
Lasso Proteins—Unifying Cysteine Knots and Miniproteins

 , , and
, , and
Abstract
:
1. Introduction
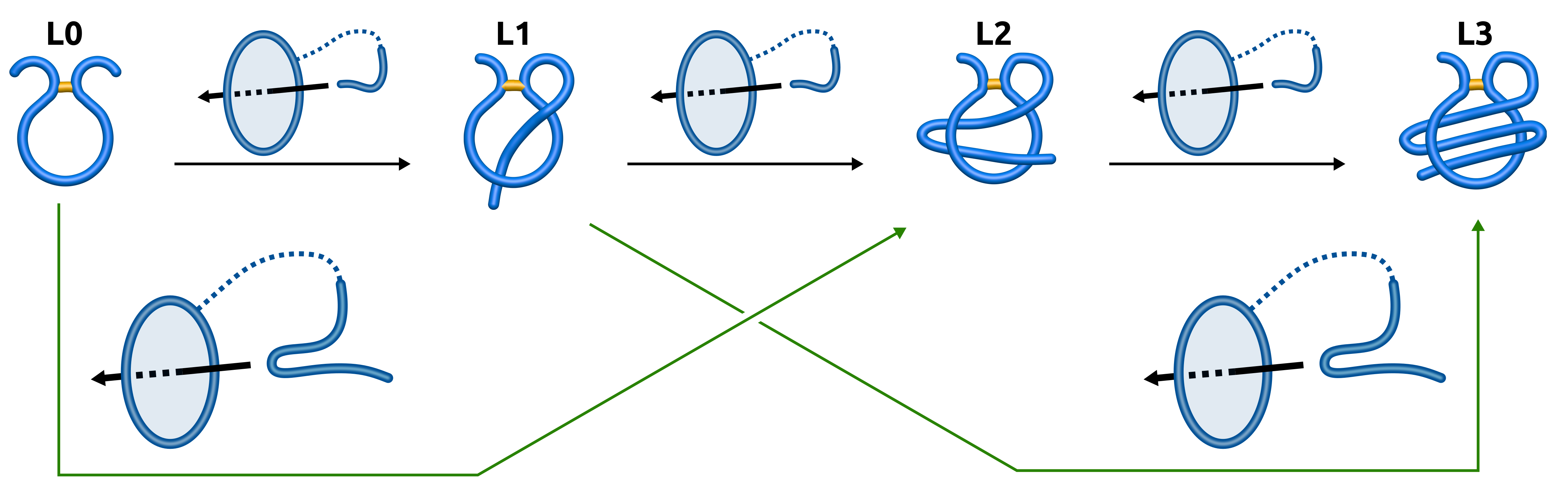
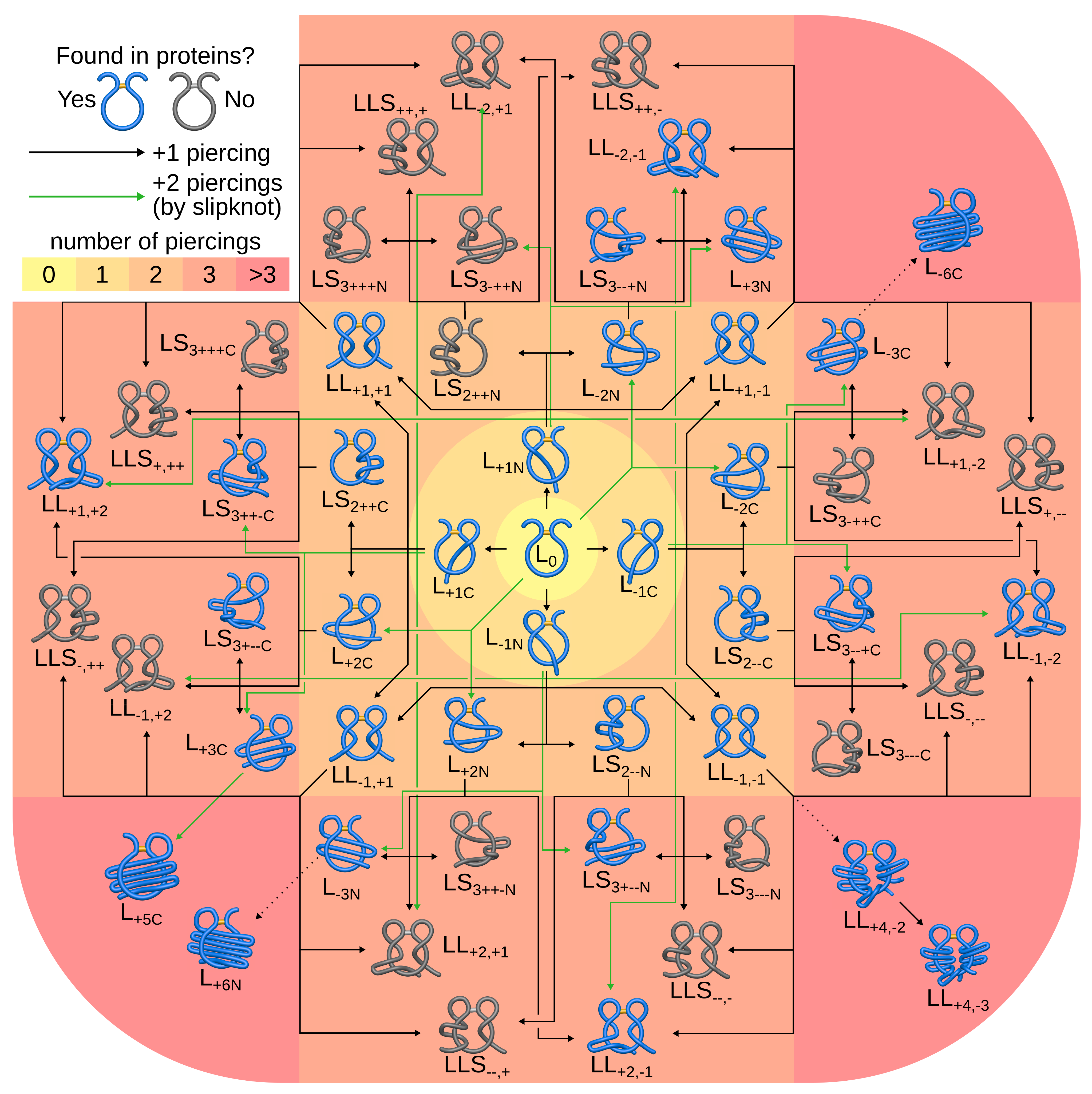
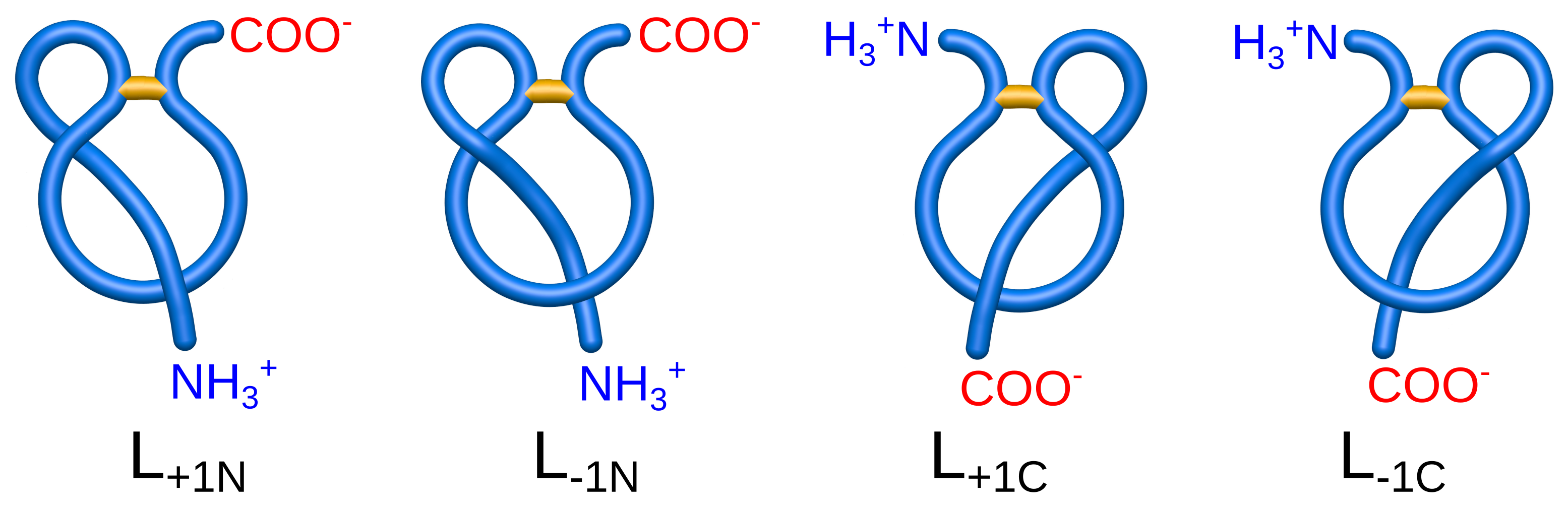
- , (, , )—with one, (two, three, six) piercing(s) of the covalent loop, where each subsequent piercing is from a different side of the loop’s surface,
- —with n piercings, but at least two consecutive ones from the same side of the loop’s surface (the tail winds around the covalent loop),
- —with i piercings performed by the N-terminal tail and j piercings by the C-terminal tail.
2. Materials and Methods
2.1. Protein Set
2.2. Lasso Type Assignment and Closed Loop Detection, Visualization
2.3. Sequence Alignment and Bridge Conservation
2.4. Protein Characteristics
3. Results
3.1. Analysis of the Entire Protein Data Bank
3.1.1. The Non-Redundant Set Is Sufficient
3.1.2. Piercings Tend to Arise from the Negative Side
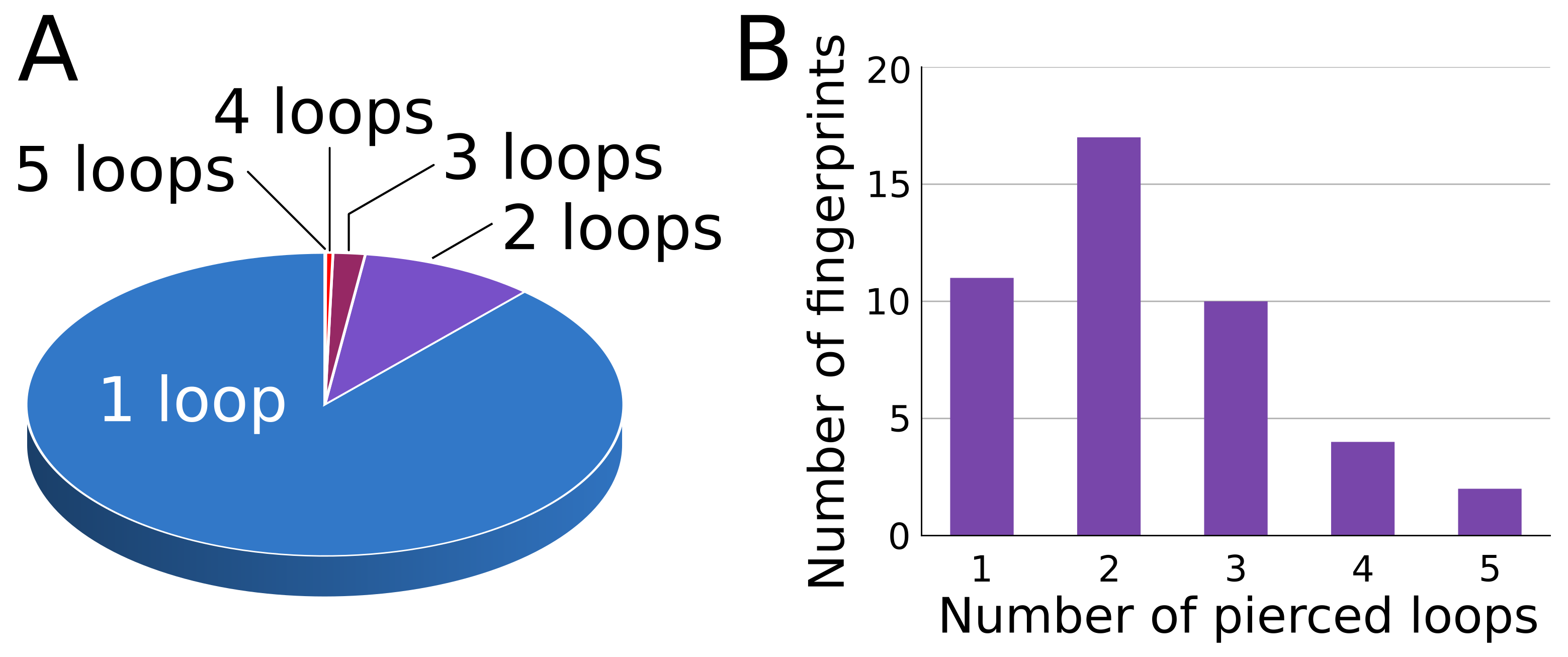
3.1.3. Lasso Fingerprint
3.1.4. Non-Disulfide Bridges
- amide based—22 chains from X miniproteins with the lasso type,
- thioester based—13 chains from 10 proteins with 5 different lasso types (, , , , , ),
- ester based—24 chains from 11 proteins with 7 different lasso types (, , , , , , ).
3.2. Topology Conservation
3.3. Complex Lasso Protein Function
3.3.1. Other Correlations
3.3.2. Stabilization of the Lasso Motif as an Indirect Proof of Functionality
3.4. Lasso Evolution
3.5. Possible Applications
4. Discussion
Significance
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| COST | European Cooperation in Science and Technology |
| EMBO | European Life Scientist Organisation |
| NIGMS | National Institute of General Medical Sciences |
| NMR | Nuclear Magnetic Resonance |
| PDB | Protein Data Bank |
| PFAM | Protein Families (database) |
| RCSB | Research Collaboratory for Structural Bioinformatics |
| UCSF | University of California, San Francisco |
References
- Taylor, W.R. A deeply knotted protein structure and how it might fold. Nature 2000, 406, 916–919. [Google Scholar] [CrossRef]
- Bölinger, D.; Sułkowska, J.I.; Hsu, H.P.; Mirny, L.A.; Kardar, M.; Onuchic, J.N.; Virnau, P. A Stevedore’s protein knot. PLoS Comput. Biol. 2010, 6, e1000731. [Google Scholar] [CrossRef] [Green Version]
- Taylor, W.R. Protein knots and fold complexity: Some new twists. Comput. Biol. Chem. 2007, 31, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Yeates, T.O.; Norcross, T.S.; King, N.P. Knotted and topologically complex proteins as models for studying folding and stability. Curr. Opin. Chem. Biol. 2007, 11, 595–603. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.Z.; Dawson, P.E. Design and synthesis of a protein catenane. Angew. Chem. 2001, 113, 3737–3739. [Google Scholar] [CrossRef]
- Boutz, D.R.; Cascio, D.; Whitelegge, J.; Perry, L.J.; Yeates, T.O. Discovery of a thermophilic protein complex stabilized by topologically interlinked chains. J. Mol. Biol. 2007, 368, 1332–1344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helgstrand, C.; Wikoff, W.R.; Duda, R.L.; Hendrix, R.W.; Johnson, J.E.; Liljas, L. The refined structure of a protein catenane: The HK97 bacteriophage capsid at 3.44 Å resolution. J. Mol. Biol. 2003, 334, 885–899. [Google Scholar] [CrossRef]
- Dabrowski-Tumanski, P.; Goundaroulis, D.; Stasiak, A.; Sulkowska, J.I. θ-curves in proteins. arXiv 2019, arXiv:1908.05919. [Google Scholar]
- McDonald, N.Q.; Hendrickson, W.A. A structural superfamily of growth factors containing a cystine knot motif. Cell 1993, 73, 421–424. [Google Scholar] [CrossRef]
- Murray-Rust, J.; McDonald, N.Q.; Blundell, T.L.; Hosang, M.; Oefner, C.; Winkler, F.; Bradshaw, R.A. Topological similarities in TGF-β2, PDGF-BB and NGF define a superfamily of polypeptide growth factors. Structure 1993, 1, 153–159. [Google Scholar] [CrossRef]
- Craik, D.J.; Daly, N.L.; Waine, C. The cystine knot motif in toxins and implications for drug design. Toxicon 2001, 39, 43–60. [Google Scholar] [CrossRef]
- Postic, G.; Gracy, J.; Périn, C.; Chiche, L.; Gelly, J.C. KNOTTIN: The database of inhibitor cystine knot scaffold after 10 years, toward a systematic structure modeling. Nucleic Acids Res. 2017, 46, D454–D458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dabrowski-Tumanski, P.; Rubach, P.; Goundaroulis, D.; Dorier, J.; Sułkowski, P.; Millett, K.C.; Rawdon, E.J.; Stasiak, A.; Sulkowska, J.I. KnotProt 2.0: A database of proteins with knots and other entangled structures. Nucleic Acids Res. 2018, 47, D367–D375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blond, A.; Péduzzi, J.; Goulard, C.; Chiuchiolo, M.J.; Barthélémy, M.; Prigent, Y.; Salomón, R.A.; Farías, R.N.; Moreno, F.; Rebuffat, S. The cyclic structure of microcin J25, a 21-residue peptide antibiotic from Escherichia coli. Eur. J. Biochem. 1999, 259, 747–756. [Google Scholar] [CrossRef] [PubMed]
- Rosengren, K.J.; Clark, R.J.; Daly, N.L.; Göransson, U.; Jones, A.; Craik, D.J. Microcin J25 has a threaded sidechain-to-backbone ring structure and not a head-to-tail cyclized backbone. J. Am. Chem. Soc. 2003, 125, 12464–12474. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, M.; Hegemann, J.D.; Xie, X.; Marahiel, M.A. The astexin-1 lasso peptides: Biosynthesis, stability, and structural studies. Chem. Biol. 2013, 20, 558–569. [Google Scholar] [CrossRef] [Green Version]
- Hegemann, J.D.; Zimmermann, M.; Xie, X.; Marahiel, M.A. Lasso peptides: An intriguing class of bacterial natural products. Accounts Chem. Res. 2015, 48, 1909–1919. [Google Scholar] [CrossRef]
- Li, Y.; Zirah, S.; Rebuffat, S. Lasso Peptide Bioengineering and Bioprospecting. In Lasso Peptides; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Martin-Gómez, H.; Tulla-Puche, J. Lasso peptides: Chemical approaches and structural elucidation. Org. Biomol. Chem. 2018, 16, 5065–5080. [Google Scholar] [CrossRef] [PubMed]
- Frechet, D.; Guitton, J.; Herman, F.; Faucher, D.; Helynck, G.; Monegier du Sorbier, B.; Ridoux, J.; James-Surcouf, E.; Vuilhorgne, M. Solution structure of RP 71955, a new 21 amino acid tricyclic peptide active against HIV-1 virus. Biochemistry 1994, 33, 42–50. [Google Scholar] [CrossRef]
- Knappe, T.A.; Manzenrieder, F.; Mas-Moruno, C.; Linne, U.; Sasse, F.; Kessler, H.; Xie, X.; Marahiel, M.A. Introducing lasso peptides as molecular scaffolds for drug design: Engineering of an integrin antagonist. Angew. Chem. Int. Ed. 2011, 50, 8714–8717. [Google Scholar] [CrossRef] [Green Version]
- Dabrowski-Tumanski, P.; Jarmolinska, A.I.; Niemyska, W.; Rawdon, E.J.; Millett, K.C.; Sulkowska, J.I. LinkProt: A database collecting information about biological links. Nucleic Acids Res. 2017, 45, D243–D249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niemyska, W.; Dabrowski-Tumanski, P.; Kadlof, M.; Haglund, E.; Sułkowski, P.; Sulkowska, J.I. Complex lasso: New entangled motifs in proteins. Sci. Rep. 2016, 6, 36895. [Google Scholar] [CrossRef]
- Dabrowski-Tumanski, P.; Niemyska, W.; Pasznik, P.; Sulkowska, J.I. LassoProt: Server to analyze biopolymers with lassos. Nucleic Acid Res. 2016, 44, W383–W389. [Google Scholar] [CrossRef] [PubMed]
- Dabrowski-Tumanski, P.; Rubach, P.; Niemyska, W.; Gren, B.A.; Sulkowska, J.I. Topoly: Python package to analyze topology of polymers. Briefings Bioinform. 2019, 22, bbaa196. [Google Scholar] [CrossRef]
- Gierut, A.M.; Niemyska, W.; Dabrowski-Tumanski, P.; Sułkowski, P.; Sulkowska, J.I. PyLasso: A PyMOL plugin to identify lassos. Bioinformatics 2017, 33, 3819–3821. [Google Scholar] [CrossRef] [PubMed]
- Dabrowski-Tumanski, P.; Gren, B.; Sulkowska, J.I. Statistical properties of lasso-shape polymers and their implications for complex lasso proteins function. Polymers 2019, 11, 707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dabrowski-Tumanski, P.; Sulkowska, J.I. Topological knots and links in proteins. Proc. Natl. Acad. Sci. USA 2017, 114, 3415–3420. [Google Scholar] [CrossRef] [Green Version]
- Haglund, E. Engineering covalent loops in proteins can serve as an on/off switch to regulate threaded topologies. J. Phys. Condens. Matter 2015, 27, 354107. [Google Scholar] [CrossRef]
- Green, J.E.; Choi, J.W.; Boukai, A.; Bunimovich, Y.; Johnston-Halperin, E.; DeIonno, E.; Luo, Y.; Sheriff, B.A.; Xu, K.; Shin, Y.S.; et al. A 160-kilobit molecular electronic memory patterned at 1011 bits per square centimetre. Nature 2007, 445, 414–417. [Google Scholar] [CrossRef]
- Clavel, C.; Fournel-Marotte, K.; Coutrot, F. A pH-sensitive peptide-containing lasso molecular switch. Molecules 2013, 18, 11553–11575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schröder, H.V.; Wollschläger, J.M.; Schalley, C.A. Redox-controlled self-inclusion of a lasso-type pseudo [1] rotaxane. Chem. Commun. 2017, 53, 9218–9221. [Google Scholar] [CrossRef]
- Wu, C.; Lecavalier, P.R.; Shen, Y.X.; Gibson, H.W. Synthesis of a rotaxane via the template method. Chem. Mater. 1991, 3, 569–572. [Google Scholar] [CrossRef]
- Hübner, G.M.; Gläser, J.; Seel, C.; Vögtle, F. High-yielding rotaxane synthesis with an anion template. Angew. Chem. Int. Ed. 1999, 38, 383–386. [Google Scholar] [CrossRef]
- Hänni, K.D.; Leigh, D.A. The application of CuAAC ‘click’chemistry to catenane and rotaxane synthesis. Chem. Soc. Rev. 2010, 39, 1240–1251. [Google Scholar] [CrossRef] [PubMed]
- Heard, A.W.; Goldup, S.M. Synthesis of a mechanically planar chiral rotaxane ligand for enantioselective catalysis. Chem 2020, 6, 994–1006. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.D.; Link, A.J. Self-assembly of catenanes from lasso peptides. J. Am. Chem. Soc. 2016, 138, 14214–14217. [Google Scholar] [CrossRef] [Green Version]
- Schröder, H.V.; Zhang, Y.; Link, A.J. Dynamic covalent self-assembly of mechanically interlocked molecules solely made from peptides. Nat. Chem. 2021, 13, 850–857. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Sułkowska, J.I.; Rawdon, E.J.; Millett, K.C.; Onuchic, J.N.; Stasiak, A. Conservation of complex knotting and slipknotting patterns in proteins. Proc. Natl. Acad. Sci. USA 2012, 109, E1715–E1723. [Google Scholar] [CrossRef] [Green Version]
- Bayro, M.J.; Mukhopadhyay, J.; Swapna, G.; Huang, J.Y.; Ma, L.C.; Sineva, E.; Dawson, P.E.; Montelione, G.T.; Ebright, R.H. Structure of antibacterial peptide microcin J25: A 21-residue lariat protoknot. J. Am. Chem. Soc. 2003, 125, 12382–12383. [Google Scholar] [CrossRef] [PubMed]
- Hashizume, R.; Maki, Y.; Mizutani, K.; Takahashi, N.; Matsubara, H.; Sugita, A.; Sato, K.; Yamaguchi, S.; Mikami, B. Crystal structures of protein glutaminase and its pro forms converted into enzyme-substrate complex. J. Biol. Chem. 2011, 286, 38691–38702. [Google Scholar] [CrossRef] [Green Version]
- Chung, I.Y.W.; Paetzel, M. Crystal Structure of a Viral Protease Intramolecular Acyl-enzyme Complex Insights into cis-Cleavage at the VP4/VP3 Junction of Tellina Birnavirus. J. Biol. Chem. 2011, 286, 12475–12482. [Google Scholar] [CrossRef] [Green Version]
- Thangudu, R.R.; Manoharan, M.; Srinivasan, N.; Cadet, F.; Sowdhamini, R.; Offmann, B. Analysis on conservation of disulphide bonds and their structural features in homologous protein domain families. BMC Struct. Biol. 2008, 8, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haglund, E.; Sułkowska, J.I.; He, Z.; Feng, G.S.; Jennings, P.A.; Onuchic, J.N. The unique cysteine knot regulates the pleotropic hormone leptin. PLoS ONE 2012, 7, e45654. [Google Scholar] [CrossRef] [Green Version]
- Adelman, K.; Yuzenkova, J.; La Porta, A.; Zenkin, N.; Lee, J.; Lis, J.T.; Borukhov, S.; Wang, M.D.; Severinov, K. Molecular mechanism of transcription inhibition by peptide antibiotic Microcin J25. Mol. Cell 2004, 14, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Niemyska, W.; Millett, K.C.; Sulkowska, J.I. GLN—A method to reveal unique properties of lasso type topology in proteins. Sci. Rep. 2020, 10, 15186. [Google Scholar] [CrossRef] [PubMed]
- Sułkowska, J.I.; Sułkowski, P.; Onuchic, J. Dodging the crisis of folding proteins with knots. Proc. Natl. Acad. Sci. USA 2009, 106, 3119–3124. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.; Van Vlijmen, H.W.; Singh, J. A classification of disulfide patterns and its relationship to protein structure and function. Protein Sci. 2004, 13, 2045–2058. [Google Scholar] [CrossRef] [PubMed]
- Mas, J.M.; Aloy, P.; Marti-Renom, M.A.; Oliva, B.; de Llorens, R.; Aviles, F.X.; Querol, E. Classification of protein disulphide-bridge topologies. J. Comput.-Aided Mol. Des. 2001, 15, 477–487. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | # Loops | # Repr. Loops | Class | # Loops | # Repr. Loops | Class | # Loops | # Repr. Loops |
|---|---|---|---|---|---|---|---|---|
| 1459 426 2202 546 504 56 180 31 88 5 329 11 3 4 1 | 244 85 204 127 60 12 21 8 7 1 24 3 0 2 1 | 60 95 195 4 3 1 12 9 | 8 4 8 2 2 0 0 3 | 11 34 43 1 1 3 3 1 2 1 3 3 | 4 6 6 1 1 1 1 1 0 0 1 1 | |||
| Sum | 7845 | 799 | Sum | 379 | 27 | Sum | 106 | 23 |
| Total | All loops—6330 All chains—5559 | Loops in representative set—849 Chains in representative set—732 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Greń, B.A.; Dabrowski-Tumanski, P.; Niemyska, W.; Sulkowska, J.I. Lasso Proteins—Unifying Cysteine Knots and Miniproteins. Polymers 2021, 13, 3988. https://doi.org/10.3390/polym13223988
Greń BA, Dabrowski-Tumanski P, Niemyska W, Sulkowska JI. Lasso Proteins—Unifying Cysteine Knots and Miniproteins. Polymers. 2021; 13(22):3988. https://doi.org/10.3390/polym13223988
Chicago/Turabian StyleGreń, Bartosz Ambroży, Pawel Dabrowski-Tumanski, Wanda Niemyska, and Joanna Ida Sulkowska. 2021. "Lasso Proteins—Unifying Cysteine Knots and Miniproteins" Polymers 13, no. 22: 3988. https://doi.org/10.3390/polym13223988