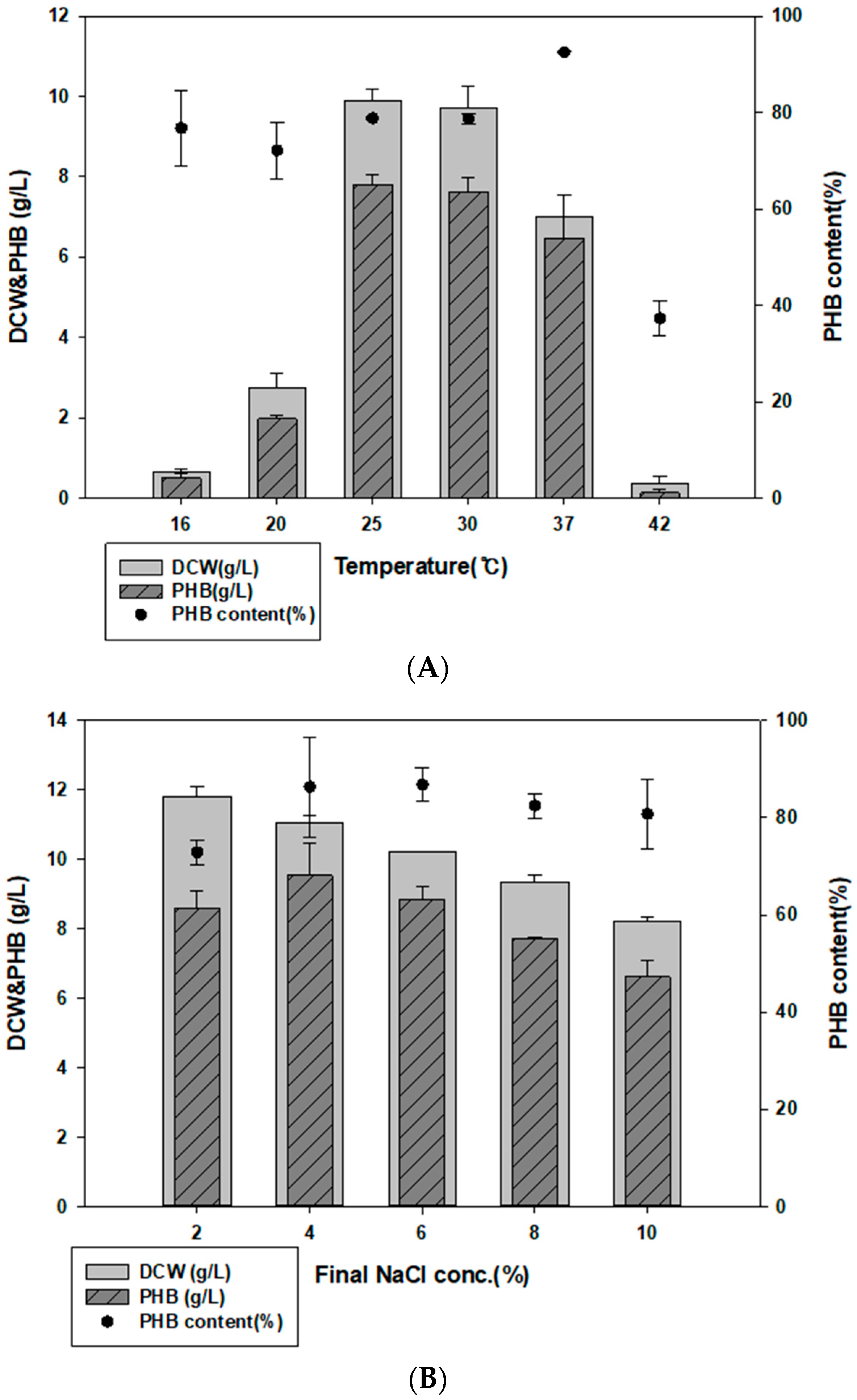
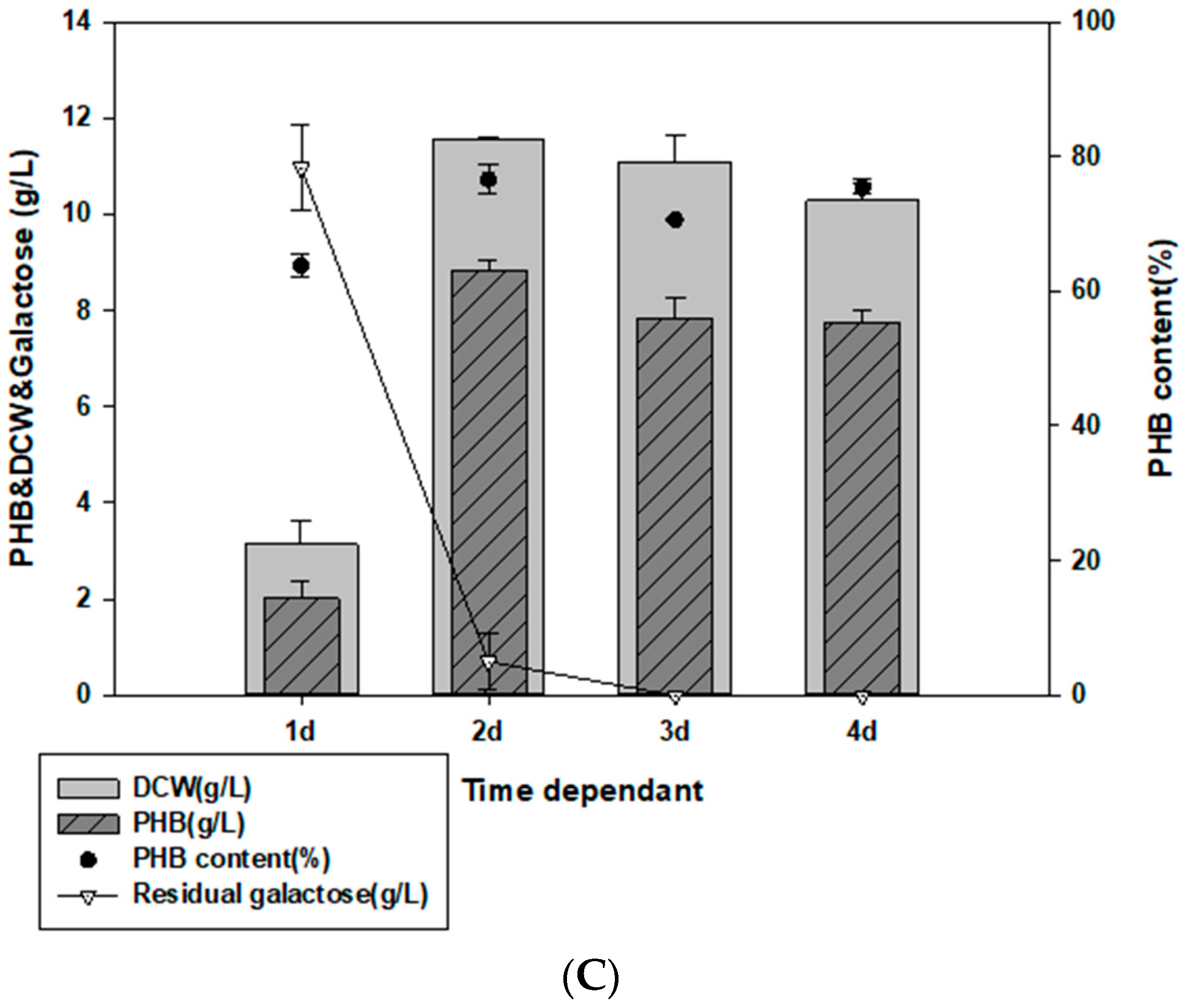
3.3. Examination of Culture Temperature, Time, and NaCl Concentration
3.4. Chemical Properties and Molecular Weight of PHB from Halomonas sp. YK44
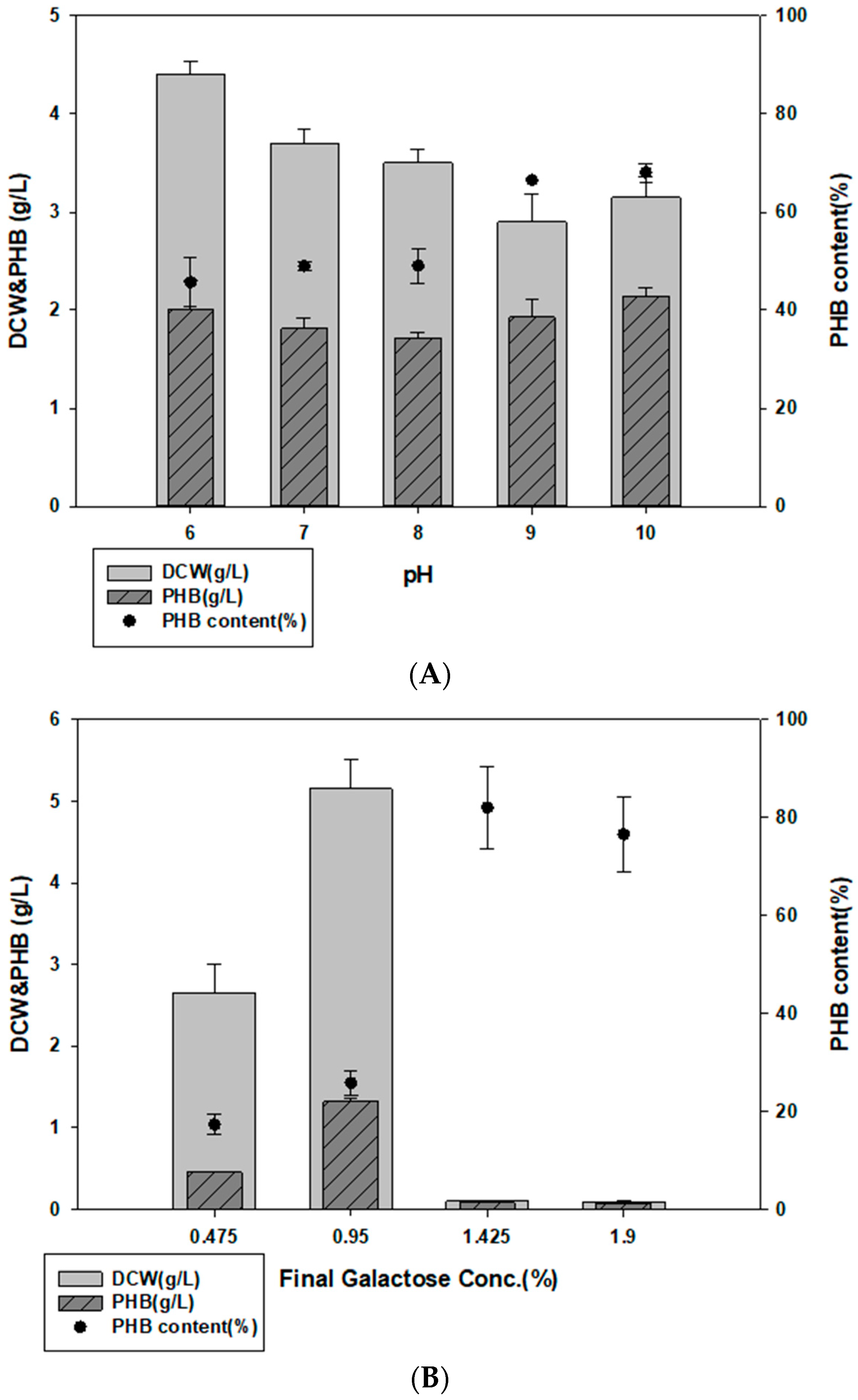
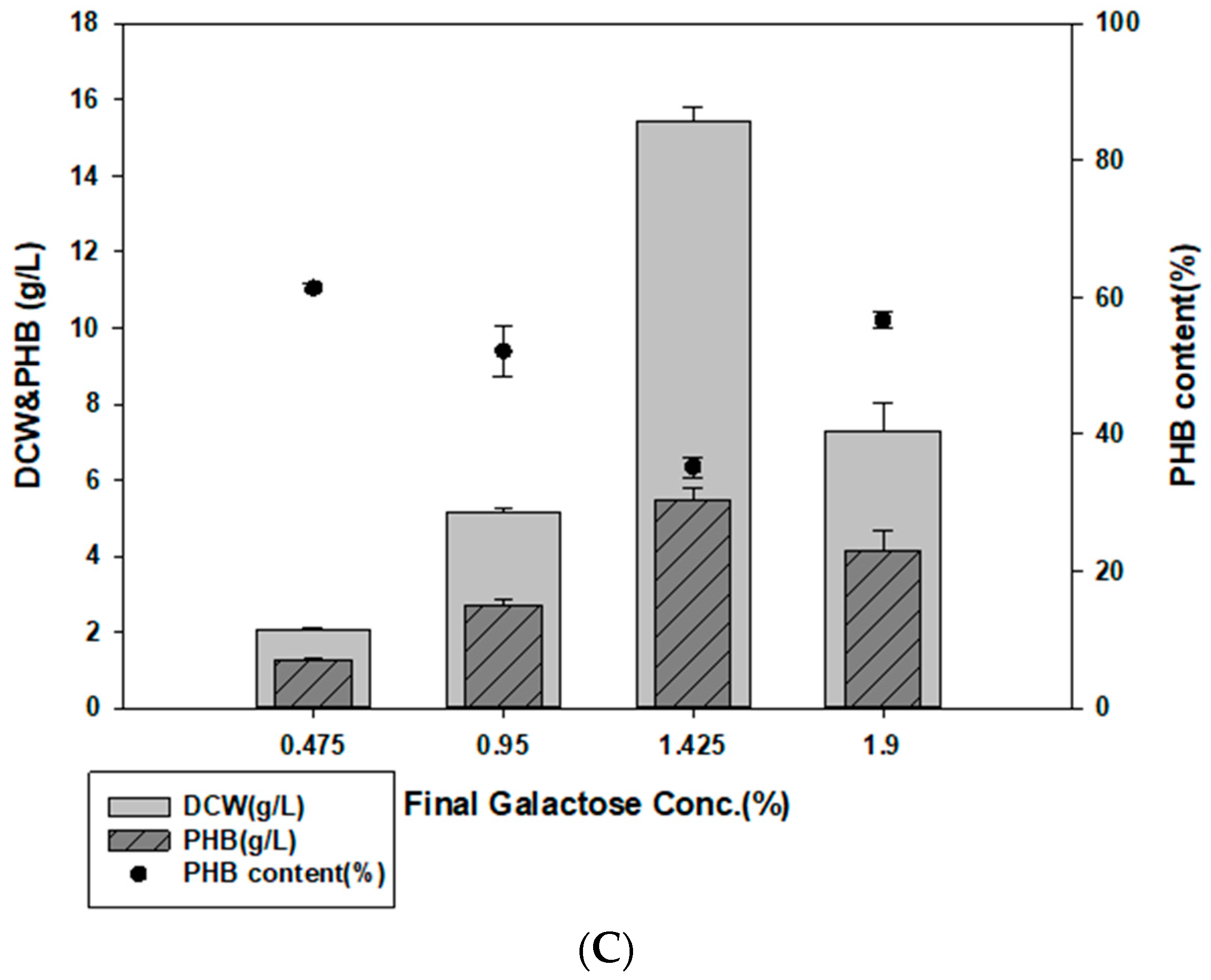
3.5. PHB Production using E. spinosum Hydrolysates
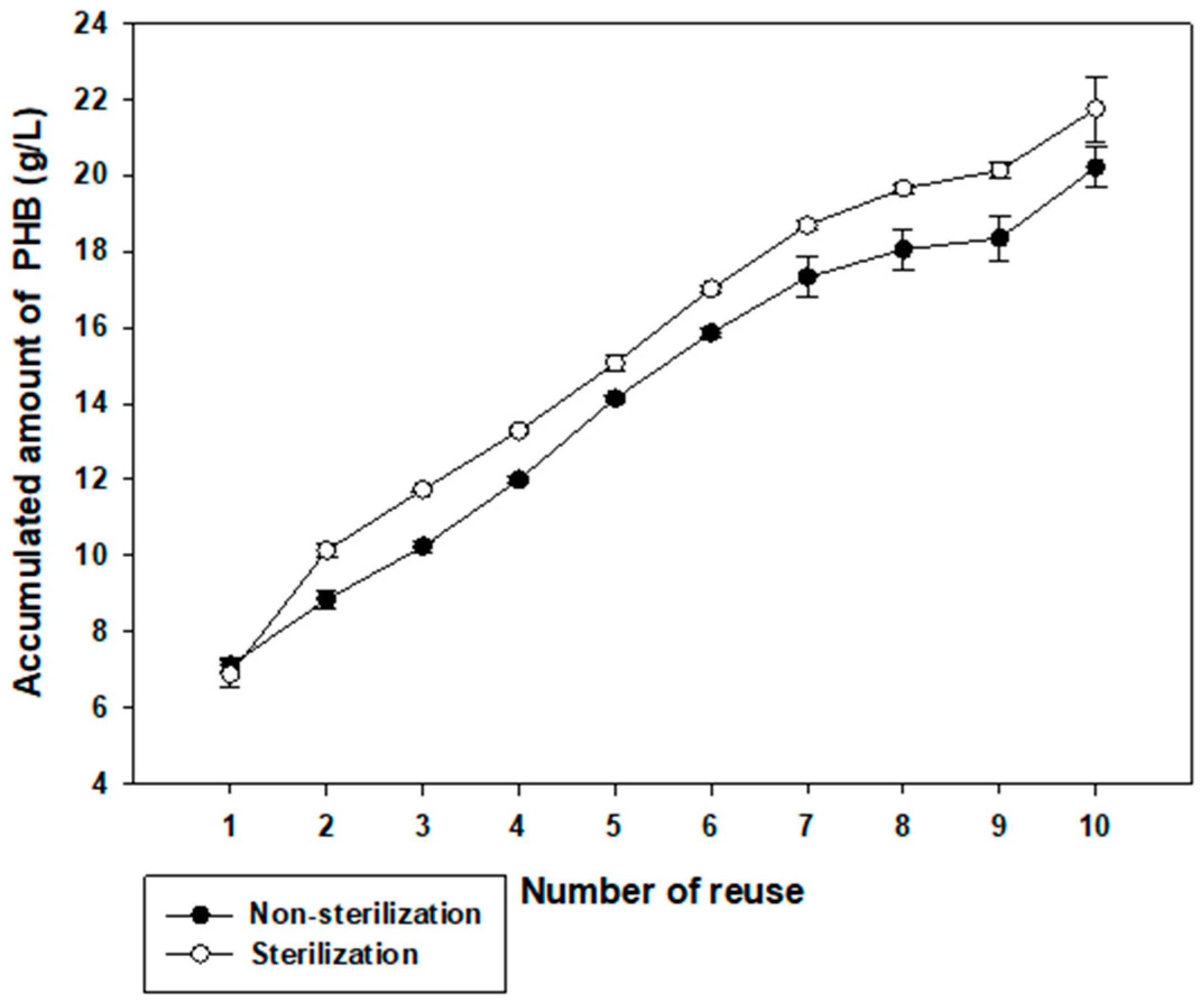
3.6. PHB Production in Reusing High Saline Non-Sterilized Medium and Sterilized Medium
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Barnes, D.K.A.; Galgani, F.; Thompson, R.C.; Barlaz, M. Accumulation and Fragmentation of Plastic Debris in Global Environments. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 1985–1998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chamas, A.; Moon, H.; Zheng, J.; Qiu, Y.; Tabassum, T.; Jang, J.H.; Abu-Omar, M.; Scott, S.L.; Suh, S. Degradation Rates of Plastics in the Environment. ACS Sustain. Chem. Eng. 2020, 8, 3494–3511. [Google Scholar] [CrossRef] [Green Version]
- Patrício Silva, A.L.; Prata, J.C.; Walker, T.R.; Duarte, A.C.; Ouyang, W.; Barcelò, D.; Rocha-Santos, T. Increased Plastic Pollution Due to COVID-19 Pandemic: Challenges and Recommendations. Chem. Eng. J. 2021, 405, 126683. [Google Scholar] [CrossRef] [PubMed]
- Vieira, M.G.A.; da Silva, M.A.; dos Santos, L.O.; Beppu, M.M. Natural-Based Plasticizers and Biopolymer Films: A Review. Eur. Polym. J. 2011, 47, 254–263. [Google Scholar] [CrossRef] [Green Version]
- Kabir, E.; Kaur, R.; Lee, J.; Kim, K.H.; Kwon, E.E. Prospects of Biopolymer Technology as an Alternative Option for Non-Degradable Plastics and Sustainable Management of Plastic Wastes. J. Clean. Prod. 2020, 258, 120536. [Google Scholar] [CrossRef]
- Costa, S.S.; Miranda, A.L.; de Morais, M.G.; Costa, J.A.V.; Druzian, J.I. Microalgae as Source of Polyhydroxyalkanoates (PHAs)—A Review. Int. J. Biol. Macromol. 2019, 131, 536–547. [Google Scholar] [CrossRef]
- Khosravi-Darani, K.; Mokhtari, Z.B.; Amai, T.; Tanaka, K. Microbial Production of Poly(Hydroxybutyrate) from C1 Carbon Sources. Appl. Microbiol. Biotechnol. 2013, 97, 1407–1424. [Google Scholar] [CrossRef]
- Venkateswar Reddy, M.; Mawatari, Y.; Yajima, Y.; Seki, C.; Hoshino, T.; Chang, Y.C. Poly-3-Hydroxybutyrate (PHB) Production from Alkylphenols, Mono and Poly-Aromatic Hydrocarbons Using Bacillus sp. CYR1: A New Strategy for Wealth from Waste. Bioresour. Technol. 2015, 192, 711–717. [Google Scholar] [CrossRef] [Green Version]
- Tsuge, T.; Hyakutake, M.; Mizuno, K. Class IV Polyhydroxyalkanoate (PHA) Synthases and PHA-Producing Bacillus. Appl. Microbiol. Biotechnol. 2015, 99, 6231–6240. [Google Scholar] [CrossRef]
- Hempel, F.; Bozarth, A.S.; Lindenkamp, N.; Klingl, A.; Zauner, S.; Linne, U.; Steinbüchel, A.; Maier, U.G. Microalgae as Bioreactors for Bioplastic Production. Microb. Cell Factories 2011, 10, 81. [Google Scholar] [CrossRef]
- Verlinden, R.A.J.; Hill, D.J.; Kenward, M.A.; Williams, C.D.; Radecka, I. Bacterial Synthesis of Biodegradable Polyhydroxyalkanoates. J. Appl. Microbiol. 2007, 102, 1437–1449. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.W.; Zhang, H.Y.; Lu, P.F.; Peng, Y.Z. Effects of Carbon Sources on the Enrichment of Halophilic Polyhydroxyalkanoate-Storing Mixed Microbial Culture in an Aerobic Dynamic Feeding Process. Sci. Rep. 2016, 6, 30766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Getachew, A.; Woldesenbet, F. Production of Biodegradable Plastic by Polyhydroxybutyrate (PHB) Accumulating Bacteria Using Low Cost Agricultural Waste Material. BMC Res. Notes 2016, 9, 509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saratale, R.G.; Saratale, G.D.; Cho, S.K.; Kim, D.S.; Ghodake, G.S.; Kadam, A.; Kumar, G.; Bharagava, R.N.; Banu, R.; Shin, H.S. Pretreatment of Kenaf (Hibiscus cannabinus L.) Biomass Feedstock for Polyhydroxybutyrate (PHB) Production and Characterization. Bioresour. Technol. 2019, 282, 75–80. [Google Scholar] [CrossRef]
- Sawant, S.S.; Salunke, B.K.; Kim, B.S. Consolidated Bioprocessing for Production of Polyhydroxyalkanotes from Red Algae Gelidium amansii. Int. J. Biol. Macromol. 2018, 109, 1012–1018. [Google Scholar] [CrossRef]
- Jung, K.A.; Lim, S.R.; Kim, Y.; Park, J.M. Potentials of Macroalgae as Feedstocks for Biorefinery. Bioresour. Technol. 2013, 135, 182–190. [Google Scholar] [CrossRef]
- Wei, N.; Quarterman, J.; Jin, Y.S. Marine Macroalgae: An Untapped Resource for Producing Fuels and Chemicals. Trends Biotechnol. 2013, 31, 70–77. [Google Scholar] [CrossRef]
- Rhein-Knudsen, N.; Ale, M.T.; Meyer, A.S. Marine Drugs Seaweed Hydrocolloid Production: An Update on Enzyme Assisted Extraction and Modification Technologies. Mar. Drugs 2015, 13, 3340–3359. [Google Scholar] [CrossRef]
- Estevez, J.M.; Ciancia, M.; Cerezo, A.S. The System of Sulfated Galactans from the Red Seaweed Gymnogongrus torulosus (Phyllophoraceae, Rhodophyta): Location and Structural Analysis. Carbohydr. Polym. 2008, 73, 594–605. [Google Scholar] [CrossRef]
- Campo, V.L.; Kawano, D.F.; da Silva, D.B.; Carvalho, I. Carrageenans: Biological Properties, Chemical Modifications and Structural Analysis—A Review. Carbohydr. Polym. 2009, 77, 167–180. [Google Scholar] [CrossRef]
- Kim, M.-J.; Kim, J.-S.; Ra, C.H.; Kim, S.-K. Bioethanol Production from Eucheuma spinosum Using Various Yeasts. KSBB J. 2013, 28, 315–318. [Google Scholar] [CrossRef] [Green Version]
- Ra, C.H.; Jung, J.H.; Sunwoo, I.Y.; Kang, C.H.; Jeong, G.T.; Kim, S.K. Detoxification of Eucheuma Spinosum Hydrolysates with Activated Carbon for Ethanol Production by the Salt-Tolerant Yeast Candida tropicalis. J. Microbiol. Biotechnol. 2015, 25, 856–862. [Google Scholar] [CrossRef] [PubMed]
- Adnan, H.; Porse, H. Culture of Eucheuma cottonii and Eucheuma spinosum in Indonesia. Hydrobiologia 1987, 151–152, 355–358. [Google Scholar] [CrossRef]
- Andersen, N.S.; Dolan, T.C.S.; Rees, D.A. Carrageenans. Part VII. Polysaccharides from Eucheuma spinosum and Eucheuma cottonii. The Covalent Structure of L-Carrageenan. J. Chem. Soc. Perkin 1 1973, 19, 2173–2176. [Google Scholar] [CrossRef] [PubMed]
- Pernicova, I.; Kucera, D.; Novackova, I.; Vodicka, J.; Kovalcik, A.; Obruca, S. Extremophiles—Platform Strains for Sustainable Production of Polyhydroxyalkanoates. In Materials Science Forum; Trans Tech Publications Ltd.: Wollerau, Switzerland, 2019; Volume 955, pp. 74–79. [Google Scholar]
- Thirumala, M.; Reddy, S.V.; Mahmood, S.K. Production and Characterization of PHB from Two Novel Strains of Bacillus Spp. Isolated from Soil and Activated Sludge. J. Ind. Microbiol. Biotechnol. 2010, 37, 271–278. [Google Scholar] [CrossRef]
- Kucera, D.; Pernicová, I.; Kovalcik, A.; Koller, M.; Mullerova, L.; Sedlacek, P.; Mravec, F.; Nebesarova, J.; Kalina, M.; Marova, I.; et al. Characterization of the Promising Poly(3-Hydroxybutyrate) Producing Halophilic Bacterium Halomonas halophila. Bioresour. Technol. 2018, 256, 552–556. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.; Sudesh, K.; Bazire, A.; Elain, A.; Tan, H.T.; Lim, H.; Bruzaud, S. PHA Production and Pha Synthases of the Halophilic Bacterium Halomonas sp. SF2003. Bioengineering 2020, 7, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poli, A.; Nicolaus, B.; Denizci, A.A.; Yavuzturk, B.; Kazan, D. Halomonas Smyrnensis sp. Nov., a Moderately Halophilic, Exopolysaccharide-Producing Bacterium. Int. J. Syst. Evol. Microbiol. 2013, 63, 10–18. [Google Scholar] [CrossRef]
- Xu, X.W.; Wu, Y.H.; Zhou, Z.; Wang, C.S.; Zhou, Y.G.; Zhang, H.B.; Wang, Y.; Wu, M. Halomonas saccharevitans sp. Nov., Halomonas arcis sp. Nov. and Halomonas subterranea sp. Nov., Halophilic Bacteria Isolated from Hypersaline Environments of China. Int. J. Syst. Evol. Microbiol. 2007, 57, 1619–1624. [Google Scholar] [CrossRef]
- Ling, C.; Qiao, G.Q.; Shuai, B.W.; Song, K.N.; Yao, W.X.; Jiang, X.R.; Chen, G.Q. Engineering Self-Flocculating Halomonas campaniensis for Wastewaterless Open and Continuous Fermentation. Biotechnol. Bioeng. 2019, 116, 805–815. [Google Scholar] [CrossRef]
- Fu, X.Z.; Tan, D.; Aibaidula, G.; Wu, Q.; Chen, J.C.; Chen, G.Q. Development of Halomonas TD01 as a Host for Open Production of Chemicals. Metab. Eng. 2014, 23, 78–91. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Lee, H.J.; Kim, S.H.; Suh, M.J.; Cho, J.Y.; Ham, S.; Jeon, J.M.; Yoon, J.J.; Bhatia, S.K.; Gurav, R.; et al. Screening of the Strictly Xylose-Utilizing Bacillus sp. SM01 for Polyhydroxybutyrate and Its Co-Culture with Cupriavidus necator NCIMB 11599 for Enhanced Production of PHB. Int. J. Biol. Macromol. 2021, 181, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.L.; Bhatia, S.K.; Gurav, R.; Choi, T.R.; Kim, H.J.; Song, H.S.; Park, J.Y.; Han, Y.H.; Lee, S.M.; Park, S.L.; et al. Fructose Based Hyper Production of Poly-3-Hydroxybutyrate from Halomonas sp. YLGW01 and Impact of Carbon Sources on Bacteria Morphologies. Int. J. Biol. Macromol. 2020, 154, 929–936. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.R.; Choi, T.R.; Han, Y.H.; Park, Y.L.; Park, J.Y.; Song, H.S.; Yang, S.Y.; Bhatia, S.K.; Gurav, R.; Park, H.A.; et al. Production of Blue-Colored Polyhydroxybutyrate (PHB) by One-Pot Production and Coextraction of Indigo and PHB from Recombinant Escherichia coli. Dye. Pigment. 2020, 173, 107889. [Google Scholar] [CrossRef]
- Hong, J.W.; Song, H.S.; Moon, Y.M.; Hong, Y.G.; Bhatia, S.K.; Jung, H.R.; Choi, T.R.; Yang, S.Y.; Park, H.Y.; Choi, Y.K.; et al. Polyhydroxybutyrate Production in Halophilic Marine Bacteria Vibrio Proteolyticus Isolated from the Korean Peninsula. Bioprocess. Biosyst. Eng. 2019, 42, 603–610. [Google Scholar] [CrossRef]
- Bhatia, S.K.; Gurav, R.; Choi, T.R.; Jung, H.R.; Yang, S.Y.; Moon, Y.M.; Song, H.S.; Jeon, J.M.; Choi, K.Y.; Yang, Y.H. Bioconversion of Plant Biomass Hydrolysate into Bioplastic (Polyhydroxyalkanoates) Using Ralstonia eutropha 5119. Bioresour. Technol. 2019, 271, 306–315. [Google Scholar] [CrossRef]
- Bhatia, S.K.; Wadhwa, P.; Hong, J.W.; Hong, Y.G.; Jeon, J.M.; Lee, E.S.; Yang, Y.H. Lipase Mediated Functionalization of Poly(3-Hydroxybutyrate-Co-3-Hydroxyvalerate) with Ascorbic Acid into an Antioxidant Active Biomaterial. Int. J. Biol. Macromol. 2019, 123, 117–123. [Google Scholar] [CrossRef]
- Tohme, S.; Hacıosmanoğlu, G.G.; Eroğlu, M.S.; Kasavi, C.; Genç, S.; Can, Z.S.; Toksoy Oner, E. Halomonas smyrnensis as a Cell Factory for Co-Production of PHB and Levan. Int. J. Biol. Macromol. 2018, 118, 1238–1246. [Google Scholar] [CrossRef]
- Kulkarni, S.O.; Kanekar, P.P.; Jog, J.P.; Sarnaik, S.S.; Nilegaonkar, S.S. Production of Copolymer, Poly (Hydroxybutyrate-Co-Hydroxyvalerate) by Halomonas campisalis MCM B-1027 Using Agro-Wastes. Int. J. Biol. Macromol. 2015, 72, 784–789. [Google Scholar] [CrossRef]
- Jiang, X.R.; Yao, Z.H.; Chen, G.Q. Controlling Cell Volume for Efficient PHB Production by Halomonas. Metab. Eng. 2017, 44, 30–37. [Google Scholar] [CrossRef]
- Trakunjae, C.; Boondaeng, A.; Apiwatanapiwat, W.; Kosugi, A.; Arai, T.; Sudesh, K.; Vaithanomsat, P. Enhanced Polyhydroxybutyrate (PHB) Production by Newly Isolated Rare Actinomycetes Rhodococcus sp. Strain BSRT1-1 Using Response Surface Methodology. Sci. Rep. 2021, 11, 1896. [Google Scholar] [CrossRef] [PubMed]
- Radhika, D.; Murugesan, A.G. Bioproduction, Statistical Optimization and Characterization of Microbial Plastic (Poly 3-Hydroxy Butyrate) Employing Various Hydrolysates of Water Hyacinth (Eichhornia crassipes) as Sole Carbon Source. Bioresour. Technol. 2012, 121, 83–92. [Google Scholar] [CrossRef]
- Wellen, R.M.R.; Rabello, M.S.; Fechine, G.J.M.; Canedo, E.L. The Melting Behaviour of Poly(3-Hydroxybutyrate) by DSC. Reproducibility Study. Polym. Test 2013, 32, 215–220. [Google Scholar] [CrossRef]
- Wendlandt, K.D.; Geyer, W.; Mirschel, G.; Al-Haj Hemidi, F. Possibilities for Controlling a PHB Accumulation Process Using Various Analytical Methods. J. Biotechnol. 2005, 117, 119–129. [Google Scholar] [CrossRef]
- Alsafadi, D.; Ibrahim, M.I.; Alamry, K.A.; Hussein, M.A.; Mansour, A. Utilizing the Crop Waste of Date Palm Fruit to Biosynthesize Polyhydroxyalkanoate Bioplastics with Favorable Properties. Sci. Total Environ. 2020, 737, 139716. [Google Scholar] [CrossRef]
- Arminda, M.; Josúe, C.; Cristina, D.; Fabiana, S.; Yolanda, M. Use of Activated Carbons for Detoxification of a Lignocellulosic Hydrolysate: Statistical Optimisation. J. Environ. Manag. 2021, 296, 113320. [Google Scholar] [CrossRef] [PubMed]
- Sahu, A.K.; Srivastava, V.C.; Mall, I.D.; Lataye, D.H. Adsorption of Furfural from Aqueous Solution onto Activated Carbon: Kinetic, Equilibrium and Thermodynamic Study. Sep. Sci. Technol. 2008, 43, 1239–1259. [Google Scholar] [CrossRef]
- Tan, D.; Xue, Y.S.; Aibaidula, G.; Chen, G.Q. Unsterile and Continuous Production of Polyhydroxybutyrate by Halomonas TD01. Bioresour. Technol. 2011, 102, 8130–8136. [Google Scholar] [CrossRef]
- Park, Y.L.; Song, H.S.; Choi, T.R.; Lee, S.M.; Park, S.L.; Lee, H.S.; Kim, H.J.; Bhatia, S.K.; Gurav, R.; Park, K.; et al. Revealing of Sugar Utilization Systems in Halomonas sp. YLGW01 and Application for Poly(3-Hydroxybutyrate) Production with Low-Cost Medium and Easy Recovery. Int. J. Biol. Macromol. 2021, 167, 151–159. [Google Scholar] [CrossRef]
- Parawira, W.; Tekere, M. Biotechnological Strategies to Overcome Inhibitors in Lignocellulose Hydrolysates for Ethanol Production: Review. Crit. Rev. Biotechnol. 2011, 31, 20–31. [Google Scholar] [CrossRef]
- Lee, J.M.; Venditti, R.A.; Jameel, H.; Kenealy, W.R. Detoxification of Woody Hydrolyzates with Activated Carbon for Bioconversion to Ethanol by the Thermophilic Anaerobic Bacterium Thermoanaerobacterium saccharolyticum. Biomass Bioenergy 2011, 35, 626–636. [Google Scholar] [CrossRef]
- El-malek, F.A.; Farag, A.; Omar, S.; Khairy, H. Polyhydroxyalkanoates (PHA) from Halomonas Pacifica ASL10 and Halomonas salifodiane ASL11 Isolated from Mariout Salt Lakes. Int. J. Biol. Macromol. 2020, 161, 1318–1328. [Google Scholar] [CrossRef] [PubMed]
- Coronado, M.-J.; Vargas, C.; Hofemeister, J.; Ventosa, A.; Nieto, J.J. Production and Biochemical Characterization of an α-Amylase from the Moderate Halophile Halomonas meridiana. FEMS Microbiol. Lett. 2000, 183, 67–71. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganism | Carbon Source | DCW (g/L) | PHB Content (wt%) | PHB (g/L) | Culture | Reference |
|---|---|---|---|---|---|---|
| Halomonas halophila | Galactose | 4.22 ± 0.10 | 80.7 ± 2.0 | 3.41 ± 0.12 | Batch | [27] |
| Halomonas salina | Galactose | 0.97 ± 0.03 | 12.3 ± 0.48 | 0.12 ± 0.01 | Batch | [25] |
| Halomonas organivorans | Galactose | 5.80 ± 0.22 | 90.55 ± 4.08 | 5.61 ± 0.01 | Batch | [25] |
| Bacillus sp. 112A | Galactose | 1.02 | 35.50 | 0.879 | Batch | [26] |
| Halomonas sp. SF2003 | Galactose | 3.16 | 39 | 1.23 | Batch | [28] |
| Similar Species | Strain | Isolated Temperature | Resource Number | |
|---|---|---|---|---|
| 1 | Halomonas alkalicola | DH-10 | 25 °C | MABIK MI00000003 |
| 2 | Halomonas cerina | YK44 | 20 °C | MABIK MI00000284 |
| 3 | Halomonas sulfidaeris | J05-14M-11R | 20 °C | MABIK MI00000306 |
| 4 | Halomonas fontilapidosi | O12 | 25 °C | MABIK MI00000338 |
| 5 | Halomonas gomseomensis | CJCa107 | 25 °C | MABIK MI00000370 |
| 6 | Halomonas arcis | CJCbj078 | 25 °C | MABIK MI00000396 |
| 7 | Halomonas janggokensis | CJCbj082 | 25 °C | MABIK MI00000412 |
| 8 | Halomonas salicampi | CJCbj041 | 25 °C | MABIK MI00000417 |
| 9 | Halomonas lutescens | CJCbj058 | 25 °C | MABIK MI00000427 |
| 10 | Halomonas fontilapidosi | MEBiC12169 | 25 °C | MABIK MI00005505 |
| 11 | Halomonas campaniensis | S510 | 25 °C | MABIK MI00005561 |
| 12 | Halomonas saccharevitans | MJ005 | 25 °C | MABIK MI00005664 |
| 13 | Halomonas shengliensis | MEBiC12098 | 25 °C | MABIK MI00005446 |
| 14 | Halomonas denitrificans | MEBiC13328 | 25 °C | MABIK MI00005839 |
| 15 | Halomonas aestuarii | MEBiC13369 | 25 °C | MABIK MI00005871 |
| 16 | Halomonas lutea | 15A021 | 27 °C | MABIK MI00005926 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, H.J.; Kim, S.H.; Cho, D.H.; Kim, B.C.; Bhatia, S.K.; Lee, J.; Jeon, J.-M.; Yoon, J.-J.; Yang, Y.-H. Finding of Novel Galactose Utilizing Halomonas sp. YK44 for Polyhydroxybutyrate (PHB) Production. Polymers 2022, 14, 5407. https://doi.org/10.3390/polym14245407
Jung HJ, Kim SH, Cho DH, Kim BC, Bhatia SK, Lee J, Jeon J-M, Yoon J-J, Yang Y-H. Finding of Novel Galactose Utilizing Halomonas sp. YK44 for Polyhydroxybutyrate (PHB) Production. Polymers. 2022; 14(24):5407. https://doi.org/10.3390/polym14245407
Chicago/Turabian StyleJung, Hee Ju, Su Hyun Kim, Do Hyun Cho, Byung Chan Kim, Shashi Kant Bhatia, Jongbok Lee, Jong-Min Jeon, Jeong-Jun Yoon, and Yung-Hun Yang. 2022. "Finding of Novel Galactose Utilizing Halomonas sp. YK44 for Polyhydroxybutyrate (PHB) Production" Polymers 14, no. 24: 5407. https://doi.org/10.3390/polym14245407