
Macromolecular Model of the Pectic Polysaccharides Isolated from the Bark of Norway Spruce (Picea abies)



Abstract
:1. Introduction
2. Experimental
2.1. Material
2.2. Isolation of Pectins from the Inner Bark
2.3. Fractionation of the Pectins
2.4. Size Distributions and Absolute Molar Mass Determination with SEC-DRI-MALLS
2.5. Carbohydrate Analyses
2.6. Glycosidic Linkage Analyses
3. Results
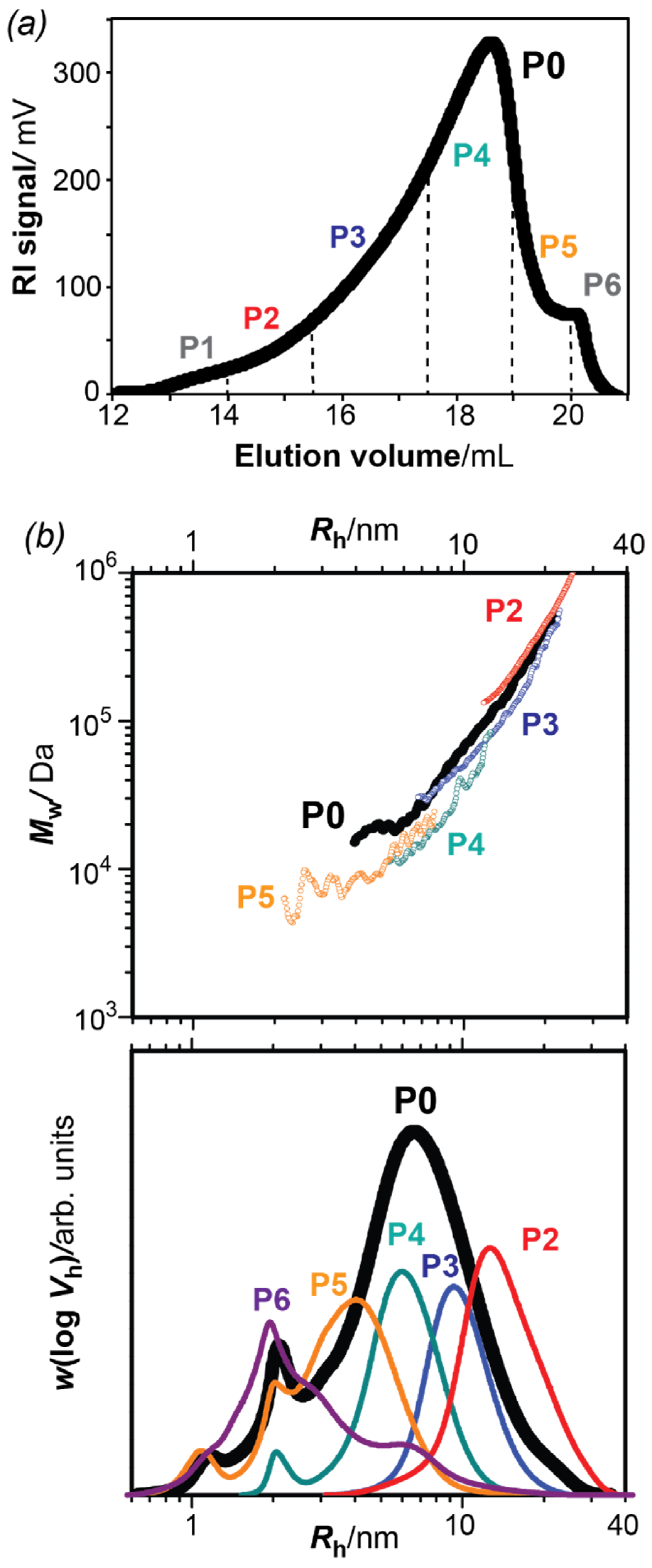
3.1. Fractionation of Bark Pectins and Size Distributions of the Fractions
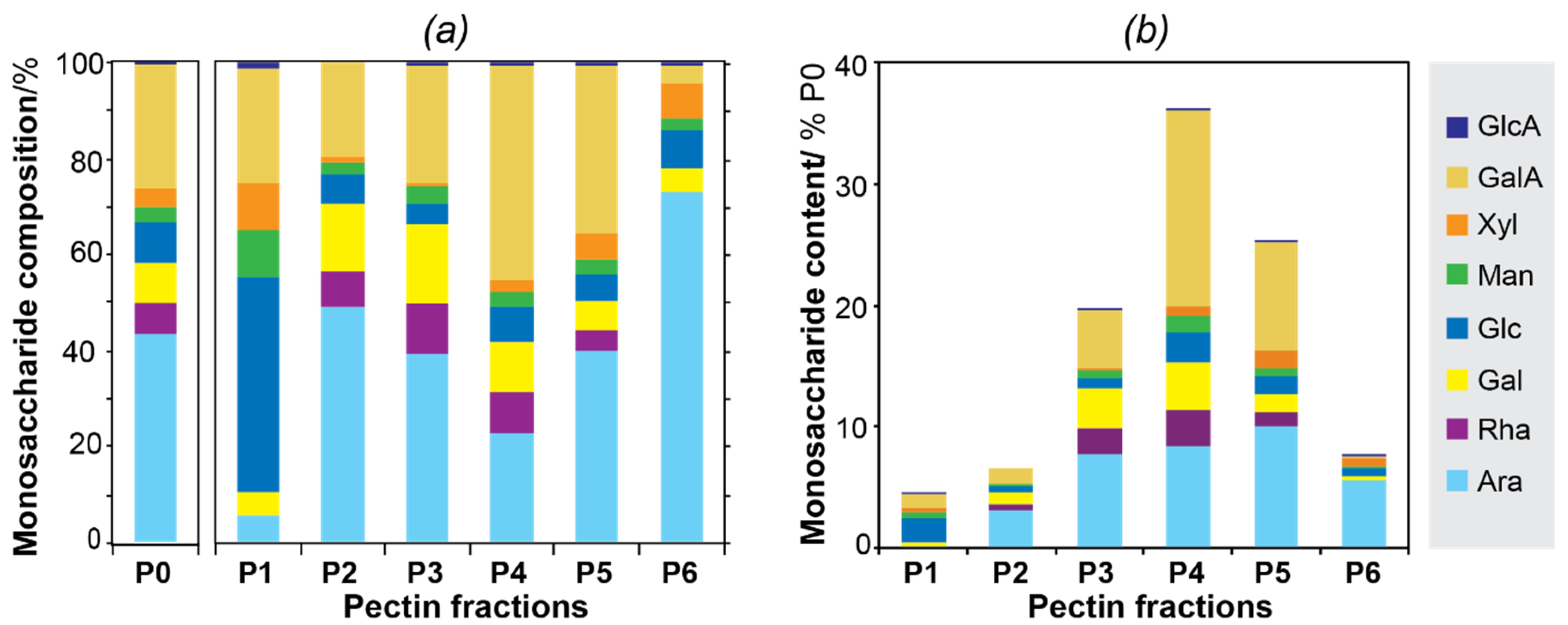
3.2. Carbohydrate Composition
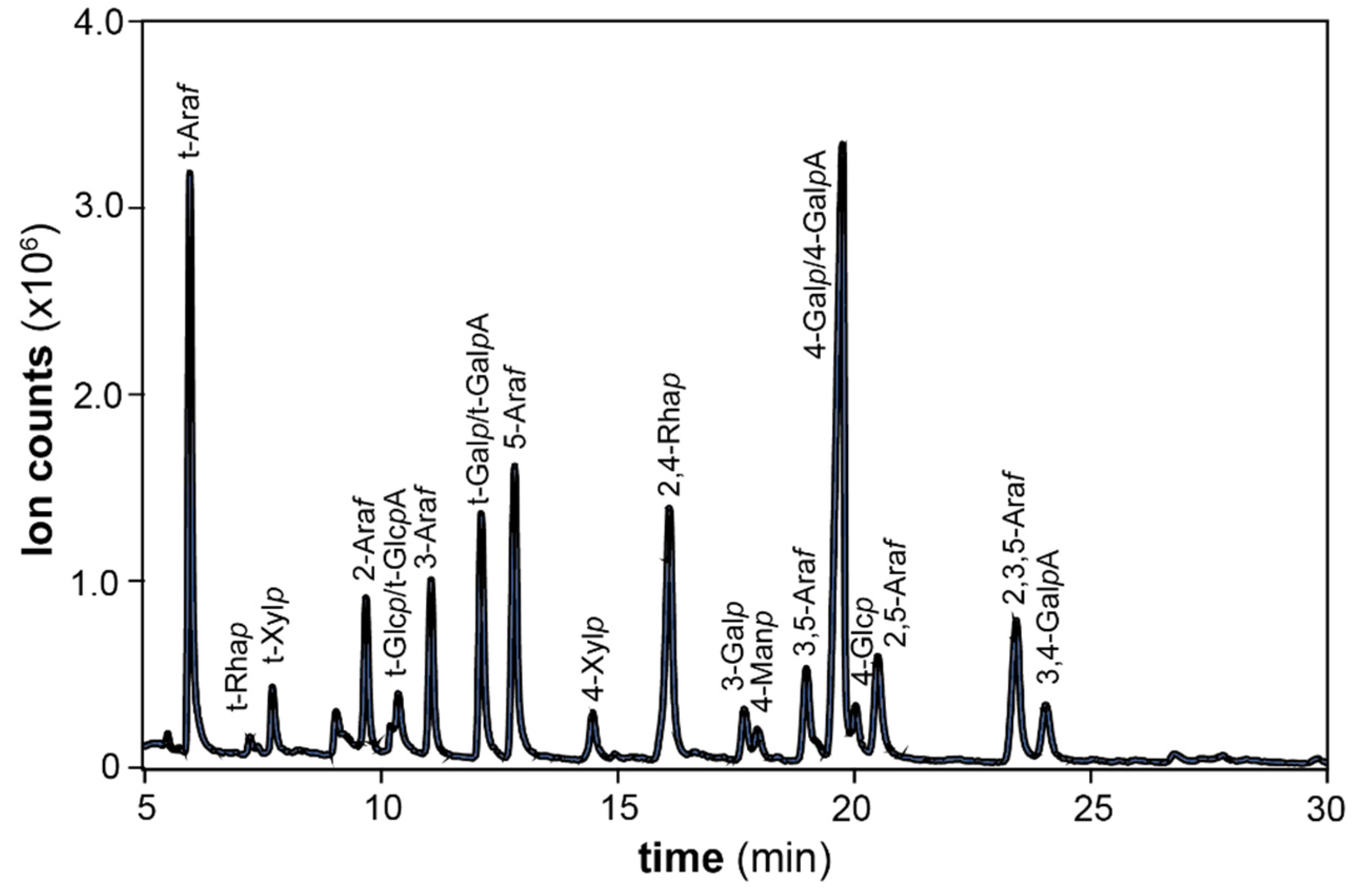
3.3. Glycosidic Linkage Analysis
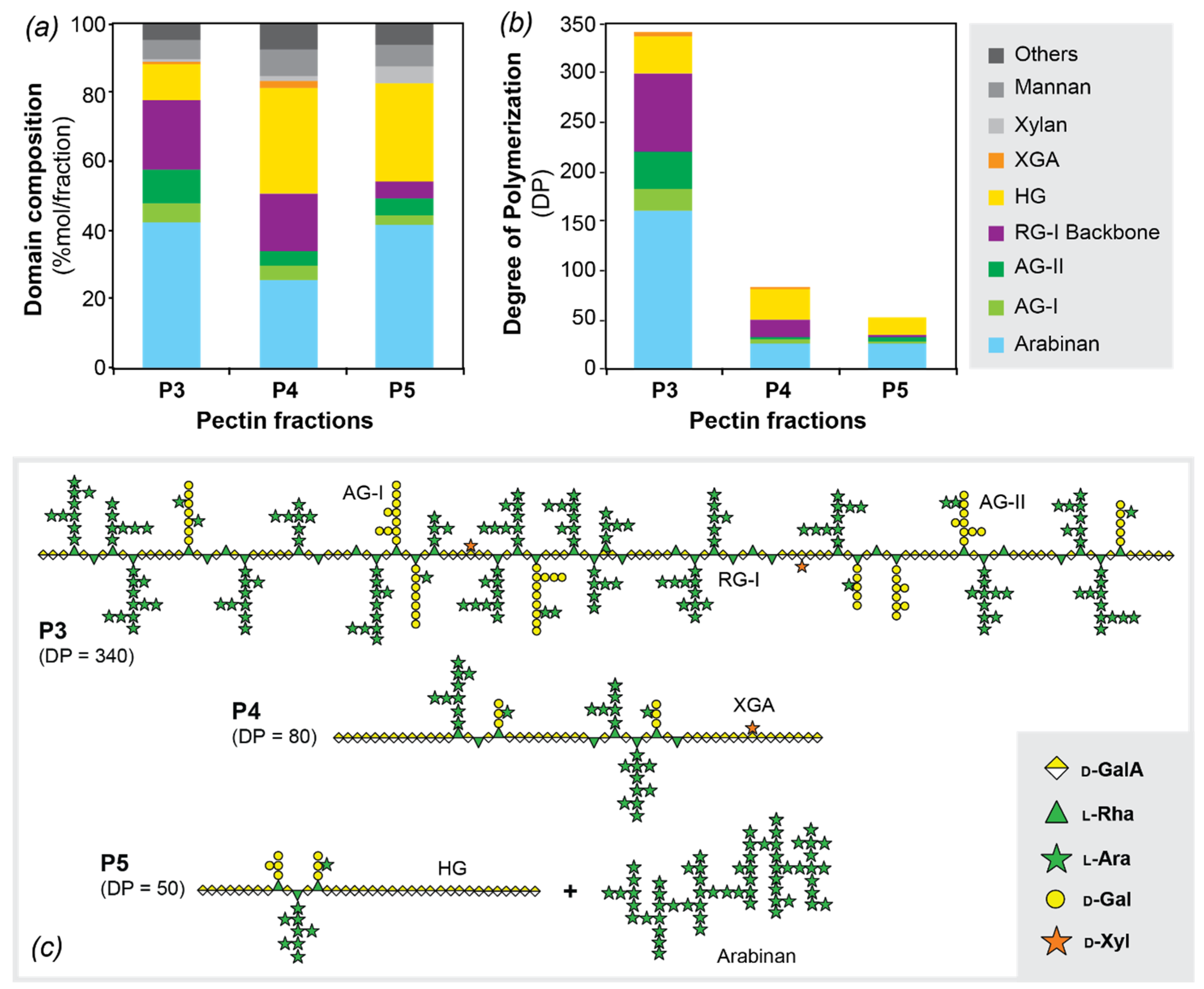
3.4. Pectic Domain Composition. Structural Model for the Pectins in Norway Spruce Bark
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Eriksson, T. Stocks and Consumption of Raw Materials; Swedish Forest Agency: Borås, Sweden, 2013; pp. 189–197. [Google Scholar]
- Ylitalo, E. Wood Consumption. Finnish Statistical Yearbook of Forestry; Finnish Forest Research Institute: Vantaa, Finland, 2013; pp. 247–256. [Google Scholar]
- Le Normand, M.; Edlund, U.; Holmbom, B.; Ek, M. Hot-water extraction and characterization of spruce bark non-cellulosic polysaccharides. Nord. Pulp Pap. Res. J. 2012, 27, 18–23. [Google Scholar] [CrossRef]
- Le Normand, M.; Melida, H.; Holmbom, B.; Michaelsen, T.E.; Inngjerdingen, M.; Bulone, V.; Paulsen, B.S.; Ek, M. Hot-water extracts from the inner bark of Norway spruce with immunomodulating activities. Carbohydr. Polym. 2014, 101, 699–704. [Google Scholar] [CrossRef] [PubMed]
- Krogell, J.; Holmbom, B.; Pranovich, A.; Hemming, J.; Willfor, S. Extraction and chemical characterization of Norway spruce inner and outer bark. Nord. Pulp Pap. Res. J. 2012, 27, 6–17. [Google Scholar] [CrossRef]
- Anderson, E.; Pigman, W.W. A Study of the Inner Bark and Cambial Zone of Black Spruce (Picea mariana B.S.P.). Science 1947, 105, 601–602. [Google Scholar] [CrossRef]
- Painter, T.J.; Purves, C.B. Polysaccharides in the inner bark of white spruce. Tappi J. 1960, 43, 729–736. [Google Scholar]
- Bhattacharjee, S.S.; Timell, T.E. A Study of the Pectin Present in the Bark of Amabilis Fir (Abies Amabilis). Can. J. Chem. 1965, 43, 758–765. [Google Scholar] [CrossRef]
- Valentin, L.; Kluczek-Turpeinen, B.; Willfor, S.; Hemming, J.; Hatakka, A.; Steffen, K.; Tuomela, M. Scots pine (Pinus sylvestris) bark composition and degradation by fungi: Potential substrate for bioremediation. Bioresour. Technol. 2010, 101, 2203–2209. [Google Scholar] [CrossRef]
- Willats, W.G.; McCartney, L.; Mackie, W.; Knox, J.P. Pectin: Cell biology and prospects for functional analysis. Plant Mol. Biol. 2001, 47, 9–27. [Google Scholar] [CrossRef]
- Vincken, J.P.; Schols, H.A.; Oomen, R.J.; McCann, M.C.; Ulvskov, P.; Voragen, A.G.; Visser, R.G. If homogalacturonan were a side chain of rhamnogalacturonan I. Implications for cell wall architecture. Plant Physiol. 2003, 132, 1781–1789. [Google Scholar] [CrossRef] [Green Version]
- Yapo, B.M. Pineapple and banana pectins comprise fewer homogalacturonan building blocks with a smaller degree of polymerization as compared with yellow passion fruit and lemon pectins: Implication for gelling properties. Biomacromolecules 2009, 10, 717–721. [Google Scholar] [CrossRef]
- Albersheim, P.; Darvill, A.G.; O’Neill, M.A.; Schols, H.A.; Voragen, A.G.J. An hypothesis: The same six polysaccharides are components of the primary cell walls of all higher plants. Prog. Biotechnol. 1996, 14, 47–55. [Google Scholar]
- O’Neill, M.A.; Warrenfeltz, D.; Kates, K.; Pellerin, P.; Doco, T.; Darvill, A.G.; Albersheim, P. Rhamnogalacturonan-II, a pectic polysaccharide in the walls of growing plant cell, forms a dimer that is covalently cross-linked by a borate ester. In vitro conditions for the formation and hydrolysis of the dimer. J. Biol. Chem. 1996, 271, 22923–22930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noreen, A.; Nazli, Z.I.; Akram, J.; Rasul, I.; Mansha, A.; Yaqoob, N.; Iqbal, R.; Tabasum, S.; Zuber, M.; Zia, K.M. Pectins functionalized biomaterials; a new viable approach for biomedical applications: A review. Int. J. Biol. Macromol. 2017, 101, 254–272. [Google Scholar] [CrossRef] [PubMed]
- Scheller, H.V.; Jensen, J.K.; Sorensen, S.O.; Harholt, J.; Geshi, N. Biosynthesis of pectin. Physiol. Plant 2007, 129, 283–295. [Google Scholar] [CrossRef]
- Caffall, K.H.; Mohnen, D. The structure, function, and biosynthesis of plant cell wall pectic polysaccharides. Carbohydr. Res. 2009, 344, 1879–1900. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.J.; Chen, W.J.; Zou, M.M.; Lv, R.L.; Wang, D.L.; Hou, F.R.; Feng, H.; Ma, X.B.; Zhong, J.J.; Ding, T.; et al. Applications of power ultrasound in oriented modification and degradation of pectin: A review. J. Food Eng. 2018, 234, 98–107. [Google Scholar] [CrossRef]
- Shafie, M.H.; Yusof, R.; Samsudin, D.; Gan, C.Y. Averrhoa bilimbi pectin-based edible films: Effects of the linearity and branching of the pectin on the physicochemical, mechanical, and barrier properties of the films. Int. J. Biol. Macromol. 2020, 163, 1276–1282. [Google Scholar] [CrossRef]
- Mohamed, S.A.A.; El-Sakhawy, M.; El-Sakhawy, M.A. Polysaccharides, Protein and Lipid -Based Natural Edible Films in Food Packaging: A Review. Carbohydr. Polym. 2020, 238, 116178. [Google Scholar] [CrossRef]
- Sun, Q.L.; Li, Y.X.; Cui, Y.S.; Jiang, S.L.; Dong, C.X.; Du, J. Structural characterization of three polysaccharides from the roots of Codonopsis pilosula and their immunomodulatory effects on RAW264.7 macrophages. Int. J. Biol. Macromol. 2019, 130, 556–563. [Google Scholar] [CrossRef]
- Coimbra, P.; Ferreira, P.; de Sousa, H.C.; Batista, P.; Rodrigues, M.A.; Correia, I.J.; Gil, M.H. Preparation and chemical and biological characterization of a pectin/chitosan polyelectrolyte complex scaffold for possible bone tissue engineering applications. Int. J. Biol. Macromol. 2011, 48, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Smistad, G.; Boyum, S.; Alund, S.J.; Samuelsen, A.B.; Hiorth, M. The potential of pectin as a stabilizer for liposomal drug delivery systems. Carbohydr. Polym. 2012, 90, 1337–1344. [Google Scholar] [CrossRef]
- Kilpeläinen, P.O.; Hautala, S.S.; Byman, O.O.; Tanner, L.J.; Korpinen, R.I.; Lillandt, M.K.J.; Pranovich, A.V.; Kitunen, V.H.; Willför, S.M.; Ilvesniemi, H.S. Pressurized hot water flow-through extraction system scale up from the laboratory to the pilot scale. Green Chem. 2014, 16, 3186–3194. [Google Scholar] [CrossRef]
- Rissanen, J.V.; Grenman, H.; Xu, C.L.; Willfor, S.; Murzin, D.Y.; Salmi, T. Obtaining Spruce Hemicelluloses of Desired Molar Mass by using Pressurized Hot Water Extraction. ChemSusChem 2014, 7, 2947–2953. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Abad, A.; Giummarella, N.; Lawoko, M.; Vilaplana, F. Differences in extractability under subcritical water reveal interconnected hemicellulose and lignin recalcitrance in birch hardwoods. Green Chem. 2018, 20, 2534–2546. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Abad, A.; Jiménez-Quero, A.; Wohlert, J.; Vilaplana, F. Influence of the molecular motifs of mannan and xylan populations on their recalcitrance and organization in spruce softwoods. Green Chem. 2020, 22, 3956–3970. [Google Scholar] [CrossRef]
- Rudjito, R.C.; Ruthes, A.C.; Jimenez-Quero, A.; Vilaplana, F. Feruloylated Arabinoxylans from Wheat Bran: Optimization of Extraction Process and Validation at Pilot Scale. ACS Sustain. Chem. Eng. 2019, 7, 13167–13177. [Google Scholar] [CrossRef]
- Kemppainen, K.; Siika-Aho, M.; Pattathil, S.; Giovando, S.; Kruus, K. Spruce bark as an industrial source of condensed tannins and non-cellulosic sugars. Ind. Crop. Prod. 2014, 52, 158–168. [Google Scholar] [CrossRef]
- Grubisic, Z.; Rempp, P.; Benoit, H. A universal calibration for gel permeation chromatography. J. Polym. Sci. Part B Polym. Lett. 1967, 5, 753–759. [Google Scholar] [CrossRef]
- Hamielec, A.E.; Ouano, A.C. Generalized Universal Molecular-Weight Calibration Parameter in Gpc. J. Liq. Chromatogr. 1978, 1, 111–120. [Google Scholar] [CrossRef]
- Sullivan, M.A.; Powell, P.O.; Witt, T.; Vilaplana, F.; Roura, E.; Gilbert, R.G. Improving size-exclusion chromatography separation for glycogen. J. Chromatogr. A 2014, 1332, 21–29. [Google Scholar] [CrossRef]
- Ciric, J.; Oostland, J.; de Vries, J.W.; Woortman, A.J.; Loos, K. Size exclusion chromatography with multi detection in combination with matrix-assisted laser desorption ionization-time-of-flight mass spectrometry as a tool for unraveling the mechanism of the enzymatic polymerization of polysaccharides. Anal. Chem. 2012, 84, 10463–10470. [Google Scholar] [CrossRef] [PubMed]
- Vilaplana, F.; Gilbert, R.G. Characterization of branched polysaccharides using multiple-detection size separation techniques. J. Sep. Sci. 2010, 33, 3537–3554. [Google Scholar] [CrossRef] [PubMed]
- Zimm, B.H. The Scattering of Light and the Radial Distribution Function of High Polymer Solutions. J. Chem. Phys. 1948, 16, 1093–1099. [Google Scholar] [CrossRef]
- Chapman, H.D.; Morris, V.J.; Selvendran, R.R.; Oneill, M.A. Static and Dynamic Light-Scattering-Studies of Pectic Polysaccharides from the Middle Lamellae and Primary-Cell Walls of Cider Apples. Carbohydr. Res. 1987, 165, 53–68. [Google Scholar] [CrossRef]
- De Ruiter, G.A.; Schols, H.A.; Voragen, A.G.J.; Rombouts, F.M. Carbohydrate analysis of water-soluble uronic acid-containing polysaccharides with high-performance anion-exchange chromatography using methanolysis combined with TFA hydrolysis is superior to four other methods. Anal. Biochem. 1992, 207, 176–185. [Google Scholar] [CrossRef]
- McKee, L.S.; Sunner, H.; Anasontzis, G.E.; Toriz, G.; Gatenholm, P.; Bulone, V.; Vilaplana, F.; Olsson, L. A GH115 alpha-glucuronidase from Schizophyllum commune contributes to the synergistic enzymatic deconstruction of softwood glucuronoarabinoxylan. Biotechnol. Biofuels 2016, 9, 2. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.B.; Carpita, N.C. Changes in Esterification of the Uronic Acid Groups of Cell Wall Polysaccharides during Elongation of Maize Coleoptiles. Plant Physiol. 1992, 98, 646–653. [Google Scholar] [CrossRef] [Green Version]
- Ciucanu, I.; Kerek, F. A Simple and Rapid Method for the Permethylation of Carbohydrates. Carbohydr. Res. 1984, 131, 209–217. [Google Scholar] [CrossRef]
- Albersheim, P.; Nevins, D.J.; English, P.D.; Karr, A. A method for the analysis of sugars in plant cell-wall polysaccharides by gas-liquid chromatography. Carbohydr. Res. 1967, 5, 340–345. [Google Scholar] [CrossRef]
- Carpita, N.C.; Shea, E.M. Linkage structure of carbohydrates by gas chromatography-mass spectrometry (GC-MS) of partially methylated alditol acetates. In Analysis of Carbohydrates by GLC and MS; Biermann, C.J., Ed.; CRC Press, Inc.: Boca Raton, FL, USA, 1989; pp. 157–216. [Google Scholar]
- Fishman, M.L.; Chau, H.K.; Hoagland, P.D.; Hotchkiss, A.T. Microwave-assisted extraction of lime pectin. Food Hydrocoll. 2006, 20, 1170–1177. [Google Scholar] [CrossRef]
- Zykwinska, A.; Boiffard, M.H.; Kontkanen, H.; Buchert, J.; Thibault, J.F.; Bonnin, E. Extraction of green labeled pectins and pectic oligosaccharides from plant byproducts. J. Agric. Food Chem. 2008, 56, 8926–8935. [Google Scholar] [CrossRef]
- Hunter, J.L.; Wicker, L. De-esterification of pectin by alkali, plant and fungal pectinmethylesterases and effect on molecular weight. J. Sci. Food Agric. 2005, 85, 2243–2248. [Google Scholar] [CrossRef]
- Nakauma, M.; Funami, T.; Noda, S.; Ishihara, S.; Al-Assaf, S.; Nishinari, K.; Phillips, G.O. Comparison of sugar beet pectin, soybean soluble polysaccharide, and gum arabic as food emulsifiers. 1. Effect of concentration, pH, and salts on the emulsifying properties. Food Hydrocoll. 2008, 22, 1254–1267. [Google Scholar] [CrossRef]
- Inngjerdingen, K.T.; Coulibaly, A.; Diallo, D.; Michaelsen, T.E.; Paulsen, B.S. A complement fixing polysaccharide from Biophytum petersianum Klotzsch, a medicinal plant from Mali, West Africa. Biomacromolecules 2006, 7, 48–53. [Google Scholar] [CrossRef]
- Hokputsa, S.; Harding, S.E.; Inngjerdingen, K.; Jumel, K.; Michaelsen, T.E.; Heinze, T.; Koschella, A.; Paulsen, B.S. Bioactive polysaccharides from the stems of the Thai medicinal plant Acanthus ebracteatus: Their chemical and physical features. Carbohydr. Res. 2004, 339, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Samuelsen, A.B.; Westereng, B.; Yousif, O.; Holtekjolen, A.K.; Michaelsen, T.E.; Knutsen, S.H. Structural features and complement-fixing activity of pectin from three Brassica oleracea varieties: White cabbage, kale, and red kale. Biomacromolecules 2007, 8, 644–649. [Google Scholar] [CrossRef]
- Schols, H.A.; Bakx, E.J.; Schipper, D.; Voragen, A.G.J. A xylogalacturonan subunit present in the modified hairy regions of apple pectin. Carbohydr. Res. 1995, 279, 265–279. [Google Scholar] [CrossRef]
- Makarova, E.N.; Shakhmatov, E.G.; Belyy, V.A. Structural studies of water-extractable pectic polysaccharides and arabinogalactan proteins from Picea abies greenery. Carbohydr. Polym. 2018, 195, 207–217. [Google Scholar] [CrossRef]
- Shakhmatov, E.G.; Belyy, V.A.; Makarova, E.N. Structure of acid-extractable polysaccharides of tree greenery of Picea abies. Carbohydr. Polym. 2018, 199, 320–330. [Google Scholar] [CrossRef]
- Pettolino, F.A.; Walsh, C.; Fincher, G.B.; Bacic, A. Determining the polysaccharide composition of plant cell walls. Nat. Protoc. 2012, 7, 1590–1607. [Google Scholar] [CrossRef]
- Yapo, B.M.; Lerouge, P.; Thibault, J.F.; Ralet, M.C. Pectins from citrus peel cell walls contain homogalacturonans homogenous with respect to molar mass, rhamnogalacturonan I and rhamnogalacturonan II. Carbohydr. Polym. 2007, 69, 426–435. [Google Scholar] [CrossRef]
- Mohnen, D. Pectin structure and biosynthesis. Curr. Opin. Plant Biol. 2008, 11, 266–277. [Google Scholar] [CrossRef]
- Khodaei, N.; Karboune, S. Extraction and structural characterisation of rhamnogalacturonan I-type pectic polysaccharides from potato cell wall. Food Chem. 2013, 139, 617–623. [Google Scholar] [CrossRef]
- Ele-Ekouna, J.P.; Pau-Roblot, C.; Courtois, B.; Courtois, J. Chemical characterization of pectin from green tea (Camellia sinensis). Carbohydr. Polym. 2011, 83, 1232–1239. [Google Scholar] [CrossRef]
- Yang, J.S.; Mu, T.H.; Ma, M.M. Extraction, structure, and emulsifying properties of pectin from potato pulp. Food Chem. 2018, 244, 197–205. [Google Scholar] [CrossRef]
- Shakhmatov, E.G.; Toukach, P.V.; Michailowa, C.; Makarova, E.N. Structural studies of arabinan-rich pectic polysaccharides from Abies sibirica L. Biological activity of pectins of A. sibirica. Carbohydr. Polym. 2014, 113, 515–524. [Google Scholar] [CrossRef]
- Willför, S.; Holmbom, B. Isolation and characterisation of water soluble polysaccharides from Norway spruce and Scots pine. Wood Sci. Technol. 2004, 38, 173–179. [Google Scholar] [CrossRef]
- Willför, S.; Sjöholm, R.; Laine, C.; Roslund, M.; Hemming, J.; Holmbom, B. Characterisation of water-soluble galactoglucomannans from Norway spruce wood and thermomechanical pulp. Carbohydr. Polym. 2003, 52, 175–187. [Google Scholar] [CrossRef]
- Joseleau, J.-P.; Chambat, G.; Vignon, M.; Barnoud, F. Chemical and 13C N.M.R. studies on two arabinans from the inner bark of young stems of Rosa glauca. Carbohydr. Res. 1977, 58, 165–175. [Google Scholar] [CrossRef]
- Karácsonyi, S.; Toman, R.; Janeček, F.; Kubačková, M. Polysaccharides from the bark of the white willow (Salix alba L.): Structure of an arabian. Carbohydr. Res. 1975, 44, 285–290. [Google Scholar] [CrossRef]
- Jiang, K.S.; Timell, T.E. Polysaccharides in the bark of aspen (Populus tremuloides). II. Isolation and structure of an arabinan. Cellul. Chem. Technol. 1972, 6, 499–502. [Google Scholar]
- Kar, F.; Arslan, N. Effect of temperature and concentration on viscosity of orange peel pectin solutions and intrinsic viscosity-molecular weight relationship. Carbohydr. Polym. 1999, 40, 277–284. [Google Scholar] [CrossRef]
- Mesbahi, G.; Jamalian, J.; Farahnaky, A. A comparative study on functional properties of beet and citrus pectins in food systems. Food Hydrocoll. 2005, 19, 731–738. [Google Scholar] [CrossRef]
- Lofgren, C.; Guillotin, S.; Evenbratt, H.; Schols, H.; Hermansson, A.M. Effects of calcium, pH, and blockiness on kinetic rheological behavior and microstructure of HM pectin gels. Biomacromolecules 2005, 6, 646–652. [Google Scholar] [CrossRef] [PubMed]
- Fraeye, I.; Duvetter, T.; Doungla, E.; Van Loey, A.; Hendrickx, M. Fine-tuning the properties of pectin–calcium gels by control of pectin fine structure, gel composition and environmental conditions. Trends Food Sci. Technol. 2010, 21, 219–228. [Google Scholar] [CrossRef]
- Zheng, J.Q.; Chen, J.L.; Zhang, H.; Wu, D.M.; Ye, X.Q.; Linardt, R.J.; Chen, S.G. Gelling mechanism of RG-I enriched citrus pectin: Role of arabinose side-chains in cation- and acid-induced gelation. Food Hydrocoll. 2020, 101. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fraction | Mn (Da) | Mw (Da) | D | Mws(a) (Da) | DP (b) | Yield (%) |
|---|---|---|---|---|---|---|
| P0 | 17600 | 39300 | 2.2 | 168 | 230 | 100 |
| P1 | n.d | n.d | n.d | n.d | n.d | 4.5 |
| P2 | 156,500 | 240,200 | 1.5 | 165 | 948 | 6.6 |
| P3 | 46,100 | 58,600 | 1.3 | 171 | 340 | 19.8 |
| P4 | 13,200 | 14,500 | 1.1 | 175 | 80 | 36.2 |
| P5 | 7000 | 8700 | 1.2 | 163 | 50 | 25.2 |
| P6 | 5600 | 14,100 | 2.5 | 157 | 36 | 7.7 |
| Partially Methylated Alditol Acetate (PMAA) | Linkage Type | Short Name | P0 | P3 | P4 | P5 |
|---|---|---|---|---|---|---|
| 2,3,5-Me3-Araf | Araf-(1→ | t-Araf | 12.6 | 13.7 | 10.4 | 15.0 |
| 2,3,4-Me3-Arap | Arap-(1→ | t-Arap | 0.3 | 0.2 | 0.3 | 0.9 |
| 3,5-Me2-Araf | → 2)-Araf-(1→ | 2-Araf | 0.5 | 0.8 | 0.4 | 1.8 |
| 2,5-Me2-Araf | → 3)-Araf-(1→ | 3-Araf | 4.5 | 6.8 | 3.1 | 5.2 |
| 2,3-Me2-Araf | → 5)-Araf-(1→ | 5-Araf | 12.4 | 12.6 | 6.5 | 12.5 |
| 2-Me-Araf | → 3,5)-Araf-(1→ | 3,5-Araf | 3.6 | 5.3 | 2.2 | 3.5 |
| 3-Me-Araf | → 2,5)-Araf-(1→ | 2,5-Araf | 1.7 | 2.5 | 1.0 | 1.7 |
| Acetylated Araf | → 2,3,5)-Araf-(1→ | 2,3,5-Araf | 5.4 | 2.3 | 3.2 | 4.5 |
| Total Ara | (41.0) | (44.2) | (27.1) | (45.1) | ||
| 2,3,4,6-Me4-Galp | Galp-(1→ | t-Galp | 3.6 | 2.8 | 5.1 | 1.5 |
| 2,4,6-Me3-Galp | → 3)-Galp-(1→ | 3-Galp | 1.2 | 2.1 | 1.5 | 0.2 |
| 2,3,6- Me3-Galp | → 4)-Galp-(1→ | 4-Galp | 1.6 | 3.5 | 1.5 | 1.2 |
| 2,3,4-Me3-Galp | → 6)-Galp-(1→ | 6-Galp | 2.9 | 4.9 | 0.6 | 1.0 |
| 2,3-Me2-Galp | → 4,6)-Galp-(1→ | 4,6-Galp | 0.7 | 0.9 | 0.7 | 0.6 |
| 2,6-Me2-Galp | → 3,4)-Galp-(1→ | 3,4-Galp | 0.2 | 0.3 | 0.5 | 0.1 |
| 2,4-Me2-Galp | → 3,6)-Galp-(1→ | 3,6-Galp | 1.2 | 0.9 | 0.7 | 1.2 |
| Total Gal | (11.4) | (15.4) | (10.6) | (5.8) | ||
| 2,3,4,6-Me4-GalpA | GalpA-(1→ | t-GalpA | 2.5 | 0.4 | 1.5 | 2.1 |
| 2,3,6- Me3-GalpA | → 4)-GalpA-(1→ | 4-GalpA | 21.4 | 19.1 | 36.4 | 28.4 |
| 2,6-Me2-GalpA | → 3,4)-GalpA-(1→ | 3,4-GalpA | 1.7 | 1.4 | 2.5 | 0.0 |
| Total GalA | (25.6) | (20.9) | (40.4) | (30.5) | ||
| 2,3,4,6-Me4-Rhap | Rhap-(1→ | t-Rhap | 0.6 | 0.2 | 0.4 | 1.8 |
| 3,4,6-Me3-Rhap | → 2)-Rhap-(1→ | 2-Rhap | 2.2 | 3.4 | 3.5 | 0.0 |
| 3,6-Me2-Rhap | → 2,4)-Rhap-(1→ | 2,4-Rhap | 4.5 | 6.9 | 4.7 | 2.4 |
| Total Rha | (6.3) | (10.5) | (8.6) | (4.2) | ||
| 2,3,4,6-Me4-Glcp | Glcp-(1→ | t-Glcp | 2.0 | 0.2 | 0.9 | 2.6 |
| 2,3,6-Me3-Glcp | → 4)-Glcp-(1→ | 4-Glcp | 2.9 | 2.4 | 3.8 | 2.6 |
| 2,3-Me2-Glcp | → 4,6)-Glcp-(1→ | 4,6-Glcp | 2.7 | 1.4 | 2.3 | 0.2 |
| Total Glc | (7.6) | (4.0) | (7.0) | (5.4) | ||
| 2,3,4-Me3-Xylp | Xylp-(1→ | t-Xylp | 1.4 | 0.6 | 1.2 | 0.8 |
| 2,3-Me2-Xylp | → 4)-Xylp-(1→ | 4-Xylp | 2.0 | 0.6 | 1.3 | 5.4 |
| Total Xyl | (3.4) | (1.2) | (2.5) | (6.2) | ||
| 2,3,6-Me3-Manp | → 4)-Manp-(1→ | 4-Manp | 4.0 | 3.2 | 3.3 | 2.3 |
| 2,3-Me2-Manp | → 4,6)-Manp-(1→ | 4,6-Manp | 0.7 | 0.0 | 0.0 | 0.3 |
| Total Man | (4.7) | (3.2) | (3.3) | (2.6) | ||
| 2,3,4,6-Me4-GlcpA | GlcAp-(1→ | t-GlcpA | 0.3 | 0.4 | 0.4 | n.d. |
| 2,3,6-Me3-GlcpA | → 4)-GlcpA-(1→ | 4-GlcpA | 0.1 | 0.2 | 0.1 | n.d. |
| Total GlcA | (0.4) | (0.5) | (0.5) | (0.2) |
| Polysaccharides | Linkage | P3 | P4 | P5 |
|---|---|---|---|---|
| Arabinan | t-Araf | 12.4 | 9.6 | 14.2 |
| 3-Araf | 6.8 | 3.1 | 5.2 | |
| 5-Araf | 12.6 | 6.5 | 12.5 | |
| 3,5-Araf | 5.3 | 2.2 | 3.5 | |
| 2,5-Araf | 2.5 | 1.0 | 1.7 | |
| 2,3,5-Araf | 2.3 | 3.2 | 4.5 | |
| Total | 41.9 | 25.6 | 41.6 | |
| Type I AG | 4-Galp | 3.5 | 1.5 | 1.2 |
| 4,6-Galp | 0.9 | 0.7 | 0.6 | |
| 3,4-Galp | 0.3 | 0.5 | 0.1 | |
| t-Araf | 1.2 | 0.8 | 0.5 | |
| t-Galp | 0 | 0.8 | 0 | |
| Total | 5.9 | 4.3 | 2.4 | |
| Type II AG | 3-Galp | 2.1 | 1.5 | 0.2 |
| 6-Galp | 4.9 | 0.6 | 1.0 | |
| 2-Araf | 0.8 | 0.4 | 1.8 | |
| 3,6-Galp | 0.9 | 0.7 | 1.2 | |
| t-Rhap | 0.2 | 0.4 | 1.2 | |
| t-Araf/Arap | 0.1 | 0 | 0 | |
| t-Galp | 0.6 | 0.3 | 0 | |
| Total | 9.6 | 3.9 | 5.4 | |
| RG I | 2-Rhap | 3.4 | 3.5 | 0 |
| 2,4-Rhap | 6.9 | 4.7 | 2.4 | |
| 3,4-GalpA | 0.8 | 1.3 | 0 | |
| 4-GalpA | 9.5 | 6.9 | 2.4 | |
| Total | 20.6 | 16.4 | 4.8 | |
| HG | 4-GalpA | 9.6 | 29.5 | 26 |
| t-GalpA | 0.4 | 1.5 | 2.1 | |
| Total | 10.0 | 31.0 | 28.1 | |
| XGA | 3,4-GalpA | 0.6 | 1.2 | 0 |
| t-Xylp | 0.6 | 1.2 | 0 | |
| Total | 1.2 | 2.4 | 0 | |
| HX | 4-Xylp | 0.6 | 1.3 | 5.4 |
| Total | 0.6 | 1.3 | 5.4 | |
| HM | 4-Manp | 3.2 | 3.3 | 2.3 |
| 4,6-Manp | 0 | 0 | 0.3 | |
| 4-Glcp | 2.4 | 3.8 | 2.6 | |
| 4,6-Glcp | 0 | 0 | 0.3 | |
| t-Galp | 0 | 0 | 0.6 | |
| Total | 5.6 | 7.1 | 5.8 | |
| Others | 4.6 | 8.0 | 6.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Le Normand, M.; Rietzler, B.; Vilaplana, F.; Ek, M. Macromolecular Model of the Pectic Polysaccharides Isolated from the Bark of Norway Spruce (Picea abies). Polymers 2021, 13, 1106. https://doi.org/10.3390/polym13071106
Le Normand M, Rietzler B, Vilaplana F, Ek M. Macromolecular Model of the Pectic Polysaccharides Isolated from the Bark of Norway Spruce (Picea abies). Polymers. 2021; 13(7):1106. https://doi.org/10.3390/polym13071106
Chicago/Turabian StyleLe Normand, Myriam, Barbara Rietzler, Francisco Vilaplana, and Monica Ek. 2021. "Macromolecular Model of the Pectic Polysaccharides Isolated from the Bark of Norway Spruce (Picea abies)" Polymers 13, no. 7: 1106. https://doi.org/10.3390/polym13071106