
Differences in the Elastomeric Behavior of Polyglycine-Rich Regions of Spidroin 1 and 2 Proteins
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sequence Selection
2.2. Molecular Dynamics (MD) Calculations
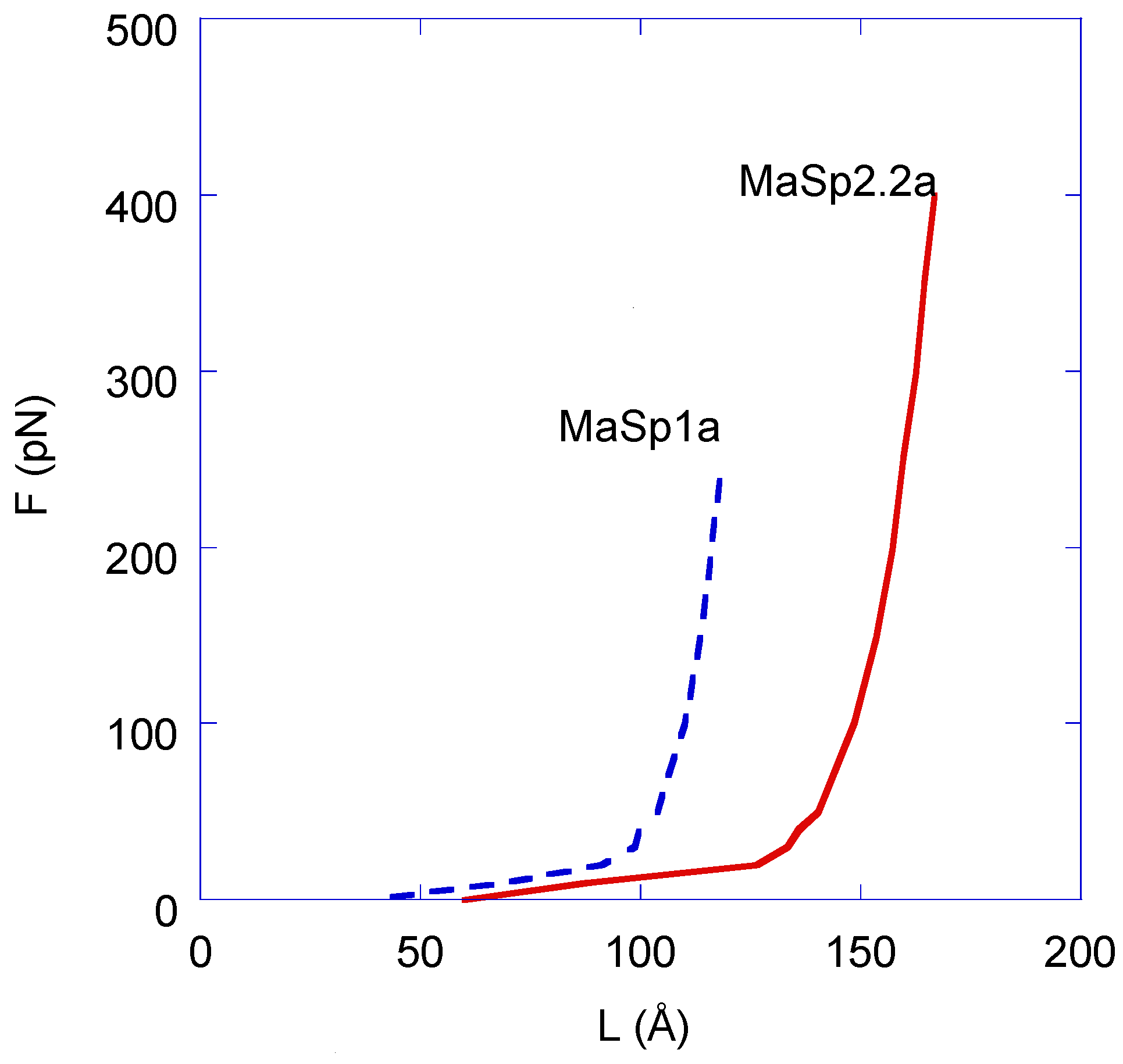
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heim, M.; Keerl, D.; Scheibel, T. Spider silk: From soluble protein to extraordinary fiber. Angew. Chem. Int. Ed. Eng. 2009, 48, 3584–3596. [Google Scholar] [CrossRef] [PubMed]
- Vepari, C.; Kaplan, D. Silk as a biomaterial. Prog. Polym. Sci. 2007, 32, 991–1007. [Google Scholar] [CrossRef] [PubMed]
- Agnarsson, I.; Kuntner, M.; Blackledge, T.A. Bioprospecting Finds the Toughest Biological Material: Extraordinary Silk from a Giant Riverine Orb Spider. PLoS ONE 2010, 5, e11234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, A.; Kim, K.; Shamble, P.S. Rapid mid-jump production of high-performance silk by jumping spiders. Curr. Biol. 2021, 31, R1422–R1423. [Google Scholar] [CrossRef]
- Blackledge, T.A.; Pérez-Rigueiro, J.; Plaza, G.R.; Perea, B.; Navarro, A.; Guinea, G.V.; Elices, M. Sequential origin in the high performance properties of orb spider dragline silk. Sci. Rep. 2012, 2, 782. [Google Scholar] [CrossRef] [Green Version]
- Keten, S.; Xu, Z.P.; Ihle, B.; Buehler, M.J. Nanoconfinement controls stiffness, strength and mechanical toughness of beta-sheet crystals in silk. Nat. Mater. 2010, 9, 359–367. [Google Scholar] [CrossRef]
- Selden, P.A.; Shear, W.A.; Sutton, M.D. Fossil evidence for the origin of spider spinnerets, and a proposed arachnid order. Proc. Natl. Acad. Sci. USA 2008, 105, 20781–20785. [Google Scholar] [CrossRef] [Green Version]
- Blackledge, T.A.; Hayashi, C.Y. Silken toolkits: Biomechanics of silk fibers spun by the orb web spider Argiope argentata (Fabricius 1775). J. Exp. Biol. 2006, 209, 2452–2461. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Lewis, R.V. Structure of a Protein Superfiber—Spider Dragline Silk. Proc. Natl. Acad. Sci. USA 1990, 87, 7120–7124. [Google Scholar] [CrossRef] [Green Version]
- Blackledge, T.A.; Scharff, N.; Coddington, J.A.; Szuts, T.; Wenzel, J.W.; Hayashi, C.Y.; Agnarsson, I. Reconstructing web evolution and spider diversification in the molecular era. Proc. Natl. Acad. Sci. USA 2009, 106, 5229–5234. [Google Scholar] [CrossRef]
- Hagn, F.; Eisoldt, L.; Hardy, J.G.; Vendrely, C.; Coles, M.; Scheibel, T.; Kessler, H. A conserved spider silk domain acts as a molecular switch that controls fibre assembly. Nature 2010, 465, 239–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Askarieh, G.; Hedhammar, M.; Nordling, K.; Saenz, A.; Casals, C.; Rising, A.; Johansson, J.; Knight, S.D. Self-assembly of spider silk proteins is controlled by a pH-sensitive relay. Nature 2010, 465, 236–239. [Google Scholar] [CrossRef]
- Gatesy, J.; Hayashi, C.; Motriuk, D.; Woods, J.; Lewis, R. Extreme diversity, conservation, and convergence of spider silk fibroin sequences. Science 2001, 291, 2603–2605. [Google Scholar] [CrossRef] [PubMed]
- Starrett, J.; Garb, J.E.; Kuelbs, A.; Azubuike, U.O.; Hayashi, C.Y. Early Events in the Evolution of Spider Silk Genes. PLoS ONE 2012, 7, e38084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauscher, S.; Baud, S.; Miao, M.; Keeley, F.W.; Pomes, R. Proline and glycine control protein self-organization into elastomeric or amyloid fibrils. Structure 2006, 14, 1667–1676. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Sponner, A.; Porter, D.; Vollrath, F. Proline and processing of spider silks. Biomacromolecules 2008, 9, 116–121. [Google Scholar] [CrossRef]
- Dunaway, D.L.; Thiel, B.L.; Viney, C. Tensile Mechanical Property Evaluation of Natural and Epoxide-Treated Silk Fibers. J. Appl. Polym. Sci. 1995, 58, 675–683. [Google Scholar] [CrossRef]
- Madsen, B.; Shao, Z.Z.; Vollrath, F. Variability in the mechanical properties of spider silks on three levels: Interspecific, intraspecific and intraindividual. Int. J. Biol. Macromol. 1999, 24, 301–306. [Google Scholar] [CrossRef]
- Babb, P.L.; Lahens, N.F.; Correa-Garhwal, S.M.; Nicholson, D.N.; Kim, E.J.; Hogenesch, J.B.; Kuntner, M.; Higgins, L.; Hayashi, C.Y.; Agnarsson, I.; et al. The Nephila clavipes genome highlights the diversity of spider silk genes and their complex expression. Nat. Genet. 2017, 49, 895–903. [Google Scholar] [CrossRef] [Green Version]
- Kono, N.; Nakamura, H.; Ohtoshi, R.; Moran, D.A.P.E.; Shinohara, A.; Yoshida, Y.; Fujiwara, M.; Mori, M.; Tomita, M.; Arakawa, K. Orb-weaving spider Araneus ventricosus genome elucidates the spidroin gene catalogue. Sci. Rep. 2019, 9, 8380. [Google Scholar] [CrossRef]
- Madurga, R.; Plaza, G.R.; Blackledge, T.A.; Guinea, G.V.; Elices, M.; Perez-Rigueiro, J. Material properties of evolutionary diverse spider silks described by variation in a single structural parameter. Sci. Rep. 2016, 6, 18991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrote, J.; Ruiz, V.; Troncoso, O.P.; Torres, F.G.; Arnedo, M.; Elices, M.; Guinea, G.V.; Perez-Rigueiro, J. Application of the Spider Silk Standardization Initiative ((SI)-I-3) methodology to the characterization of major ampullate gland silk fibers spun by spiders from Pantanos de Villa wetlands (Lima, Peru). J. Mech. Behav. Biomed. Mater. 2020, 111, 104023. [Google Scholar] [CrossRef] [PubMed]
- Arguelles, J.; Baker, R.H.; Pérez-Rigueiro, J.; Guinea, G.V.; Elices, M.; Hayashi, C.Y. Relating spidroin motif prevalence and periodicity to the mechanical properties of major ampullate spider silks. J. Comp. Physiol. B 2022. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, D.; Tokareva, O.; Rim, N.G.; Wong, J.Y.; Kaplan, D.L.; Buehler, M.J. Silk-Its Mysteries, How It Is Made, and How It Is Used. ACS Biomater. Sci. Eng. 2015, 1, 864–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollingsworth, S.A.; Dror, R.O. Molecular Dynamics Simulation for All. Neuron 2018, 99, 1129–1143. [Google Scholar] [CrossRef] [Green Version]
- Keten, S.; Buehler, M.J. Nanostructure and molecular mechanics of spider dragline silk protein assemblies. J. R. Soc. Interface 2010, 7, 1709–1721. [Google Scholar] [CrossRef] [Green Version]
- Asakura, T. Structure and Dynamics of Spider Silk Studied with Solid-State Nuclear Magnetic Resonance and Molecular Dynamics Simulation. Molecules 2020, 25, 2634. [Google Scholar] [CrossRef]
- Buehler, M.J. Tu(r)ning weakness to strength. Nano Today 2010, 5, 379–383. [Google Scholar] [CrossRef]
- Bratzel, G.; Buehler, M.J. Sequence-structure correlations in silk: Poly-Ala repeat of N. clavipes MaSp1 is naturally optimized at a critical length scale. J. Mech. Behav. Biomed. Mater. 2012, 7, 30–40. [Google Scholar] [CrossRef]
- Cetinkaya, M.; Xiao, S.; Graeter, F. Effects of crystalline subunit size on silk fiber mechanics. Soft Matter 2011, 7, 8142–8148. [Google Scholar] [CrossRef]
- Xiao, S.; Stacklies, W.; Cetinkaya, M.; Markert, B.; Graeter, F. Mechanical Response of Silk Crystalline Units from Force-Distribution Analysis. Biophys. J. 2009, 96, 3997–4005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, J.A.; Patil, S.P.; Greving, I.; Laemmel, M.; Gkagkas, K.; Seydel, T.; Mueller, M.; Markert, B.; Graeter, F. Stress-induced long-range ordering in spider silk. Sci. Rep. 2017, 7, 15273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, J.C.; Hardy, D.J.; Maia, J.D.C.; Stone, J.E.; Ribeiro, J.V.; Bernardi, R.C.; Buch, R.; Fiorin, G.; Henin, J.; Jiang, W.; et al. Scalable molecular dynamics on CPU and GPU architectures with NAMD. J. Chem. Phys. 2020, 153, 044130. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Rauscher, S.; Nawrocki, G.; Ran, T.; Feig, M.; de Groot, B.L.; Grubmueller, H.; MacKerell, A.D., Jr. CHARMM36: An Improved Force Field for Folded and Intrinsically Disordered Proteins. Biol. J. 2017, 112, 175A–176A. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. Model. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Termonia, Y. Molecular Modelling of the Stress/Strainbehaviour of Spider Dragline; Elices, M., Ed.; Pergamon Press: Amsterdam, The Netherlands, 2000; pp. 335–349. [Google Scholar]
- Termonia, Y. Molecular Modeling of Spider Silk Elasticity. Macromolecules 1994, 27, 7378–7381. [Google Scholar] [CrossRef]
- Strobl, G. The Physics of Polymers; Springer: Berlin, Germany, 1996. [Google Scholar]
- Work, R.W. Dimensions, Birefringences, and Force-Elongation Behavior of Major and Minor Ampullate Silk Fibers from Orb-Web-Spinning Spiders—Effects of Wetting on these Properties. Text Res. J. 1977, 47, 650–662. [Google Scholar] [CrossRef]
- Work, R.W. A Comparative-Study of the Super-Contraction of Major Ampullate Silk Fibers of Orb-Web-Building Spiders (Araneae). J. Arachnol. 1981, 9, 299–308. [Google Scholar]
- Work, R.W.; Morosoff, N. A Physicochemical Study of the Supercontraction of Spider Major Ampullate Silk Fibers. Text Res. J. 1982, 52, 349–356. [Google Scholar] [CrossRef]
- Gosline, J.M.; Denny, M.W.; Demont, M.E. Spider Silk as Rubber. Nature 1984, 309, 551–552. [Google Scholar] [CrossRef]
- Gosline, J.M.; Pollak, C.C.; Guerette, P.A.; Cheng, A.; Demont, M.E.; Denny, M.W. Elastomeric Network Models for the Frame and Viscid Silks from the Orb Web of the Spider Araneus diadematus. Silk Polym. 1994, 544, 328–341. [Google Scholar]
- Riekel, C.; Branden, C.; Craig, C.; Ferrero, C.; Heidelbach, F.; Muller, M. Aspects of X-ray diffraction on single spider fibers. Int. J. Biol. Macromol. 1999, 24, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Riekel, C. The Mechanical Functions of Silks and Their Correlated Structural Properties; Craig, C.L., Ed.; Oxford University Press: Oxford, UK, 2003; pp. 51–82. [Google Scholar]
- Perez-Rigueiro, J.; Elices, M.; Plaza, G.R.; Guinea, G.V. Basic Principles in the Design of Spider Silk Fibers. Molecules 2021, 26, 1794. [Google Scholar] [CrossRef] [PubMed]
- Vollrath, F. Spider Webs and Silks. Sci. Am. 1992, 266, 70–76. [Google Scholar]
- Shao, Z.Z.; Vollrath, F. The effect of solvents on the contraction and mechanical properties of spider silk. Polymer 1999, 40, 1799–1806. [Google Scholar] [CrossRef]
- Savage, K.N.; Gosline, J.M. The effect of proline on the network structure of major ampullate silks as inferred from their mechanical and optical properties. J. Exp. Biol. 2008, 211, 1937–1947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Rigueiro, J.; Plaza, G.R.; Torres, F.G.; Hijar, A.; Hayashi, C.; Perea, G.B.; Elices, M.; Guinea, G.V. Supercontraction of dragline silk spun by lynx spiders (Oxyopidae). Int. J. Biol. Macromol. 2010, 46, 555–557. [Google Scholar] [CrossRef]
- Boutry, C.; Blackledge, T.A. Evolution of supercontraction in spider silk: Structure-function relationship from tarantulas to orb-weavers. J. Exp. Biol. 2010, 213, 3505–3514. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.T.; Liivak, O.; Seidel, A.; LaVerde, G.; Zax, D.B.; Jelinski, L.W. Supercontraction and backbone dynamics in spider silk: C-13 and H-2 NMR studies. J. Am. Chem. Soc. 2000, 122, 9019–9025. [Google Scholar] [CrossRef]
- Holland, G.P.; Creager, M.S.; Jenkins, J.E.; Lewis, R.V.; Yarger, J.L. Determining secondary structure in spider dragline silk by carbon-carbon correlation solid-state NMR spectroscopy. J. Am. Chem. Soc. 2008, 130, 9871–9877. [Google Scholar] [CrossRef]
- Jenkins, J.E.; Creager, M.S.; Butler, E.B.; Lewis, R.V.; Yarger, J.L.; Holland, G.P. Solid-state NMR evidence for elastin-like beta-turn structure in spider dragline silk. Chem. Commun. 2010, 46, 6714–6716. [Google Scholar] [CrossRef]
- Eles, P.T.; Michal, C.A. Strain dependent local phase transitions observed during controlled supercontraction reveal mechanisms in spider silk. Macromolecules 2004, 37, 1342–1345. [Google Scholar]
- Guan, J.; Vollrath, F.; Porter, D. Two Mechanisms for Supercontraction in Nephila Spider Dragline Silk. Biomacromolecules 2011, 12, 4030–4035. [Google Scholar] [CrossRef] [PubMed]
- Greco, G.; Arndt, T.; Schmuck, B.; Francis, J.; Backlund, F.G.; Shilkova, O.; Barth, A.; Gonska, N.; Seisenbaeva, G.; Kessler, V.; et al. Tyrosine residues mediate supercontraction in biomimetic spider silk. Commun. Mater. 2021, 2, 43. [Google Scholar] [CrossRef]
- Tokareva, O.; Jacobsen, M.; Buehler, M.; Wong, J.; Kaplan, D.L. Structure-function-property-design interplay in biopolymers: Spider silk. Acta Biomater. 2014, 10, 1612–1626. [Google Scholar] [CrossRef] [Green Version]
- Filipe, H.A.L.; Loura, L.M.S. Molecular Dynamics Simulations: Advances and Applications. Molecules 2022, 27, 2105. [Google Scholar] [CrossRef]
- Patil, S.P.; Xiao, S.; Gkagkas, K.; Markert, B.; Graeter, F. Viscous Friction between Crystalline and Amorphous Phase of Dragline Silk. PLoS ONE 2014, 9, e104832. [Google Scholar] [CrossRef] [Green Version]
- Patil, S.P.; Markert, B.; Graeter, F. Rate-Dependent Behavior of the Amorphous Phase of Spider Dragline Silk. Biophys. J. 2014, 106, 2511–2518. [Google Scholar] [CrossRef]
- Belen Perea, G.; Solanas, C.; Plaza, G.R.; Guinea, G.V.; Jorge, I.; Vazquez, J.; Perez Mateos, J.M.; Mari-Buye, N.; Elices, M.; Perez-Rigueiro, J. Unexpected behavior of irradiated spider silk links conformational freedom to mechanical performance. Soft Matter 2015, 11, 4868–4878. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MaSp1a | MaSp2.2a | |||
|---|---|---|---|---|
| Average (Å) | Std. Deviation (Å) | Average (Å) | Std. Deviation (Å) | |
| Trajectory 1 | 40.1 | 14.6 | 52.3 | 27.6 |
| Trajectory 2 | 33.4 | 20.5 | 60.1 | 22.0 |
| Trajectory 3 | 43.9 | 15.1 | 68.8 | 24.8 |
| Mean | 39 ± 3 Å | 60 ± 5 Å | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pacios, L.F.; Arguelles, J.; Hayashi, C.Y.; Guinea, G.V.; Elices, M.; Perez-Rigueiro, J. Differences in the Elastomeric Behavior of Polyglycine-Rich Regions of Spidroin 1 and 2 Proteins. Polymers 2022, 14, 5263. https://doi.org/10.3390/polym14235263
Pacios LF, Arguelles J, Hayashi CY, Guinea GV, Elices M, Perez-Rigueiro J. Differences in the Elastomeric Behavior of Polyglycine-Rich Regions of Spidroin 1 and 2 Proteins. Polymers. 2022; 14(23):5263. https://doi.org/10.3390/polym14235263
Chicago/Turabian StylePacios, Luis F., Joseph Arguelles, Cheryl Y. Hayashi, Gustavo V. Guinea, Manuel Elices, and Jose Perez-Rigueiro. 2022. "Differences in the Elastomeric Behavior of Polyglycine-Rich Regions of Spidroin 1 and 2 Proteins" Polymers 14, no. 23: 5263. https://doi.org/10.3390/polym14235263