
Microencapsulation of a Pseudomonas Strain (VUPF506) in Alginate–Whey Protein–Carbon Nanotubes and Next-Generation Sequencing Identification of This Strain



Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms
2.2. Materials for Bacterial Microencapsulation
2.3. Encapsulation Process
2.3.1. Extrusion Technique
2.3.2. Spray-Drying Technique
2.3.3. Emulsification Technique
2.4. Scanning Electron Microscopy of Microcapsules (SEM)
2.5. X-ray Diffraction (XRD)
2.6. Determination of Encapsulation Efficiency
2.7. Release Behavior
2.8. Greenhouse Experiments
2.8.1. Preparation of R. solani Inoculum
2.8.2. Preparation of Antagonistic Suspensions
2.8.3. Planting
2.8.4. Disease Assessment
- 0 = no disease symptoms;
- 1 = small lesions (15 mm);
- 2 = large lesions (>15 mm);
- 3 = lesions girdling the stem.
2.9. Genomic DNA Isolation
2.10. Illumina Whole Genome Sequencing
2.11. Phylogenetic Analysis
2.12. Statistical Analysis
3. Results and Discussion
3.1. SEM Morphology of the ALG–WP–CNT Microcapsules
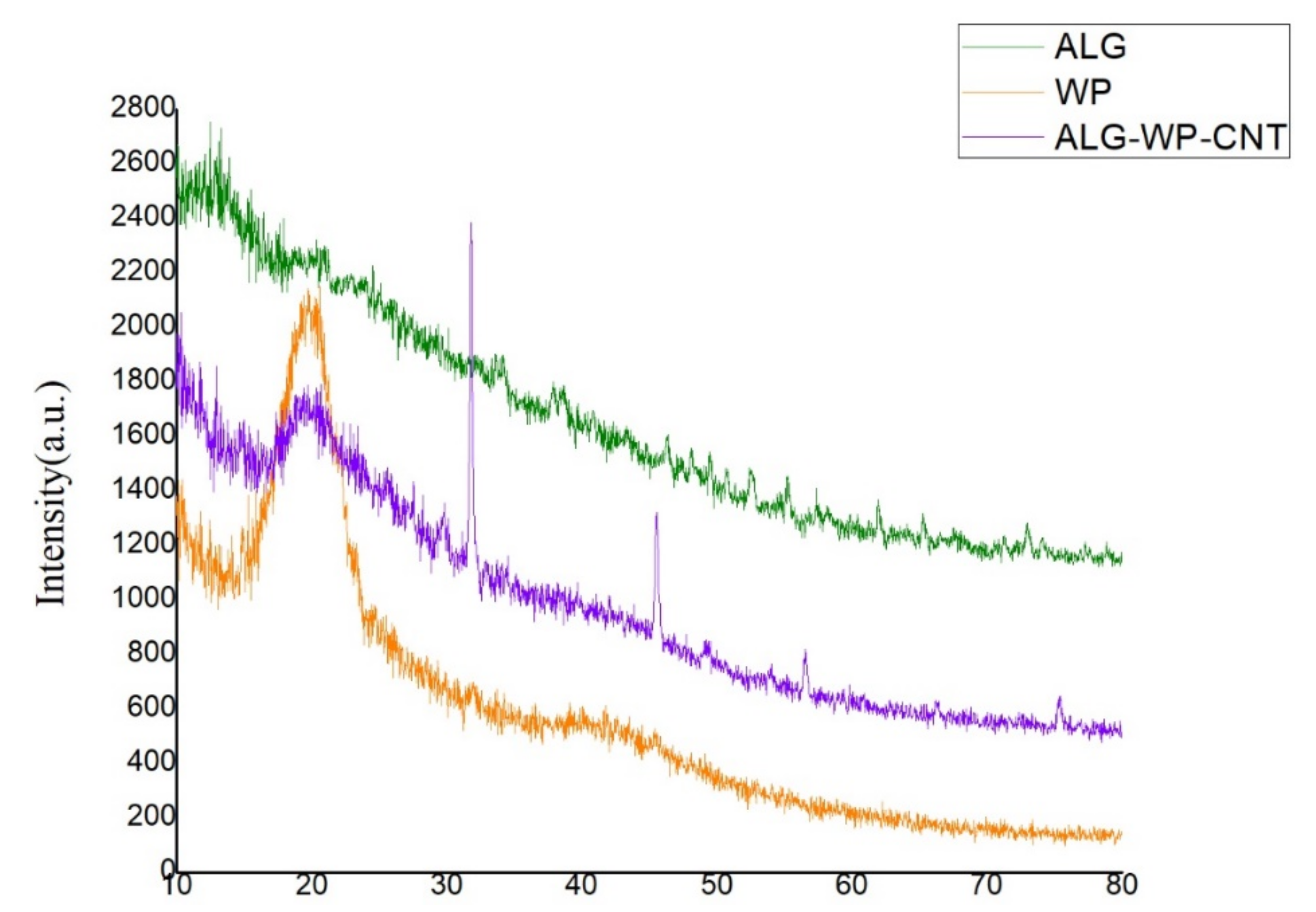
3.2. XRD Structure of the ALG–WP–CNT Microcapsules
3.3. Encapsulation Efficiency
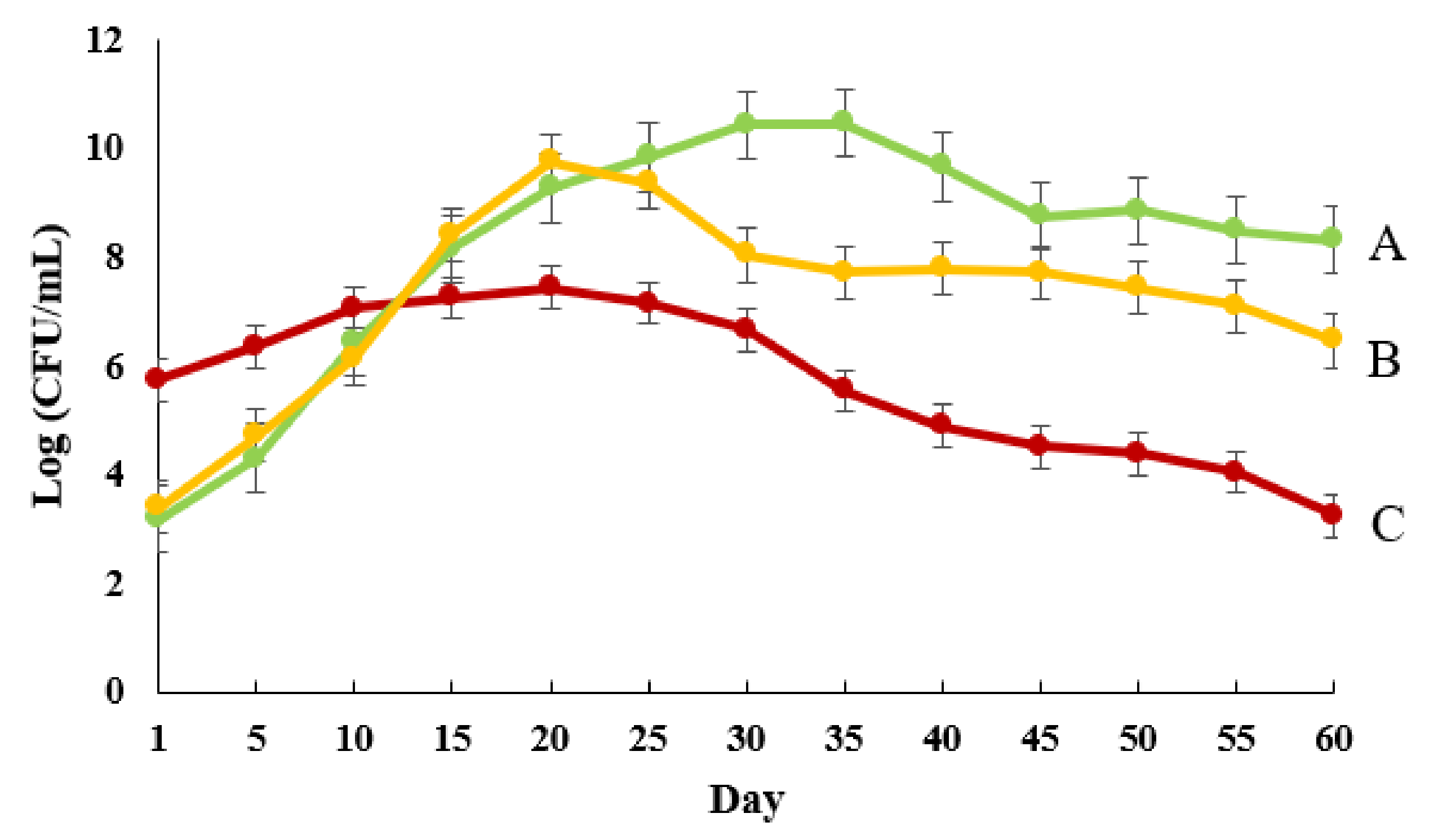
3.4. Release Patterns
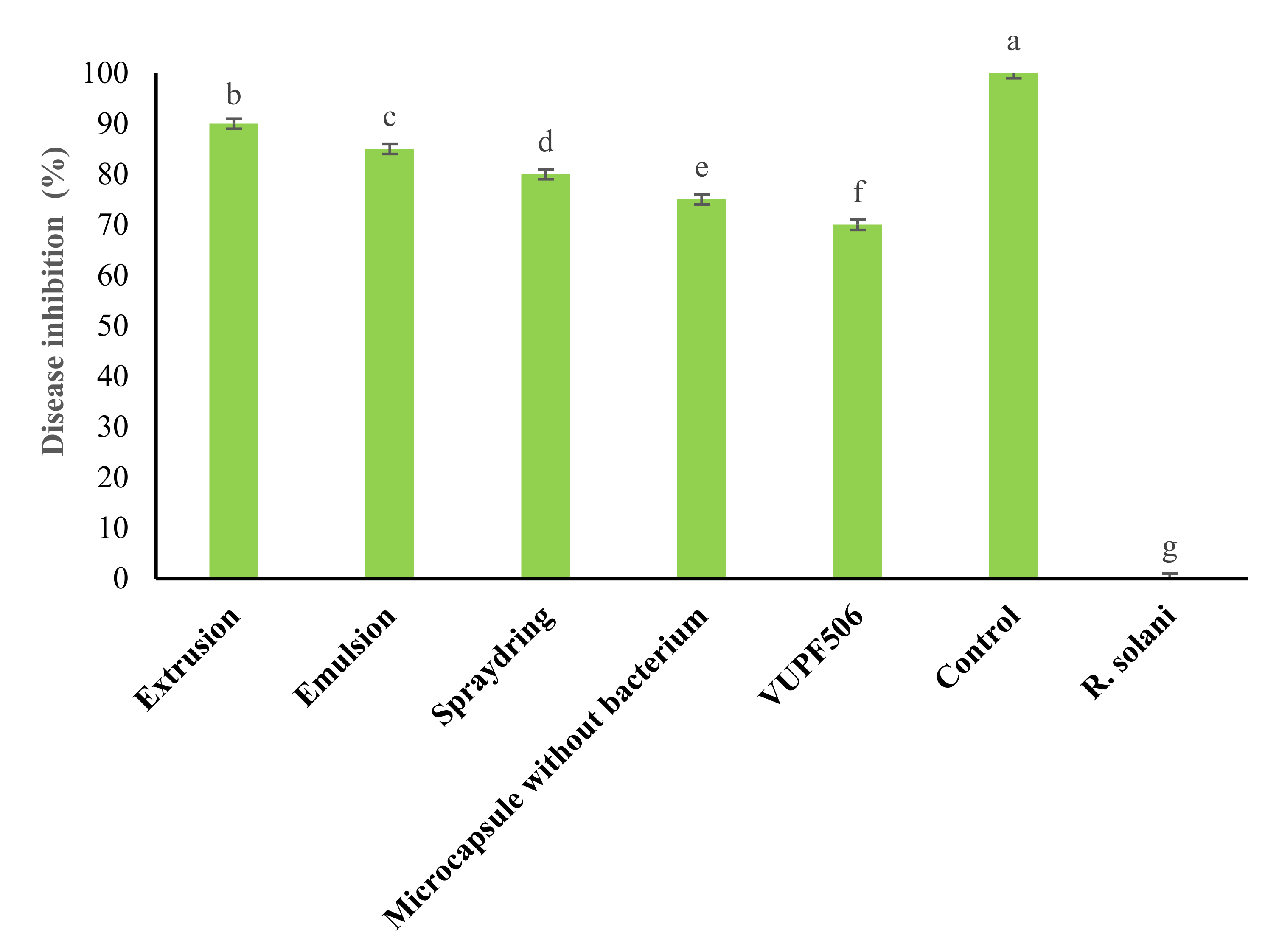
3.5. Greenhouse Results
3.6. Illumina Genome Sequence
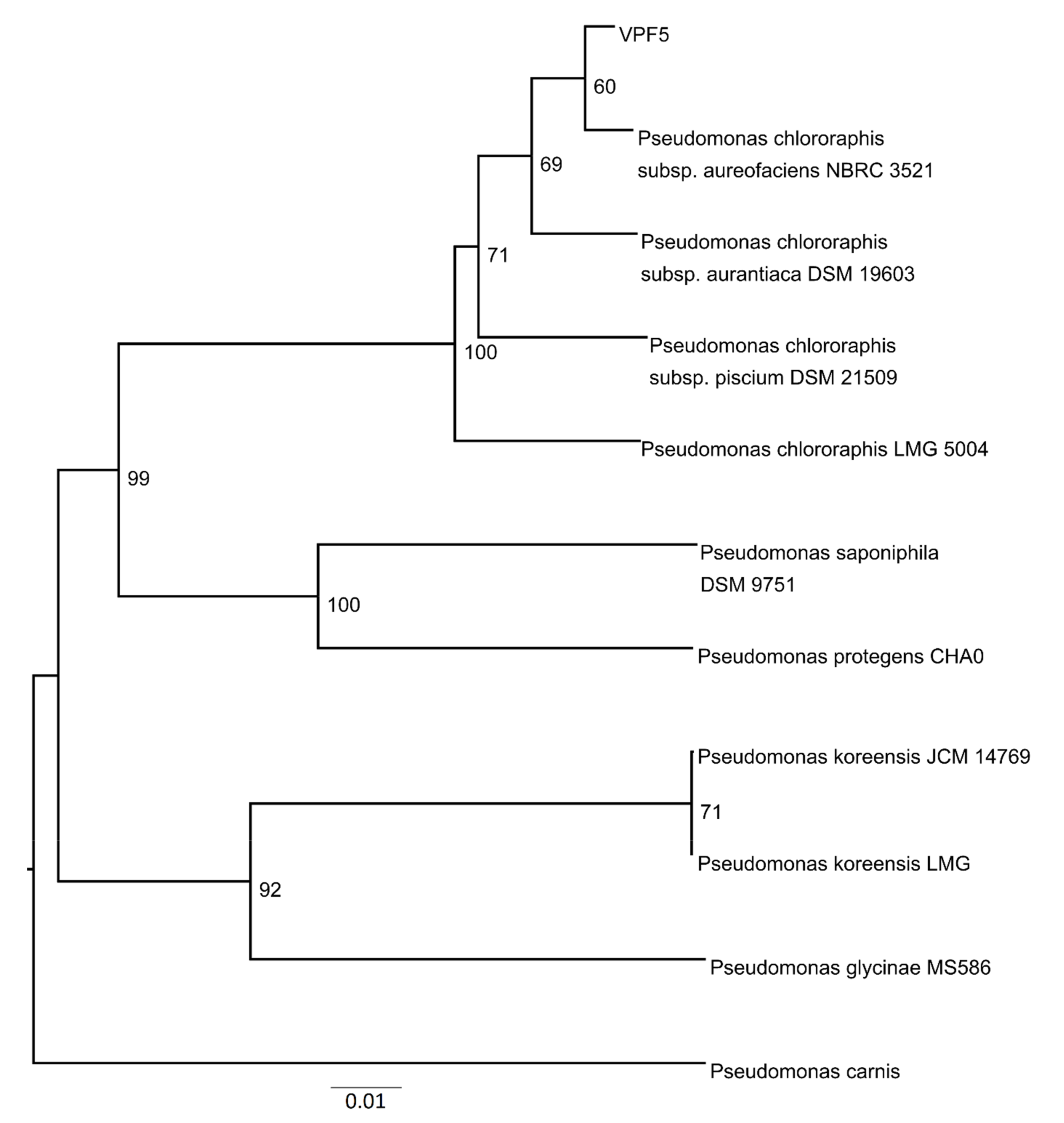
3.7. Phylogenetic Analyses
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kim, I.Y.; Pusey, P.L.; Zhao, Y.; Korban, S.S.; Choi, H.; Kim, K.K. Controlled release of Pantoea agglomerans E325 for biocon-trol of fire blight disease of apple. J. Control. Release 2012, 161, 9–15. [Google Scholar] [CrossRef] [Green Version]
- Guetsky, R.; Shtienberg, D.; Elad, Y.; Dinoor, A. Combining Biocontrol Agents to Reduce the Variability of Biological Control. Phytopathology 2001, 91, 261–267. [Google Scholar] [CrossRef] [Green Version]
- Etchepare, M.D.A.; Barin, J.S.; Cichoski, A.J.; Jacob-Lopes, E.; Wagner, R.; Fries, L.L.M.; De Menezes, C.R. Microencapsulation of probiotics using sodium alginate. Cienc. Rural 2015, 45, 1319–1326. [Google Scholar] [CrossRef] [Green Version]
- Saberi Riseh, S.; Skorik, Y.A.; Thakur, V.K.; Moradi Pour, M.; Tamanadar, E.; Noghabi, S.S. Encapsulation of Plant Biocontrol Bacteria with Alginate as a Main Polymer Material. Int. J. Mol. Sci. 2021, 22, 11165. [Google Scholar] [CrossRef]
- Fathi, F.; Saberi-Riseh, R.; Khodaygan, P. Survivability and controlled release of alginate-microencapsulated Pseudomonas fluorescens VUPF506 and their effects on biocontrol of Rhizoctonia solani on potato. Int. J. Biol. Macromol. 2021, 183, 627–634. [Google Scholar] [CrossRef]
- Rathore, S.; Desai, P.M.; Liew, C.V.; Chan, L.W.; Heng, P.W.S. Microencapsulation of microbial cells. J. Food Eng. 2013, 116, 369–381. [Google Scholar] [CrossRef]
- Lee, Y.; Ji, Y.R.; Lee, S.; Choi, M.-J.; Cho, Y. Microencapsulation of Probiotic Lactobacillus acidophilus KBL409 by Extrusion Technology to Enhance Survival under Simulated Intestinal and Freeze-Drying Conditions. J. Microbiol. Biotechnol. 2019, 29, 721–730. [Google Scholar] [CrossRef] [Green Version]
- Lengyel, M.; Kállai-Szabó, N.; Antal, V.; Laki, A.J.; Antal, I. Microparticles, Microspheres, and Microcapsules for Advanced Drug Delivery. Sci. Pharm. 2019, 87, 20. [Google Scholar] [CrossRef] [Green Version]
- Pereira, K.C.; Ferreira, D.C.M.; Alvarenga, G.F.; Pereira, M.S.S.; Barcelos, M.C.S.; Costa, J.M.G. Microencapsulation and release controlled by the diffusion of food ingredients produced by spray drying: A review, Braz. J. Food Technol. 2018, 211, e2017083. [Google Scholar]
- Fu, W.-Y.; Etzel, M.R. Spray Drying of Lactococcus lactis ssp. lactis C2 and Cellular Injury. J. Food Sci. 1995, 60, 195–200. [Google Scholar] [CrossRef]
- Piñón-Balderrama, C.I.; Leyva-Porras, C.; Terán-Figueroa, Y.; Espinosa-Solís, V.; Álvarez-Salas, C.; Saavedra-Leos, M.Z. Encapsulation of Active Ingredients in Food Industry by Spray-Drying and Nano Spray-Drying Technologies. Processes 2020, 8, 889. [Google Scholar] [CrossRef]
- Groboillot, A.F.; Champagne, C.P.; Darling, G.D.; Poncelet, D. Membrane formation by interfacial cross-linking of chitosan for microencapsulation of Lactococcus lactis. Biotechnol. Bioeng. 1993, 42, 1157–1163. [Google Scholar] [CrossRef]
- Khot, L.R.; Sankaran, S.; Maja, J.M.; Ehsani, R.; Schuster, E.W. Applications of nanomaterials in agricultural production and crop protection: A review. Crop. Prot. 2012, 35, 64–70. [Google Scholar] [CrossRef]
- Mondal, A.; Basu, R.; Das, S.; Nandy, P. Beneficial role of carbon nanotubes on mustard plant growth: An agricultural prospect. J. Nanopart. Res. 2011, 13, 4519–4528. [Google Scholar] [CrossRef]
- Wang, X.; Han, H.; Liu, X.; Gu, X.; Chen, K.; Lu, D. Multi-walled carbon nanotubes can enhance root elongation of wheat (Trit-icum aestivum) plants. J. Nanopart. Res. 2012, 14, 841. [Google Scholar] [CrossRef]
- Qin, D. Next-generation sequencing and its clinical application. Cancer Biol. Med. 2019, 16, 4–10. [Google Scholar] [CrossRef]
- Mathimaran, N.; Srivastava, R.; Wiemken, A.; Sharma, A.K.; Boller, T. Genome sequences of two plant growth-promoting flu-orescent Pseudomonas strains, R62 and R81. J. Bacteriol. 2012, 194, 3272–3273. [Google Scholar] [CrossRef] [Green Version]
- Song, J.Y.; Kim, H.A.; Kim, J.-S.; Kim, S.-Y.; Jeong, H.; Kang, S.G.; Kim, B.K.; Kwon, S.-K.; Lee, C.H.; Yu, D.S.; et al. Genome Sequence of the Plant Growth-Promoting Rhizobacterium Bacillus sp. Strain JS. J. Bacteriol. 2012, 194, 3760–3761. [Google Scholar] [CrossRef] [Green Version]
- Matilla, M.A.; Pizarro-Tobias, P.; Roca, A.; Fernaández, M.; Duque, E.; Molina, L.; Wu, X.; van der Lelie, D.; Goómez, M.J.; Segura, A.; et al. Complete Genome of the Plant Growth-Promoting Rhizobacterium Pseudomonas putida BIRD-1. J. Bacteriol. 2011, 193, 1290. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Wang, Q.; Hou, J.; Tu, C.; Luo, Y.; Christie, P. Whole genome analysis of halotolerant and alkalotolerant plant growth-promoting rhizobacterium Klebsiella sp. D5A. Sci. Rep. 2016, 6, 26710. [Google Scholar] [CrossRef]
- Rajam, R.; Karthik, P.; Parthasarathi, S.; Joseph, G.; Anandharamakrishnan, C. Effect of whey protein–alginate wall systems on survival of microencapsulated Lactobacillus plantarum in simulated gastrointestinal conditions. J. Funct. Foods 2012, 4, 891–898. [Google Scholar] [CrossRef]
- Guan, H.; Chi, D.; Yu, J.; Li, H. Encapsulated ecdysone by internal gelation of alginate microspheres for controlling its release and photostability. Chem. Eng. J. 2011, 168, 94–101. [Google Scholar] [CrossRef]
- Chavarri, M.; Marañón, I.; Ares, R.; Carmen, M. Microencapsulation of a probiotic and prebiotic in alginate-chitosan capsules improves survival in simulated gastrointestinal conditions. Int. J. Food Microbiol. 2010, 142, 185–189. [Google Scholar] [CrossRef]
- Alvarez-Roman, R.; Barre, G.; Guy, R.H.; Fessi, H. Biodegradable polymer nanocapsules containing a sunscreen agent: Prepa-ration and photoprotection. Eur. J. Pharm. Biopharm. 2001, 52, 191–195. [Google Scholar] [CrossRef]
- Abbott, W.S. A Method of Computing the Effectiveness of an Insecticide. J. Econ. Éntomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Microb. Bioinf. 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Banjare, I.S.; Gandhi, K.; Sao, K.; Sharma, R. Spray-Dried Whey Protein Concentrate-Iron Complex: Preparation and Physico-chemical Characterization. Food Technol. Biotechnol. 2019, 57, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Paulo, B.B.; Ramos, F.D.M.; Prata, A.S. An investigation of operational parameters of jet cutting method on the size of Ca-alginate beads. J. Food Process. Eng. 2017, 40, e12591. [Google Scholar] [CrossRef]
- Dehkordi, S.S.; Alemzadeh, I.; Vaziri, A.S.; Vossoughi, A. Optimization of Alginate-Whey Protein Isolate Microcapsules for Survivability and Release Behavior of Probiotic Bacteria. Appl. Biochem. Biotechnol. 2019, 190, 182–196. [Google Scholar] [CrossRef] [PubMed]
- Sasrimuang, S.; Chuchuen, O.; Artnaseaw, A. Synthesis, characterization, and electrochemical properties of carbon nanotubes used as cathode materials for Al–air batteries from a renewable source of water hyacinth. Green Process. Synth. 2020, 9, 340–348. [Google Scholar] [CrossRef]
- Maury, M.; Murphy, K.; Kumar, S.; Shi, L.; Lee, G. Effects of process variables on the powder yield of spray-dried trehalose on a laboratory spray-dryer. Eur. J. Pharm. Biopharm. 2005, 59, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Van Elsas, J.; Trevors, J.; Jain, D.; Wolters, A.; Heijnen, C.; Van Overbeek, L. Survival of, and root colonization by, algi-nate-encapsulated Pseudomonas fluorescens cells following introduction into soil. Biol. Fertil. Soils 1992, 14, 14–22. [Google Scholar] [CrossRef]
- Cañas, J.E.; Long, M.; Nations, S.; Vadan, R.; Dai, L.; Luo, M.; Olszyk, D. Effects of functionalized and non-functionalized sin-gle-walled carbon nanotubes on root elongation of select crop species. Environ. Toxicol. Chem. 2008, 27, 1922–1931. [Google Scholar] [CrossRef] [PubMed]
- Moradi Pour, M.; Saberi-Riseh, R.; Mohammadinejad, R.; Hosseini, A. Investigating the formulation of alginate- gelatin en-capsulated Pseudomonas fluorescens (VUPF5 and T17-4 strains) for controlling Fusarium solani on potato. Int. J. Biol. Macromol. 2019, 133, 603–613. [Google Scholar] [CrossRef] [PubMed]
- Lefort, V.; Desper, R.; Gascuel, O. FastME 2.0: A comprehensive, accurate, and fast distance-based phylogeny inference program. Mol. Biol. Evol. 2015, 32, 2798–2800. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Encapsulation Technique | Encapsulation Efficiency (%) * |
|---|---|
| Extrusion | 86.1 ± 1.2 a |
| Emulsification | 81.7 ± 0.5 b |
| Spray drying | 77.5 ± 1.0 c |
| P. fluorescens | P. putida | P. chlororaphis | |
|---|---|---|---|
| Number of mapped sequences | 21,062,282 | 7,294,692 | 10,071,541 |
| Mapped sequences (%) | 96.5 | 33.59 | 46.3 |
| Create Assemblies with Unicycler | |
|---|---|
| # contigs (≥0 bp) | 78 |
| # contigs (≥1000 bp) | 63 |
| Total length (≥0 bp) | 6,483,708 |
| Total length (≥100 bp) | 6,480,725 |
| # contigs | 63 |
| Largest contig | 486,020 |
| Total length | 6,480,725 |
| GC% | 6332 |
| N50 | 153,751 |
| N75 | 106,268 |
| L50 | 12 |
| L75 | 24 |
| #N’s per100 kbp | 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fathi, F.; Saberi Riseh, R.; Khodaygan, P.; Hosseini, S.; Skorik, Y.A. Microencapsulation of a Pseudomonas Strain (VUPF506) in Alginate–Whey Protein–Carbon Nanotubes and Next-Generation Sequencing Identification of This Strain. Polymers 2021, 13, 4269. https://doi.org/10.3390/polym13234269
Fathi F, Saberi Riseh R, Khodaygan P, Hosseini S, Skorik YA. Microencapsulation of a Pseudomonas Strain (VUPF506) in Alginate–Whey Protein–Carbon Nanotubes and Next-Generation Sequencing Identification of This Strain. Polymers. 2021; 13(23):4269. https://doi.org/10.3390/polym13234269
Chicago/Turabian StyleFathi, Fariba, Roohallah Saberi Riseh, Pejman Khodaygan, Samin Hosseini, and Yury A. Skorik. 2021. "Microencapsulation of a Pseudomonas Strain (VUPF506) in Alginate–Whey Protein–Carbon Nanotubes and Next-Generation Sequencing Identification of This Strain" Polymers 13, no. 23: 4269. https://doi.org/10.3390/polym13234269