
Regulation of Mesenchymal Stem Cell Morphology Using Hydrogel Substrates with Tunable Topography and Photoswitchable Stiffness
 , and
, and 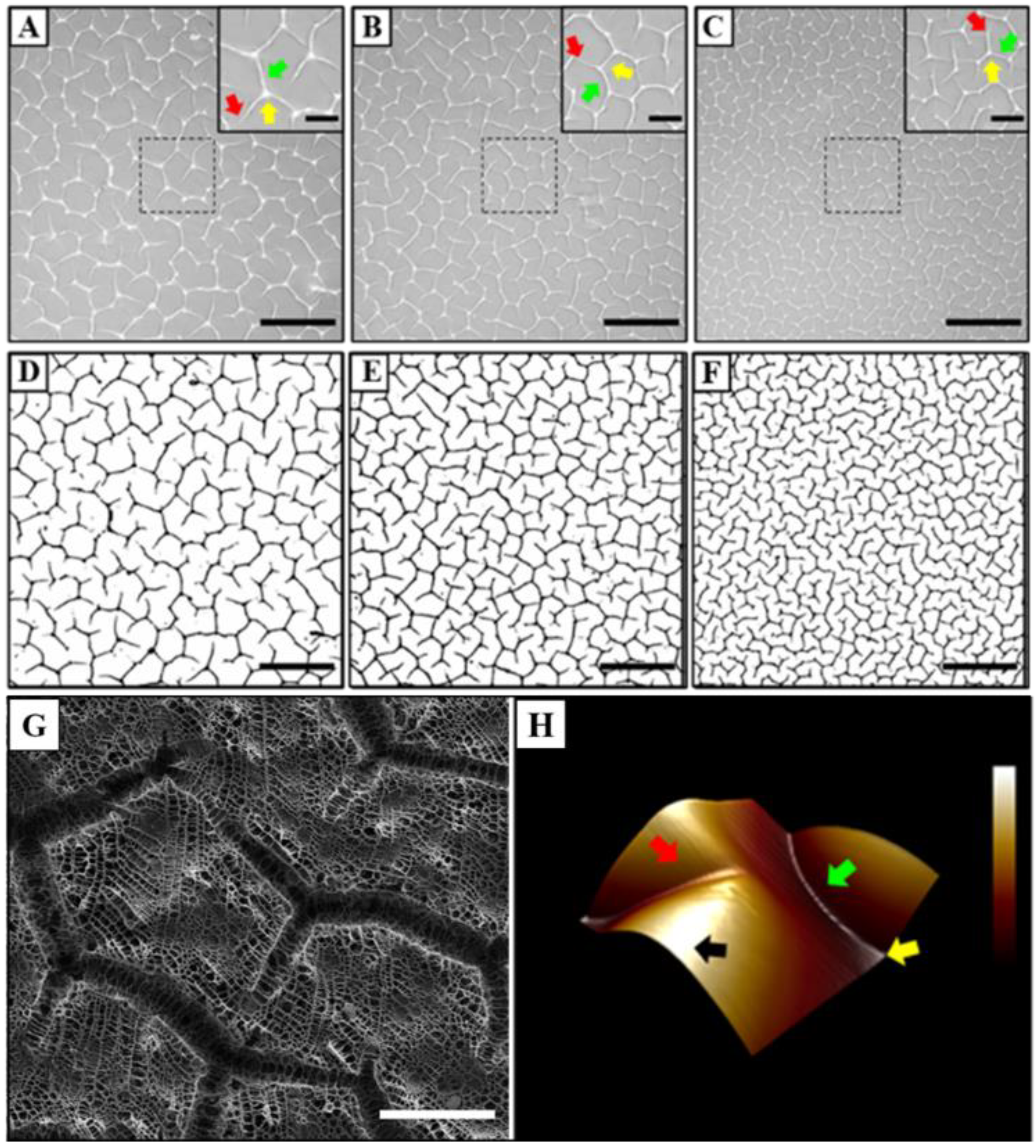{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Synthesis of AZO Crosslinked Polyacrylamide Hydrogels
2.3. Characterisation of Topographic Density
2.4. Modulation of Matrix Stiffness and Cell Seeding
2.5. Analysis of Cell Morphology and Size
2.6. Live Cell Imaging
3. Results
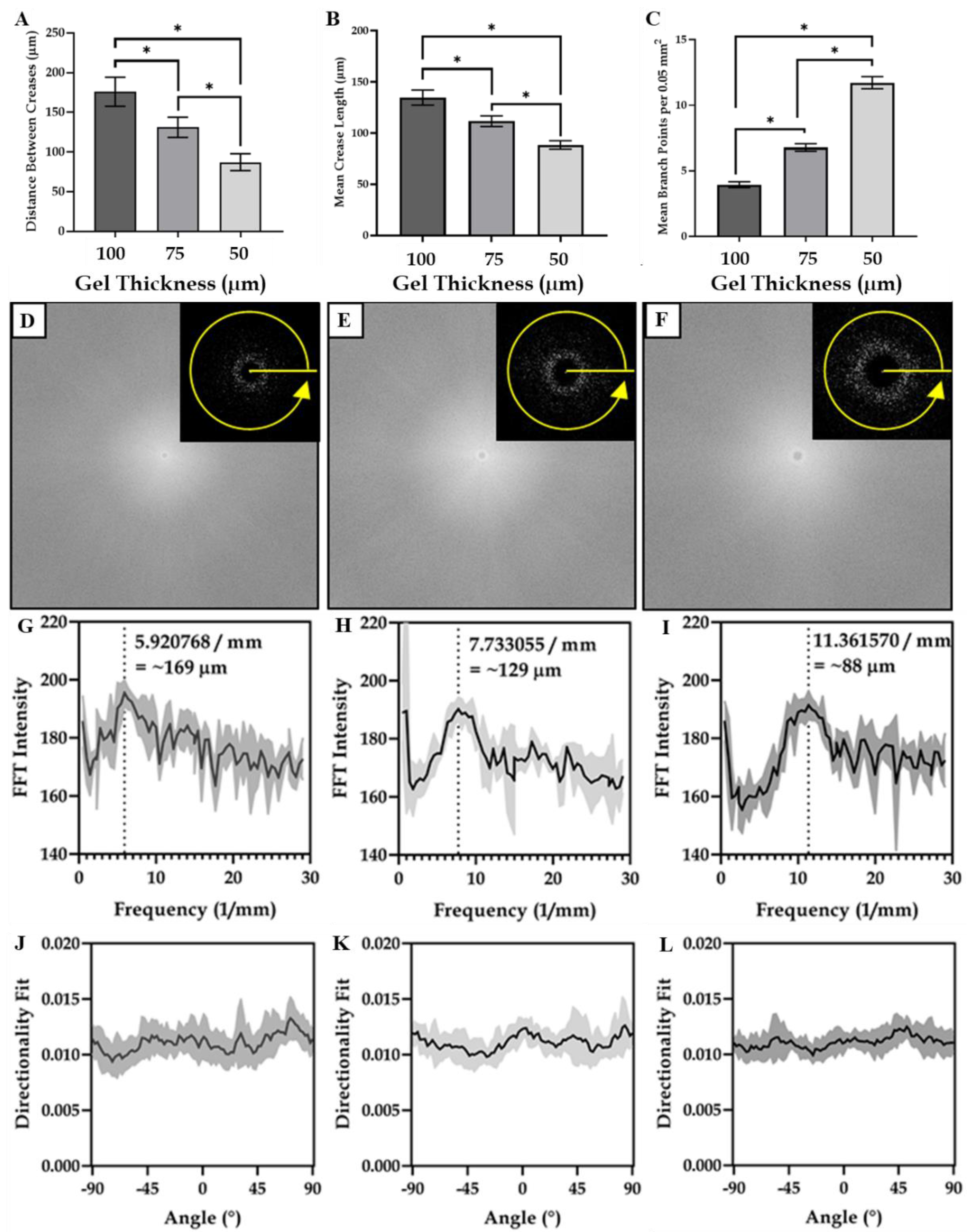
3.1. Characterisation of Hydrogel Surface Topography
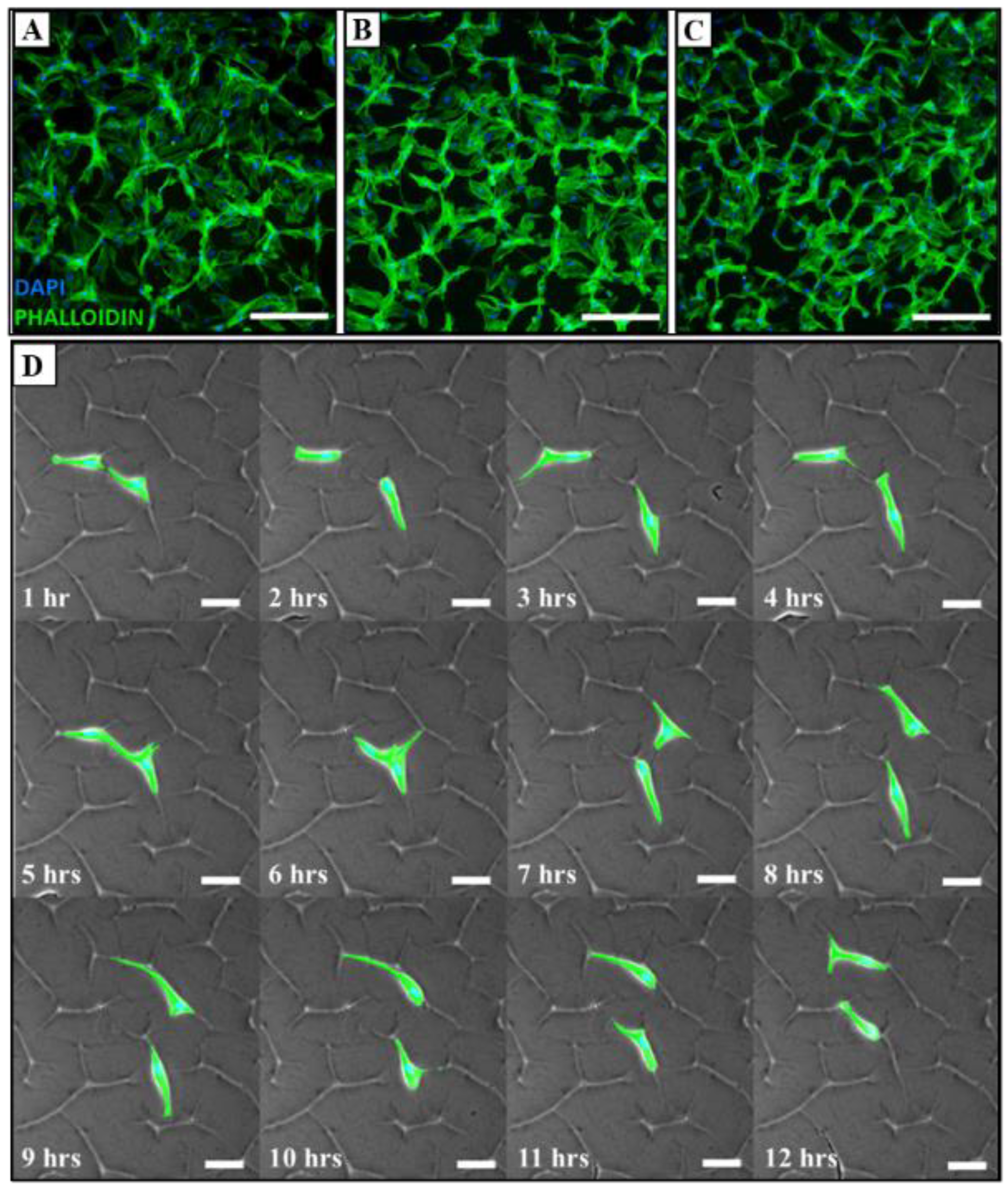
3.2. Adhesion and Migration Analysis of MSCs
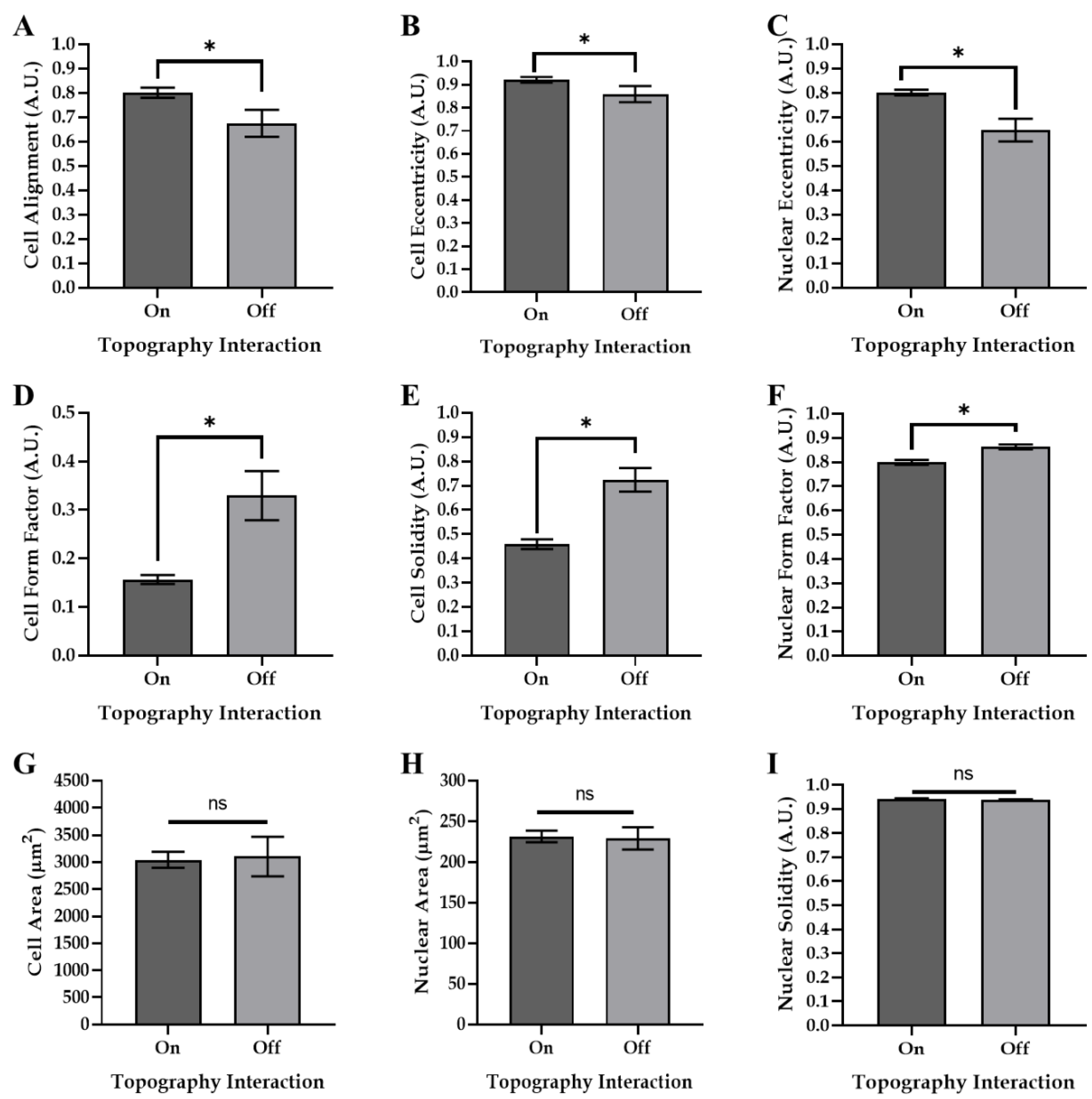
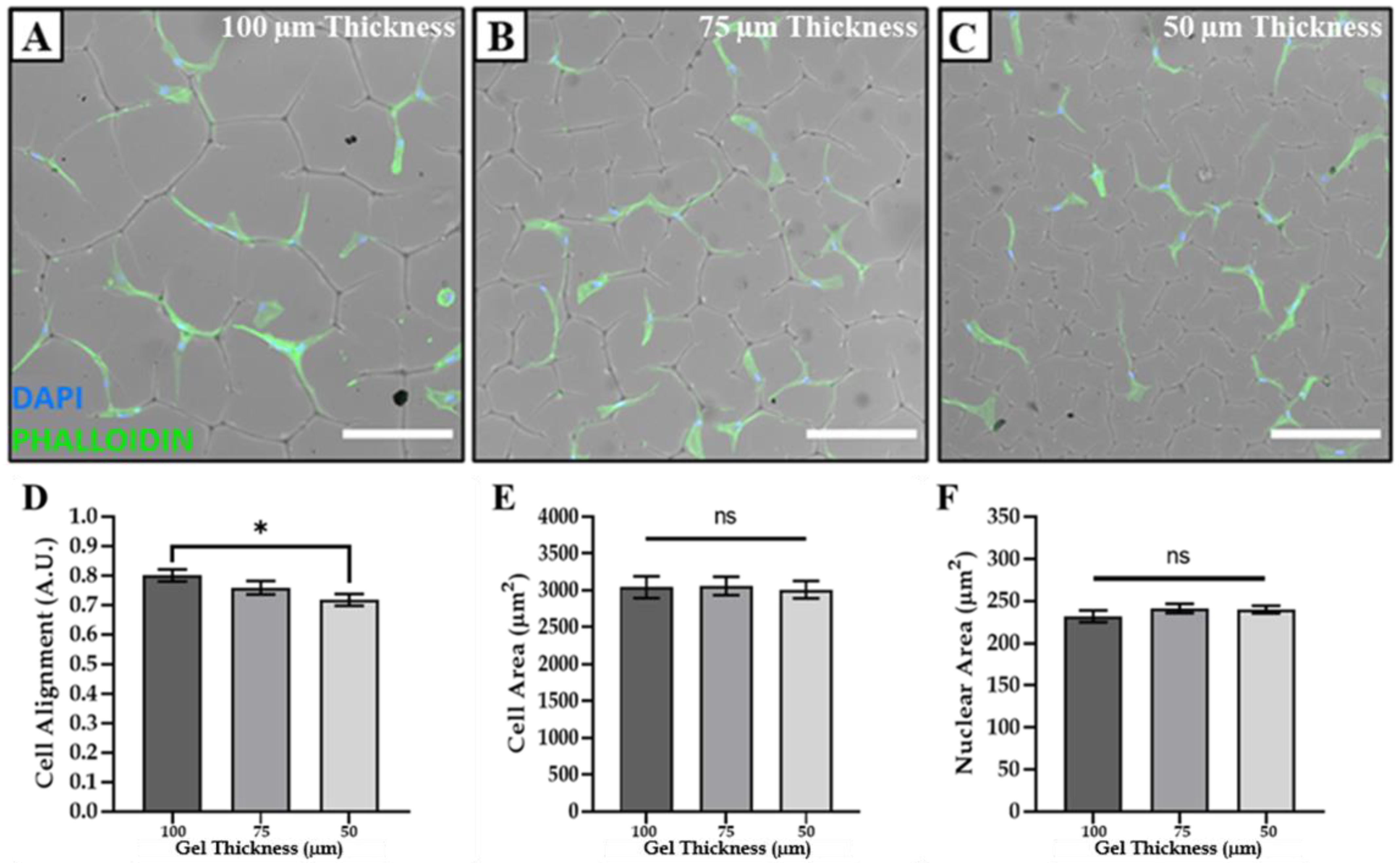
3.3. Modulating Cell Morphology and Alignment via Tuneable Surface Topographies
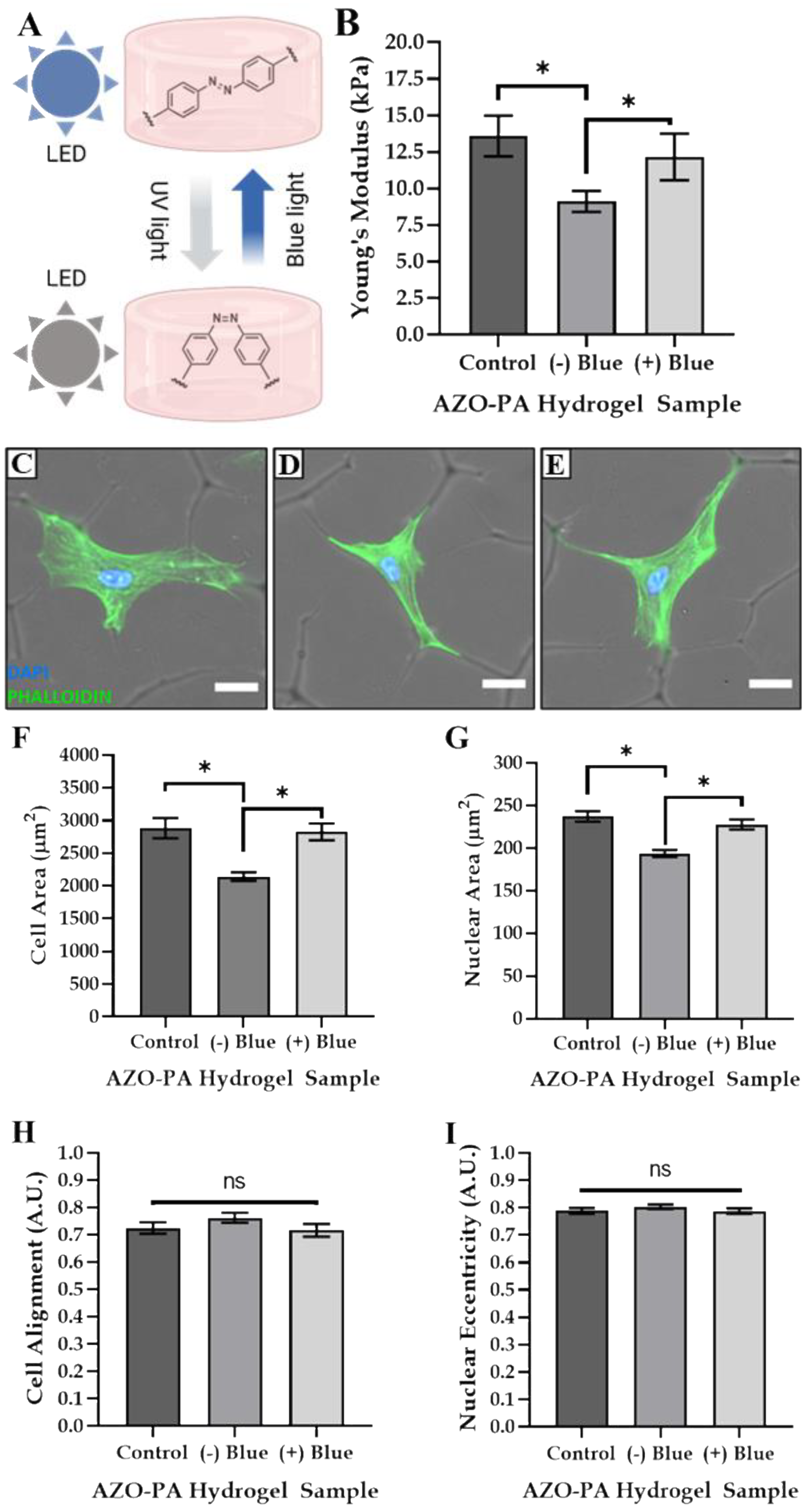
3.4. Tailoring Matrix Stiffness and Cell Size
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Prasad, A.; Alizadeh, E. Cell form and function: Interpreting and controlling the shape of adherent cells. Trends Biotechnol. 2019, 37, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Schnabel, M.; Marlovits, S.; Eckhoff, G.; Fichtel, I.; Gotzen, L.; Vécsei, V.; Schlegel, J. Dedifferentiation-associated changes in morphology and gene expression in primary human articular chondrocytes in cell culture. Osteoarthr. Cartil. 2002, 10, 62–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooke, M.E.; Pearson, M.J.; Moakes, R.J.A.; Weston, C.J.; Davis, E.T.; Jones, S.W.; Grover, L.M. Geometric confinement is required for recovery and maintenance of chondrocyte phenotype in alginate. APL Bioeng. 2017, 1, 016104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, W.; Kim, G. A functional bioink and its application in myoblast alignment and differentiation. Chem. Eng. J. 2019, 366, 150–162. [Google Scholar] [CrossRef]
- Martínez, E.C.; Hernández, J.C.R.; Machado, M.; Mano, J.F.; Ribelles, J.L.G.; Pradas, M.M.; Sánchez, M.S. Human Chondrocyte Morphology, Its Dedifferentiation, and Fibronectin Conformation on Different PLLA Microtopographies. Tissue Eng. Part A 2008, 14, 1751–1762. [Google Scholar] [CrossRef] [PubMed]
- Kuznetsova, A.V.; Kurinov, A.M.; Rzhanova, L.A.; Aleksandrova, M.A. Mechanisms of Dedifferentiation of Adult Human Retinal Pigment Epithelial Cells in vitro. Morphological and Molecular Genetic Analysis. Cell Tissue Biol. 2019, 13, 107–119. [Google Scholar] [CrossRef]
- MacGregor, M.; Williams, R.; Downes, J.; Bachhuka, A.; Vasilev, K. The Role of Controlled Surface Topography and Chemistry on Mouse Embryonic Stem Cell Attachment, Growth and Self-Renewal. Materials 2017, 10, 1081. [Google Scholar] [CrossRef] [Green Version]
- Cortella, L.R.X.; Cestari, I.A.; Lahuerta, R.D.; Araña, M.C.; Soldera, M.; Rank, A.; Lasagni, A.F.; Cestari, I.N. Conditioning of hiPSC-derived cardiomyocytes using surface topography obtained with high throughput technology. Biomed. Mater. 2021, 16, 065007. [Google Scholar] [CrossRef]
- Abagnale, G.; Sechi, A.; Steger, M.; Zhou, Q.; Kuo, C.-C.; Aydin, G.; Schalla, C.; Müller-Newen, G.; Zenke, M.; Costa, I.G.; et al. Surface Topography Guides Morphology and Spatial Pat-terning of Induced Pluripotent Stem Cell Colonies. Stem Cell Rep. 2017, 9, 654–666. [Google Scholar] [CrossRef] [Green Version]
- Hou, Y.; Xie, W.; Yu, L.; Camacho, L.C.; Nie, C.; Zhang, M.; Haag, R.; Wei, Q. Surface Roughness Gradients Reveal Topography-Specific Mecha-nosensitive Responses in Human Mesenchymal Stem Cells. Small 2020, 16, 1905422. [Google Scholar] [CrossRef]
- Tsimbouri, P.; Gadegaard, N.; Burgess, K.; White, K.; Reynolds, P.; Herzyk, P.; Oreffo, R.; Dalby, M.J. Nanotopographical Effects on Mesenchymal Stem Cell Morphology and Phenotype. J. Cell. Biochem. 2014, 115, 380–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeung, T.; Georges, P.C.; Flanagan, L.A.; Marg, B.; Ortiz, M.; Funaki, M.; Zahir, N.; Ming, W.; Weaver, V.; Janmey, P.A. Effects of substrate stiffness on cell morphology, cytoskeletal structure, and adhesion. Cell Motil. Cytoskelet. 2005, 60, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Chi, G.; Li, P.; Lv, S.; Xu, J.; Xu, Z.; Xia, Y.; Tan, Y.; Xu, J.; Li, L.; et al. Effects of Matrix Stiffness on the Morphology, Adhesion, Proliferation and Osteogenic Differentiation of Mesenchymal Stem Cells. Int. J. Med. Sci. 2018, 15, 257–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roach, P.; Parker, T.; Gadegaard, N.; Alexander, M.R. A bio-inspired neural environment to control neurons comprising radial glia, substrate chemistry and topography. Biomater. Sci. 2012, 1, 83–93. [Google Scholar] [CrossRef]
- Coyle, S.; Doss, B.; Huo, Y.; Singh, H.R.; Quinn, D.; Hsia, K.J.; LeDuc, P.R. Cell alignment modulated by surface nano-topography—Roles of cell-matrix and cell-cell interactions. Acta Biomater. 2022, 142, 149–159. [Google Scholar] [CrossRef]
- Lou, H.-Y.; Zhao, W.; Li, X.; Duan, L.; Powers, A.; Akamatsu, M.; Santoro, F.; McGuire, A.F.; Cui, Y.; Drubin, D.G.; et al. Membrane curvature underlies actin reorganization in response to nanoscale surface topography. Proc. Natl. Acad. Sci. USA 2019, 116, 23143–23151. [Google Scholar] [CrossRef]
- Schaumann, E.N.; Tian, B. Actin-packed topography: Cytoskeletal response to curvature. Proc. Natl. Acad. Sci. USA 2019, 116, 22897–22898. [Google Scholar] [CrossRef]
- Mathieu, P.S.; Loboa, E.G. Cytoskeletal and Focal Adhesion Influences on Mesenchymal Stem Cell Shape, Mechanical Properties, and Differentiation Down Osteogenic, Adipogenic, and Chondrogenic Pathways. Tissue Eng. Part B Rev. 2012, 18, 436–444. [Google Scholar] [CrossRef] [Green Version]
- Richards, D.; Swift, J.; Wong, L.S.; Richardson, S.M. Photoresponsive Hydrogels with Photoswitchable Stiffness: Emerging Platforms to Study Temporal Aspects of Mesenchymal Stem Cell Responses to Extracellular Stiffness Regulation. Adv. Exp. Med. Biol. 2018, 5, 53–69. [Google Scholar] [CrossRef]
- Fabiilli, M.L.; Wilson, C.G.; Padilla, F.; Martín-Saavedra, F.M.; Fowlkes, J.B.; Franceschi, R.T. Acoustic droplet–hydrogel composites for spatial and temporal control of growth factor delivery and scaffold stiffness. Acta Biomater. 2013, 9, 7399–7409. [Google Scholar] [CrossRef]
- Pan, H.-A.; Liang, J.-Y.; Hung, Y.-C.; Lee, C.-H.; Chiou, J.-C.; Huang, G.S. The spatial and temporal control of cell migration by na-noporous surfaces through the regulation of ERK and integrins in fibroblasts. Biomaterials 2013, 34, 841–853. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.-N.; Dobre, O.; Richards, D.; Ballestrem, C.; Curran, J.M.; Hunt, J.A.; Richardson, S.; Swift, J.; Wong, L.S. Photoresponsive Hydrogels with Photoswitchable Mechanical Properties Allow Time-Resolved Analysis of Cellular Responses to Matrix Stiffening. ACS Appl. Mater. Interfaces 2018, 10, 7765–7776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trujillo, V.; Kim, J.; Hayward, R.C. Creasing instability of surface-attached hydrogels. Soft Matter 2008, 4, 564–569. [Google Scholar] [CrossRef] [PubMed]
- James, S.; Fox, J.; Afsari, F.; Lee, J.; Clough, S.; Knight, C.; Ashmore, J.; Ashton, P.; Preham, O.; Hoogduijn, M.; et al. Multiparameter Analysis of Human Bone Marrow Stromal Cells Identifies Distinct Immunomodulatory and Differentiation-Competent Subtypes. Stem Cell Rep. 2015, 4, 1004–1015. [Google Scholar] [CrossRef] [Green Version]
- Galarza Torre, A.; Shaw, J.E.; Wood, A.; Gilbert, H.T.; Dobre, O.; Genever, P.; Brennan, K.; Richardson, S.M.; Swift, J. An immortalised mesenchymal stem cell line maintains mechano-responsive behaviour and can be used as a reporter of substrate stiffness. Sci. Rep. 2018, 8, 8981. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.-K.; Wang, M.-J.J. Image thresholding by minimizing the measures of fuzziness. Pattern Recognit. 1995, 28, 41–51. [Google Scholar] [CrossRef]
- Reversat, A.; Gaertner, F.; Merrin, J.; Stopp, J.; Tasciyan, S.; Aguilera, J.; de Vries, I.; Hauschild, R.; Hons, M.; Piel, M.; et al. Cellular locomotion using environmental topography. Nature 2020, 582, 582–585. [Google Scholar] [CrossRef]
- Brunette, D.M.; Chehroudi, B. The Effects of the Surface Topography of Micromachined Titanium Substrata on Cell Behavior in Vitro and in Vivo. J. Biomech. Eng. 1999, 121, 49–57. [Google Scholar] [CrossRef]
- Anselme, K.; Ploux, L.; Ponche, A. Cell/Material Interfaces: Influence of Surface Chemistry and Surface Topography on Cell Adhesion. J. Adhes. Sci. Technol. 2010, 24, 831–852. [Google Scholar] [CrossRef]
- Czeisler, C.; Short, A.; Nelson, T.; Gygli, P.; Ortiz, C.; Catacutan, F.P.; Stocker, B.; Cronin, J.; Lannutti, J.; Winter, J.; et al. Surface topography during neural stem cell differentiation regulates cell migration and cell morphology. J. Comp. Neurol. 2016, 524, 3485–3502. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, X.; Liu, Y.; Wang, D.; Yu, X.; Wang, H.; Bai, Z.; Jiang, Y.-C.; Li, X.; Zheng, W.; et al. Endothelial Cell Migration Regulated by Surface Topography of Poly(ε-caprolactone) Nanofibers. ACS Biomater. Sci. Eng. 2021, 7, 4959–4970. [Google Scholar] [CrossRef] [PubMed]
- Awsiuk, K.; Stetsyshyn, Y.; Raczkowska, J.; Lishchynskyi, O.; Dąbczyński, P.; Kostruba, A.; Ohar, H.; Shymborska, Y.; Nastyshyn, S.; Budkowski, A. Temperature-controlled ori-entation of proteins on temperature-responsive grafted polymer brushes: Poly (butyl methacrylate) vs poly (butyl acrylate): Morphology, wetting, and protein adsorption. Biomacromolecules 2019, 20, 2185–2197. [Google Scholar] [CrossRef] [PubMed]
- Sandmann, R.; Köster, S. Topographic Cues Reveal Two Distinct Spreading Mechanisms in Blood Platelets. Sci. Rep. 2016, 6, 22357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, H.; Gong, B.; Yin, J.; Qian, J. 3D printing topographic cues for cell contact guidance: A review. Mater. Des. 2022, 218, 110663. [Google Scholar] [CrossRef]
- Li, X.; Zhang, Q.; Yan, S.; Li, M.; You, R. Topographic cues reveal filopodia-mediated cell locomotion in 3D microenvironment. Biointerphases 2020, 15, 031001. [Google Scholar] [CrossRef]
- Song, K.H.; Kwon, K.W.; Choi, J.-C.; Jung, J.; Park, Y.; Suh, K.-Y.; Doh, J. T cells sense biophysical cues using lamellipodia and filopodia to optimize intraluminal path finding. Integr. Biol. 2014, 6, 450–459. [Google Scholar] [CrossRef] [Green Version]
- de Silva, M.S.; Alvarado, J.; Nguyen, J.; Georgoulia, N.; Mulder, B.M.; Koenderink, G.H. Self-organized patterns of actin filaments in cell-sized confinement. Soft Matter 2011, 7, 10631–10641. [Google Scholar] [CrossRef]
- Bade, N.D.; Kamien, R.D.; Assoian, R.K.; Stebe, K.J. Edges impose planar alignment in nematic monolayers by directing cell elongation and enhancing migration. Soft Matter 2018, 14, 6867–6874. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moxon, S.R.; Richards, D.; Dobre, O.; Wong, L.S.; Swift, J.; Richardson, S.M. Regulation of Mesenchymal Stem Cell Morphology Using Hydrogel Substrates with Tunable Topography and Photoswitchable Stiffness. Polymers 2022, 14, 5338. https://doi.org/10.3390/polym14245338
Moxon SR, Richards D, Dobre O, Wong LS, Swift J, Richardson SM. Regulation of Mesenchymal Stem Cell Morphology Using Hydrogel Substrates with Tunable Topography and Photoswitchable Stiffness. Polymers. 2022; 14(24):5338. https://doi.org/10.3390/polym14245338
Chicago/Turabian StyleMoxon, Samuel R., David Richards, Oana Dobre, Lu Shin Wong, Joe Swift, and Stephen M. Richardson. 2022. "Regulation of Mesenchymal Stem Cell Morphology Using Hydrogel Substrates with Tunable Topography and Photoswitchable Stiffness" Polymers 14, no. 24: 5338. https://doi.org/10.3390/polym14245338