
Mass Transport of Lignin in Confined Pores

 , , , and
, , , and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
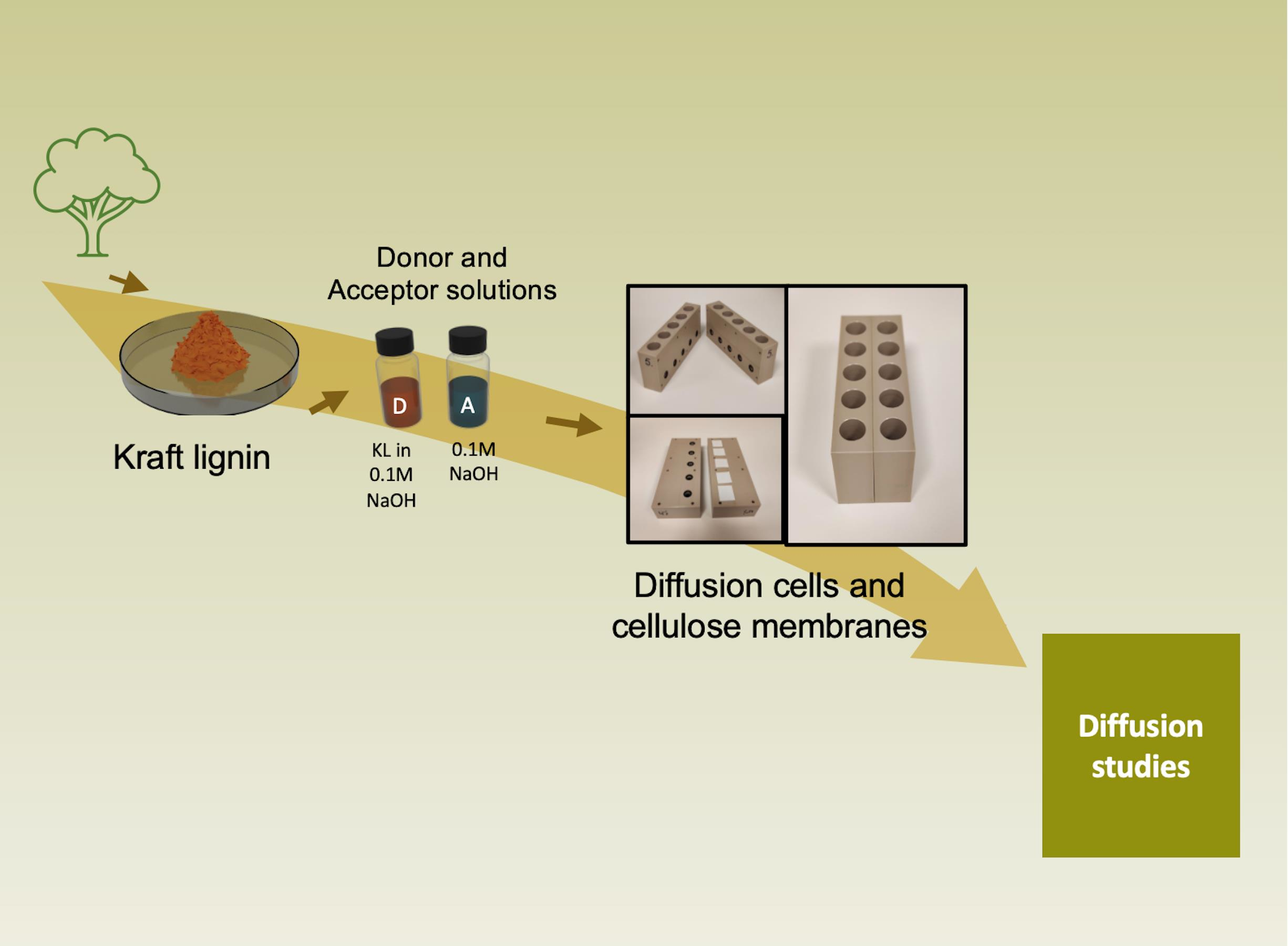
2.2. Diffusion Experiments: The Set-Up of the Diffusion Cells
2.3. Diffusion Experiments: The Procedure for Lignin Diffusion
2.4. Size-Exclusion Chromatography
2.5. Ultra-High Performance Liquid Chromatography (UHPLC)
2.6. UV–Vis Absorbance Measurements
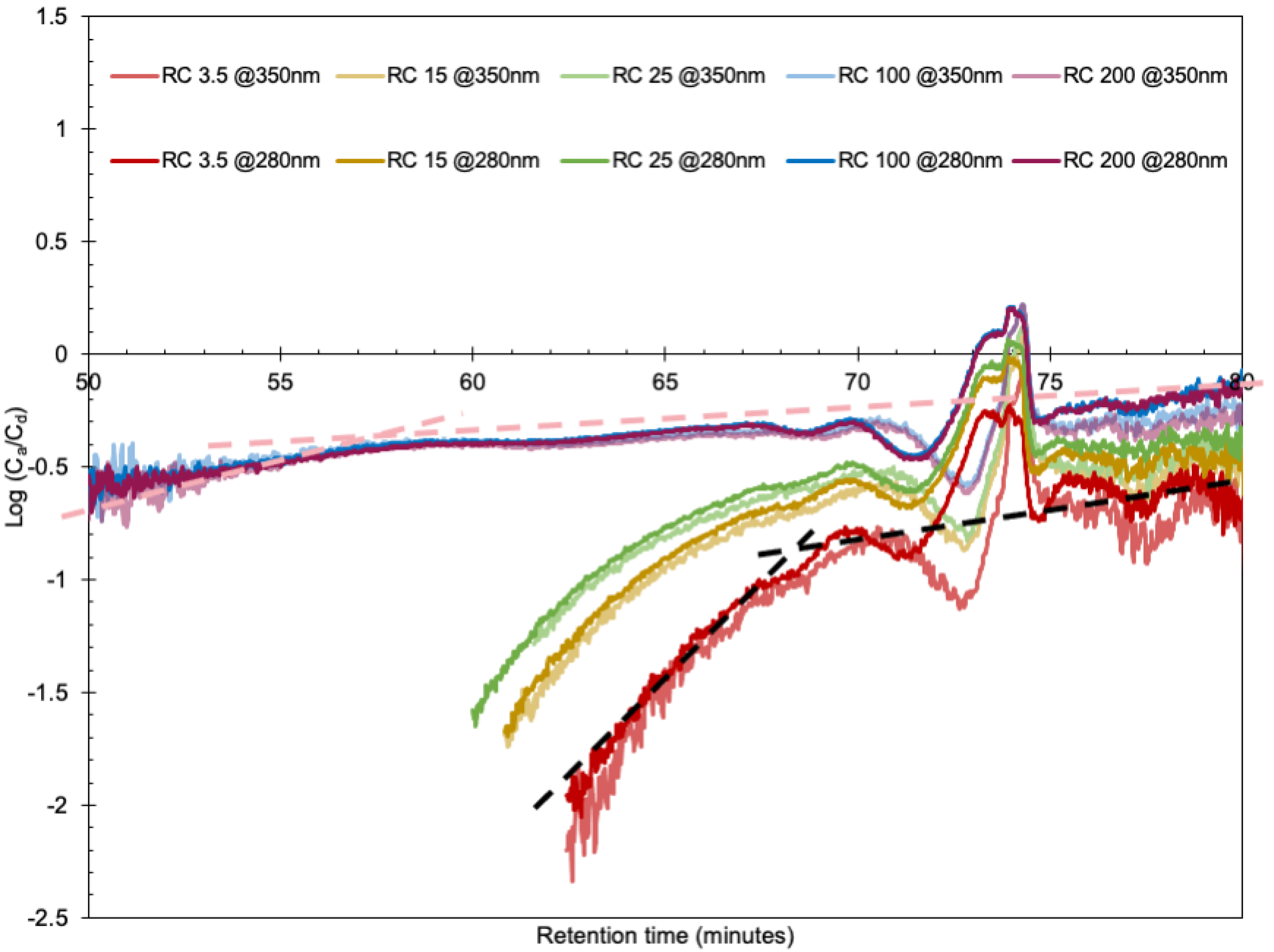
3. Results
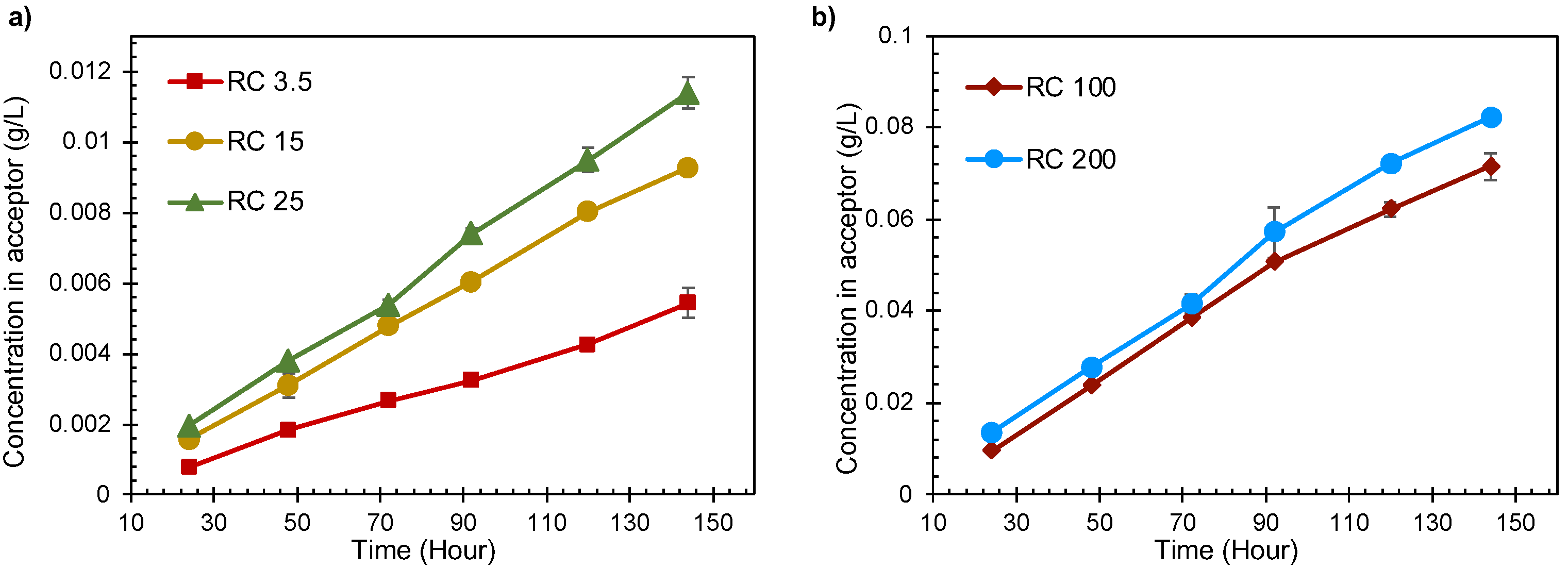
3.1. Transport through Membranes in Diffusion Cells
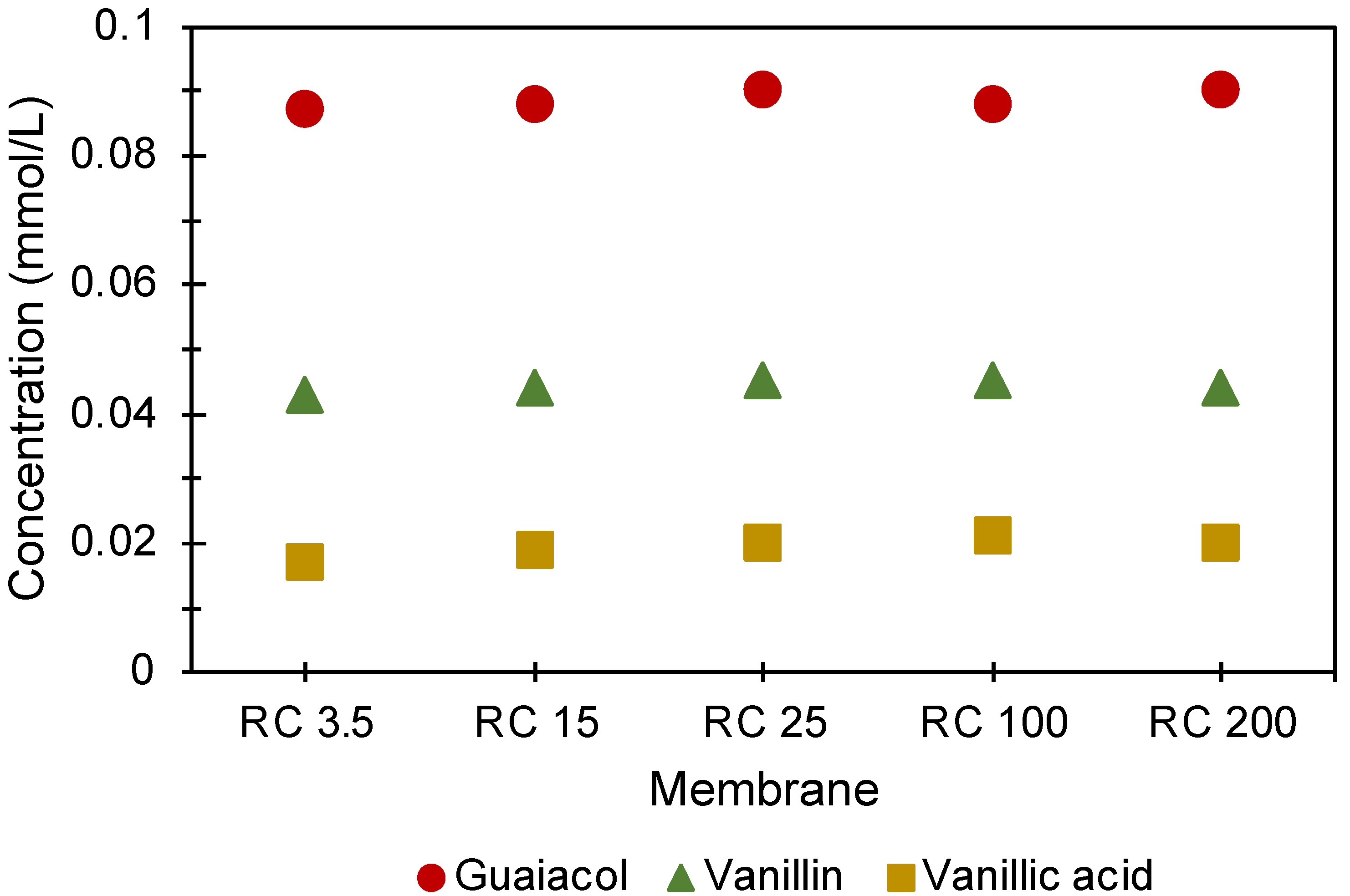
3.2. Properties of the Lignin Transported through Membranes in Diffusion Cells
- The amount of lignin transported from the donor side to the acceptor side increased with time as the pore size of the intermediate membrane increases.
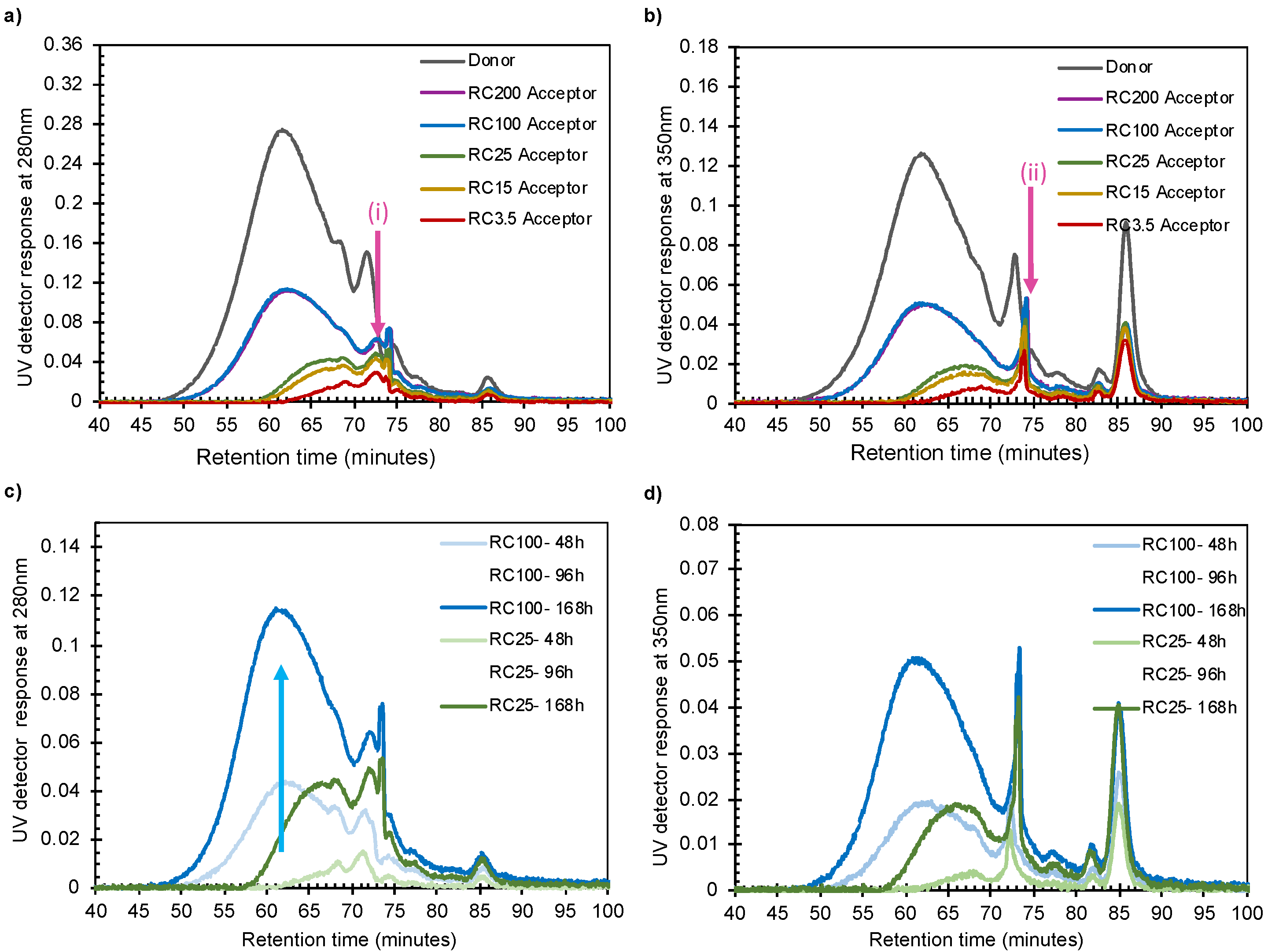
- The absorbance intensity in the SEC increased with time and increasing pore size, even for small fragments.
- The intensity in the SEC chromatogram started to increase at lignin sizes that correspond to a factor of almost 10 times smaller than the nominal pore sizes.
- Corresponding SEC curves recorded at 280 and 350 nm were not identical, especially for the smaller lignin molecules.
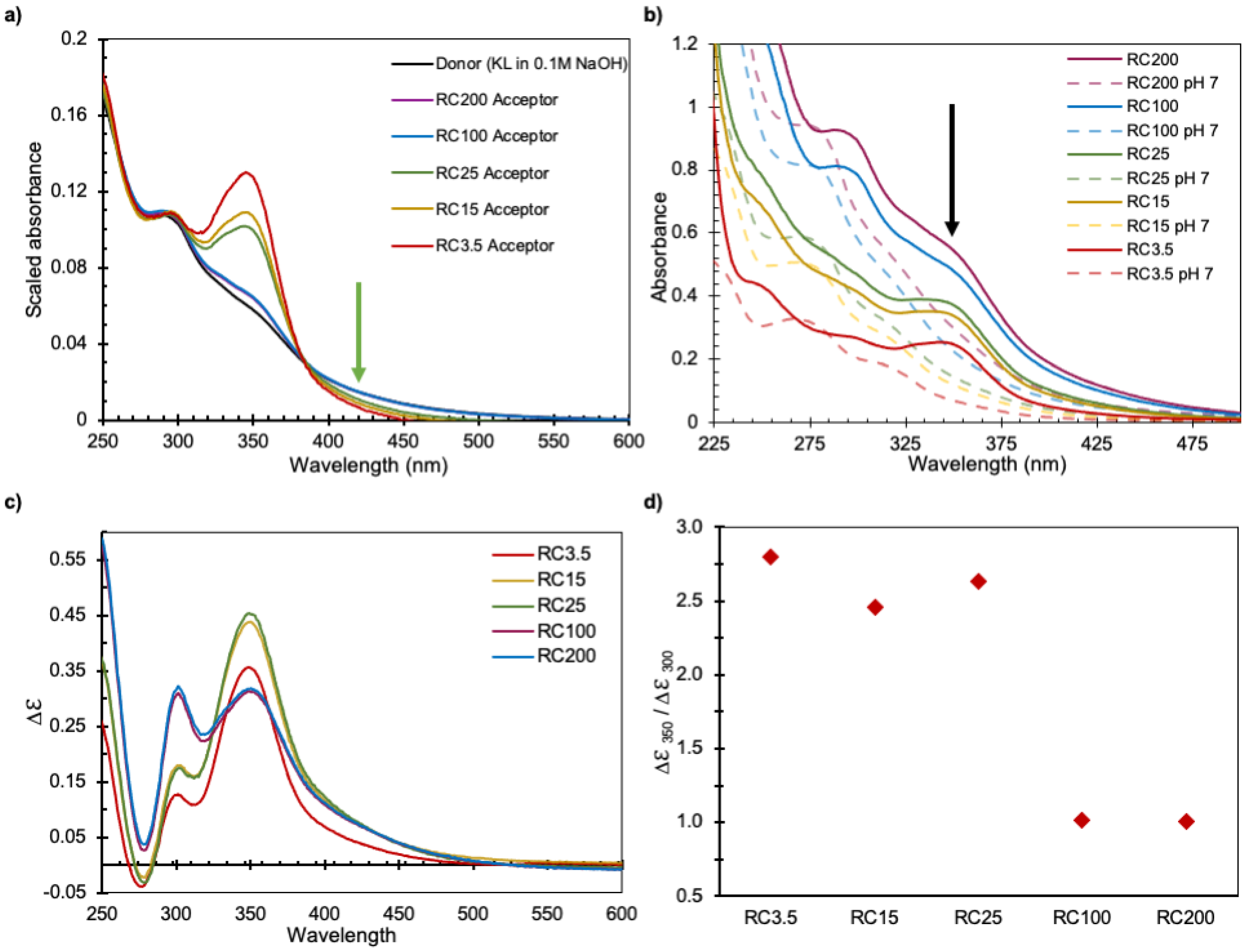
- UV–vis absorbance spectra from the acceptor cells of the different membranes were not identical.
3.2.1. Chemical Differences of the Lignin Transported through Membranes in Diffusion Cells
3.2.2. Size-Fraction of Lignin Transported through Membranes in Diffusion Cells
3.3. A Critical Lignin Size Controls the Transport Mechanism
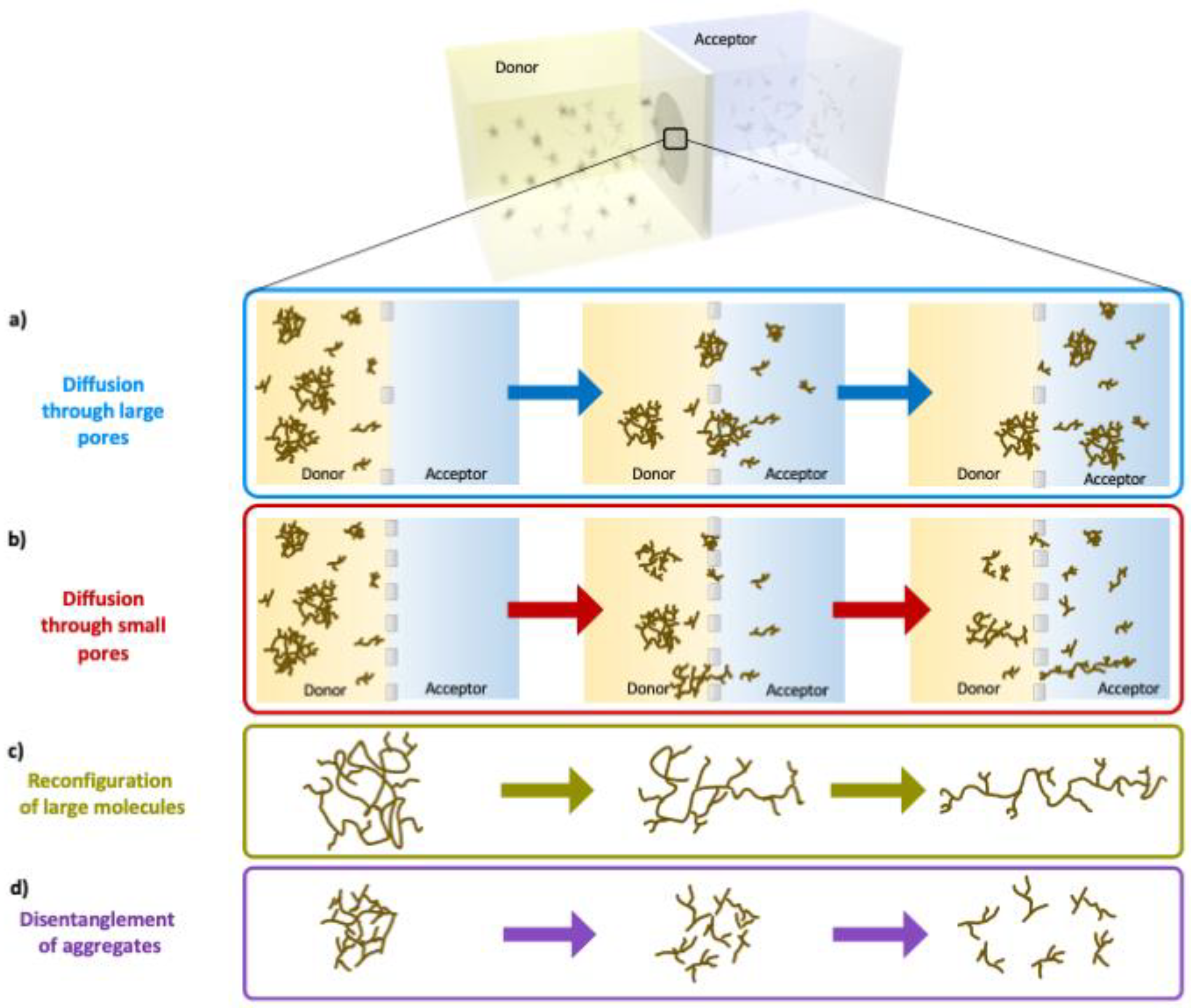
4. Discussion
4.1. Transport Mechanism of Lignin in Porous Environments
4.2. Effects on the Delignification Process
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.; Wang, H.; Sun, X.; Wang, Y.; Liu, Z. Separation and Characterization of Biomass Components (Cellulose, Hemicellulose, and Lignin) from Corn Stalk. BioResources 2021, 16, 7205–7219. [Google Scholar] [CrossRef]
- Sixta, H. Pulp Bleaching, Handbook of the Pulp; Wiley VCH; Verlag GmbH &Co. KGaA: Weinheim, Germany, 2006; ISBN 3-527-30999-3. [Google Scholar]
- Alén, R. Pulp Mills and Wood-Based Biorefineries; Elsevier B.V.: Amsterdam, The Netherlands, 2015. [Google Scholar] [CrossRef]
- Li, J.; Mui, C. Effect of lignin diffusion on kraft delignification kinetics as determined by liquor analysis. Part I: An experimental study. J. Pulp Pap. Sci. 1999, 25, 373–377. [Google Scholar]
- Simão, J.P.F.; Carvalho, M.G.V.S.; Baptista, C.M.S.G. Heterogeneous studies in pulping of wood: Modelling mass transfer of dissolved lignin. Chem. Eng. J. 2011, 170, 264–269. [Google Scholar] [CrossRef]
- Kawamata, Y.; Yoshikawa, T.; Aoki, H.; Koyama, Y.; Nakasaka, Y.; Yoshida, M.; Masuda, T. Kinetic analysis of delignification of cedar wood during organosolv treatment with a two-phase solvent using the unreacted-core model. Chem. Eng. J. 2019, 368, 71–78. [Google Scholar] [CrossRef]
- Jacobson, A. Diffusion of Chemicals into Green Wood; Georgia Institute of Technology: Atlanta, GA, USA, 2006. [Google Scholar]
- Mattsson, C.; Hasani, M.; Dang, B.; Mayzel, M.; Theliander, H. About structural changes of lignin during kraft cooking and the kinetics of delignification. Holzforschung 2017, 71, 545–553. [Google Scholar] [CrossRef]
- Li, J.; Phoenix, A.; Macleod, J.M. Diffusion of Lignin Macromolecules within the Fibre Walls of Kraft Pulp. Part I: Determination of the Diffusion Coefficient under Alkaline Conditions. Can. J. Chem. Eng. 1997, 75, 16–22. [Google Scholar] [CrossRef]
- Brännvall, E.; Rönnols, J. Analysis of entrapped and free liquor to gain new insights into kraft pulping. Cellulose 2021, 28, 2403–2418. [Google Scholar] [CrossRef]
- Guo, Y. Hindered Diffusion of Polymers in Porous Materials; University of Massachusetts Amherst: Amherst, MA, USA, 1991. [Google Scholar]
- Gårdebjer, S.; Bergstrand, A.; Idström, A.; Börstell, C.; Naana, S.; Nordstierna, L.; Larsson, A. Solid-state NMR to quantify surface coverage and chain length of lactic acid modified cellulose nanocrystals, used as fillers in biodegradable composites. Compos. Sci. Technol. 2015, 107, 1–9. [Google Scholar] [CrossRef]
- Andersson, H.; Hjärtstam, J.; Stading, M.; von Corswant, C.; Larsson, A. Effects of molecular weight on permeability and microstructure of mixed ethyl-hydroxypropyl-cellulose films. Eur. J. Pharm. Sci. 2013, 48, 240–248. [Google Scholar] [CrossRef]
- Gårdebjer, S.; Larsson, M.; Gebäck, T.; Skepö, M.; Larsson, A. An overview of the transport of liquid molecules through structured polymer films, barriers and composites–Experiments correlated to structure-based simulations. Adv. Colloid Interface Sci. 2018, 256, 48–64. [Google Scholar] [CrossRef]
- Bergstrand, A.; Andersson, H.; Cramby, J.; Sott, K.; Larsson, A. Preparation of Porous Poly(3-Hydroxybutyrate) Films by Water-Droplet Templating. J. Biomater. Nanobiotechnol. 2012, 3, 431–439. [Google Scholar] [CrossRef] [Green Version]
- Marucci, M.; Andersson, H.; Hjärtstam, J.; Stevenson, G.; Baderstedt, J.; Stading, M.; Larsson, A.; Von Corswant, C. New insights on how to adjust the release profile from coated pellets by varying the molecular weight of ethyl cellulose in the coating film. Int. J. Pharm. 2013, 458, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Rudatin, S.; Sen, Y.L.; Woerner, D.L. Association of Kraft Lignin in Aqueous Solution. ACS Symp. Ser. 1989, 397, 144–154. [Google Scholar] [CrossRef]
- Garver, T.M.; Callaghan, P.T. Hydrodynamics of Kraft Lignins. Macromolecules 1991, 24, 420–430. [Google Scholar] [CrossRef]
- Dutta, S.; Garver, T.M.; Sarkanen, S. Modes of Association between Kraft Lignin Components. Lignin 1989, 397, 155–176. [Google Scholar] [CrossRef]
- Norgren, M.; Edlund, H.; Wågberg, L. Aggregation of lignin derivatives under alkaline conditions. Kinetics and aggregate structure. Langmuir 2002, 18, 2859–2865. [Google Scholar] [CrossRef]
- Norgren, M.; Edlund, H.; Wågberg, L.; Lindström, B.; Annergren, G. Aggregation of kraft lignin derivatives under conditions relevant to the process, part I: Phase behaviour. Colloids Surf. A Physicochem. Eng. Asp. 2001, 194, 85–96. [Google Scholar] [CrossRef]
- Favis, B.D. The Leaching of Lignin Marcomolecules from Pulp Fibers during Washing. 1981. Available online: https://escholarship.mcgill.ca/concern/theses/ww72bc494 (accessed on 19 April 2022).
- Favis, B.D.; Choi, P.M.K.; Adler, P.M.; Goring, D.A.I. Leaching of lignin from unbleached kraft fibers suspended in water.pdf. Pulp Pap. Can. 1981, 82, TR35–TR40. [Google Scholar]
- Magnusdottir, S.; Aakerman, B.; Jonsson, M. DNA Electrophoresis in Agarose Gels: Three Regimes of DNA Migration Identified and Characterized by the Electrophoretic Orientational Behavior of DNA. J. Phys. Chem. 1994, 98, 2624–2633. [Google Scholar] [CrossRef]
- Larsson, A.; Åkerman, B. Period Times and Helix Alignment during the Cyclic Migration of DNA in Electrophoresis Gels Studied with Fluorescence Microscopy. Macromolecules 1995, 28, 4441–4454. [Google Scholar] [CrossRef]
- de Gennes, P.G. Reptation of a polymer chain in the presence of fixed obstacles. J. Chem. Phys. 1971, 55, 572–579. [Google Scholar] [CrossRef]
- Klein, J. Dynamics of Entangled Linear, Branched, and Cyclic Polymers. Macromolecules 1986, 19, 105–118. [Google Scholar] [CrossRef]
- Almqvist, H.; Veras, H.; Li, K.; Hidalgo, J.G.; Hulteberg, C.; Gorwa-Grauslund, M.; Parachin, N.S.; Carlquist, M. Muconic Acid Production Using Engineered Pseudomonas putida KT2440 and a Guaiacol-Rich Fraction Derived from Kraft Lignin. ACS Sustain. Chem. Eng. 2021, 9, 8097–8106. [Google Scholar] [CrossRef]
- Zakis, G.F. Functional Analysis of Lignins and Their Derivatives; Tappi Press: Norcross, GA, USA, 1994. [Google Scholar]
- Gärtner, A.; Gellerstedt, G.; Tamminen, T. Determination of phenolic hydroxyl groups in residual lignin using a modified UV-method. Nord. Pulp Pap. Res. J. 1999, 14, 163–170. [Google Scholar] [CrossRef]
- Garcia, M.T.; Leal, J.S.; Ribosa, I. Effect of various parameters on sodium dodecyl sulfate (SDS) flux through a collagen membrane. J. Am. Oil Chem. Soc. 1992, 69, 20–24. [Google Scholar] [CrossRef]
- Löfroth, J.-E.; Ödman, J. Pharmaceutical Compositions for Sustained Release of the Hmg-Coa Reductase Inhibitor Fluvastatin. Google Patent EP0948320A1, 16 April 1998. [Google Scholar]
- Goldmann, W.M.; Ahola, J.; Mankinen, O.; Kantola, A.M.; Komulainen, S.; Telkki, V.-V.; Tanskanen, J. Determination of phenolic hydroxyl groups in technical lignins by ionization difference ultraviolet spectrophotometry (∆ε-IDUS method). Period. Polytech. Chem. Eng. 2017, 61, 93–101. [Google Scholar] [CrossRef] [Green Version]
- Dence, C.W. Methods in Lignin Chemistry; Lin, S.Y., Ed.; Springer Science & Business Media: Berlin, Germany, 1992; Volume 53. [Google Scholar] [CrossRef]
- Heitner, C.; Dimmel, D.R.; Schmidt, J.A. Lignin and Lignans, Advances in Chemistry; Taylor & Francis Group: London, UK, 2010. [Google Scholar] [CrossRef]
- Bikova, T.; Treimanis, A.; Rossinska, G.; Telysheva, G. On-line study of lignin behaviour in dilute alkaline solution by the SEC-UV method. Holzforschung 2004, 58, 489–494. [Google Scholar] [CrossRef]
- Tiainen, E.; Drakenberg, T.; Tamminen, T.; Kataja, K.; Hase, A. Determination of phenolic hydroxyl groups in lignin by combined use of1H NMR and UV spectroscopy. Holzforschung 1999, 53, 529–533. [Google Scholar] [CrossRef]
- Glasser, G.W.; Sarkanen, S. Lignin Structure and Reactions. Am. Chem. Soc. 1989, 59, 1–21. [Google Scholar] [CrossRef]
- Crestini, C.; Lange, H.; Sette, M.; Argyropoulos, D.S. On the structure of softwood kraft lignin. Green Chem. 2017, 19, 4104–4121. [Google Scholar] [CrossRef]
- Fritz, C.; Salas, C.; Jameel, H.; Rojas, O.J. Self-association and aggregation of kraft lignins via electrolyte and nonionic surfactant regulation: Stabilization of lignin particles and effects on filtration. Nord. Pulp Pap. Res. J. 2017, 32, 572–585. [Google Scholar] [CrossRef]
- van den Mooter, G.; Samyn, C.; Kinget, R. Characterization of colon-specific azo polymers: A study of the swelling propertoes and the permeability of isolated polymer films. Int. J. Pharm. 1994, 111, 127–136. [Google Scholar] [CrossRef]
- Takeshi, N.; Nagata, M.; Yotsuyanagi, T.; Ikeda, K. Micelle-mediated transport of vitamin K1 through porous membranes: Contribution of phosphatidylcholine-bile salt mixed micelles. Chem. Pharm. Bull. 1989, 37, 2496–2499. Available online: http://www.mendeley.com/research/geology-volcanic-history-eruptive-style-yakedake-volcano-group-central-japan/ (accessed on 15 March 2022).
- Kazi, K.M.F.; Gauvin, H.; Jollez, P.; Chornet, E. A diffusion model for the impregnation of lignocellulosic materials. Tappi J. 1997, 80, 209–219. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghaffari, R.; Almqvist, H.; Nilsson, R.; Lidén, G.; Larsson, A. Mass Transport of Lignin in Confined Pores. Polymers 2022, 14, 1993. https://doi.org/10.3390/polym14101993
Ghaffari R, Almqvist H, Nilsson R, Lidén G, Larsson A. Mass Transport of Lignin in Confined Pores. Polymers. 2022; 14(10):1993. https://doi.org/10.3390/polym14101993
Chicago/Turabian StyleGhaffari, Roujin, Henrik Almqvist, Robin Nilsson, Gunnar Lidén, and Anette Larsson. 2022. "Mass Transport of Lignin in Confined Pores" Polymers 14, no. 10: 1993. https://doi.org/10.3390/polym14101993