
Biodegradation of Polystyrene by Tenebrio molitor, Galleria mellonella, and Zophobas atratus Larvae and Comparison of Their Degradation Effects


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Test Materials
2.2. Feeding Tests
2.3. Collection and Characterization of Frass
2.4. Analysis of Gut Microbial Community
2.5. Determination of Degradation Products
2.6. Statistical Analysis
3. Results and Discussion
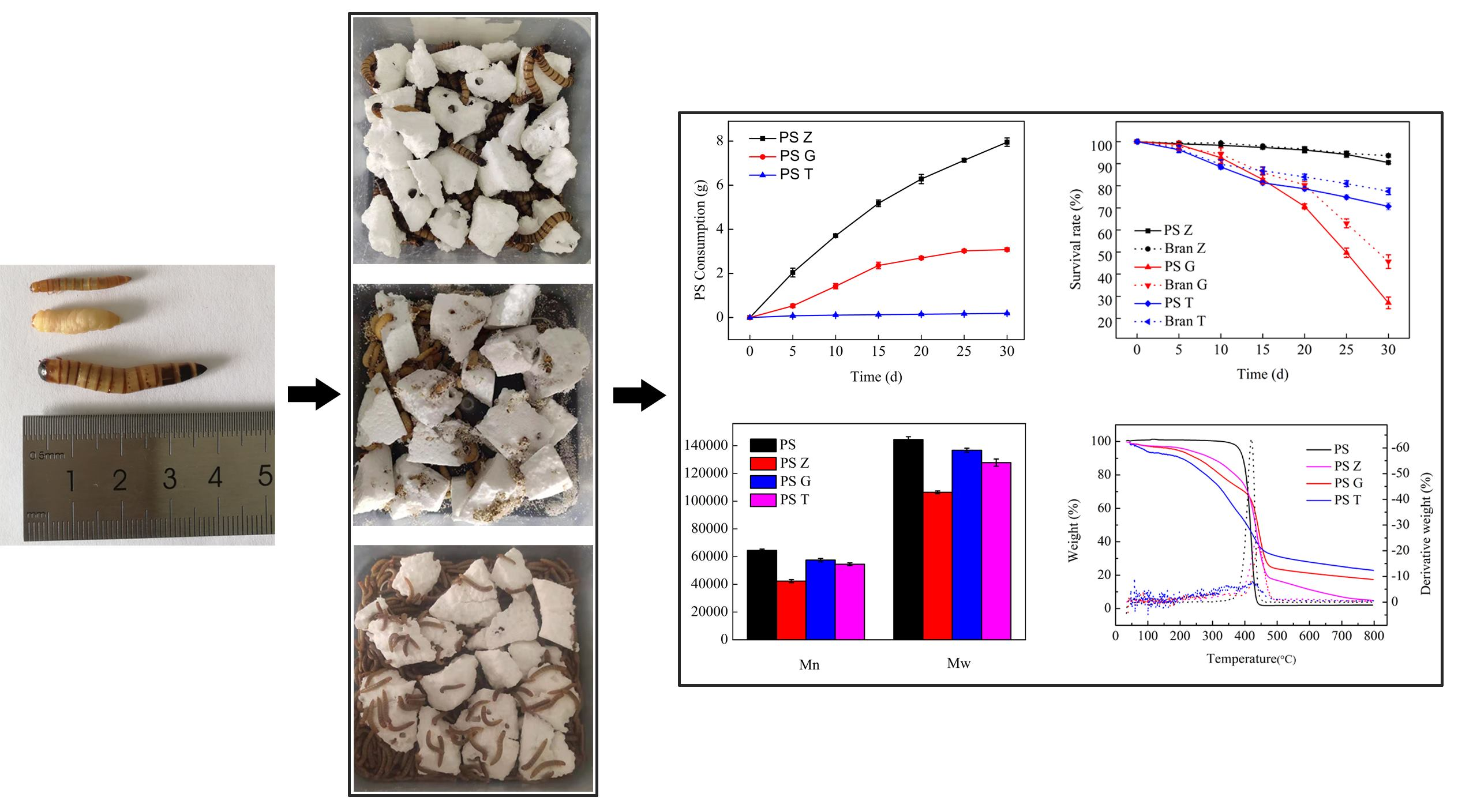
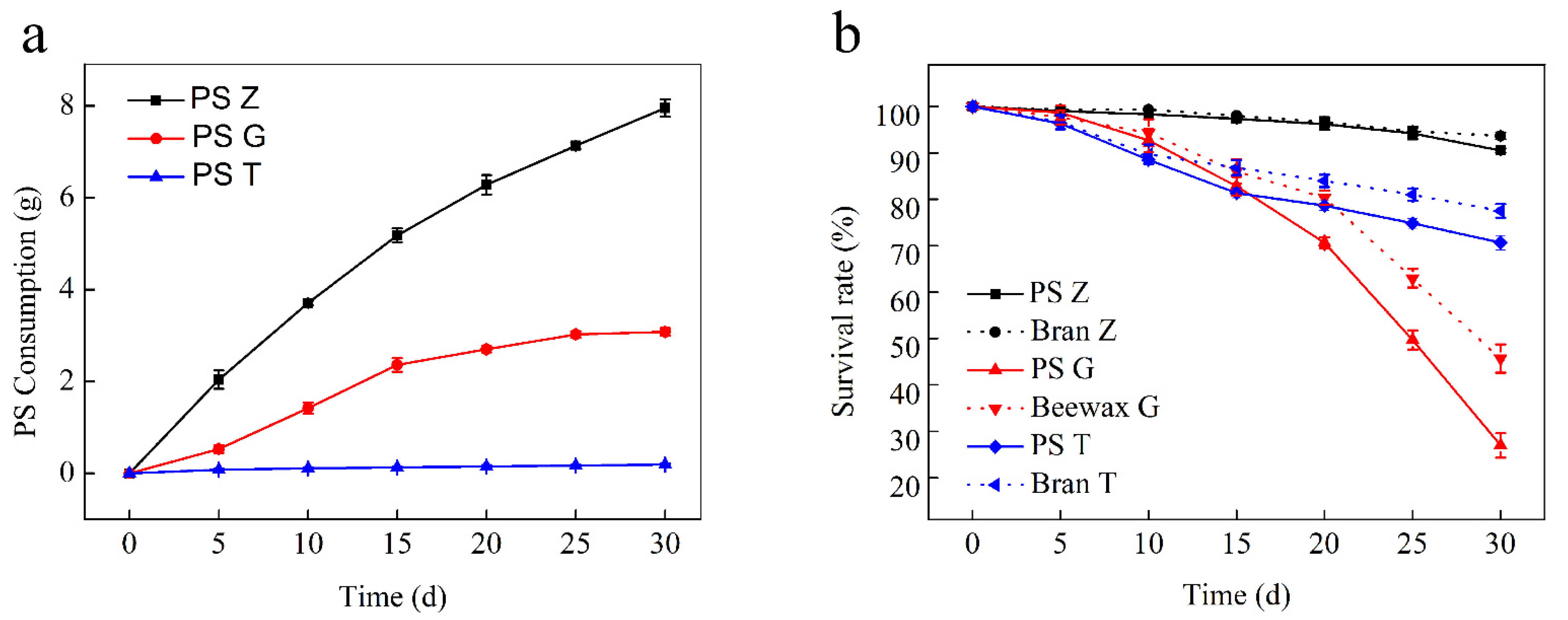
3.1. Changes in PS Consumption, Larvae Weight, and Survival Rates
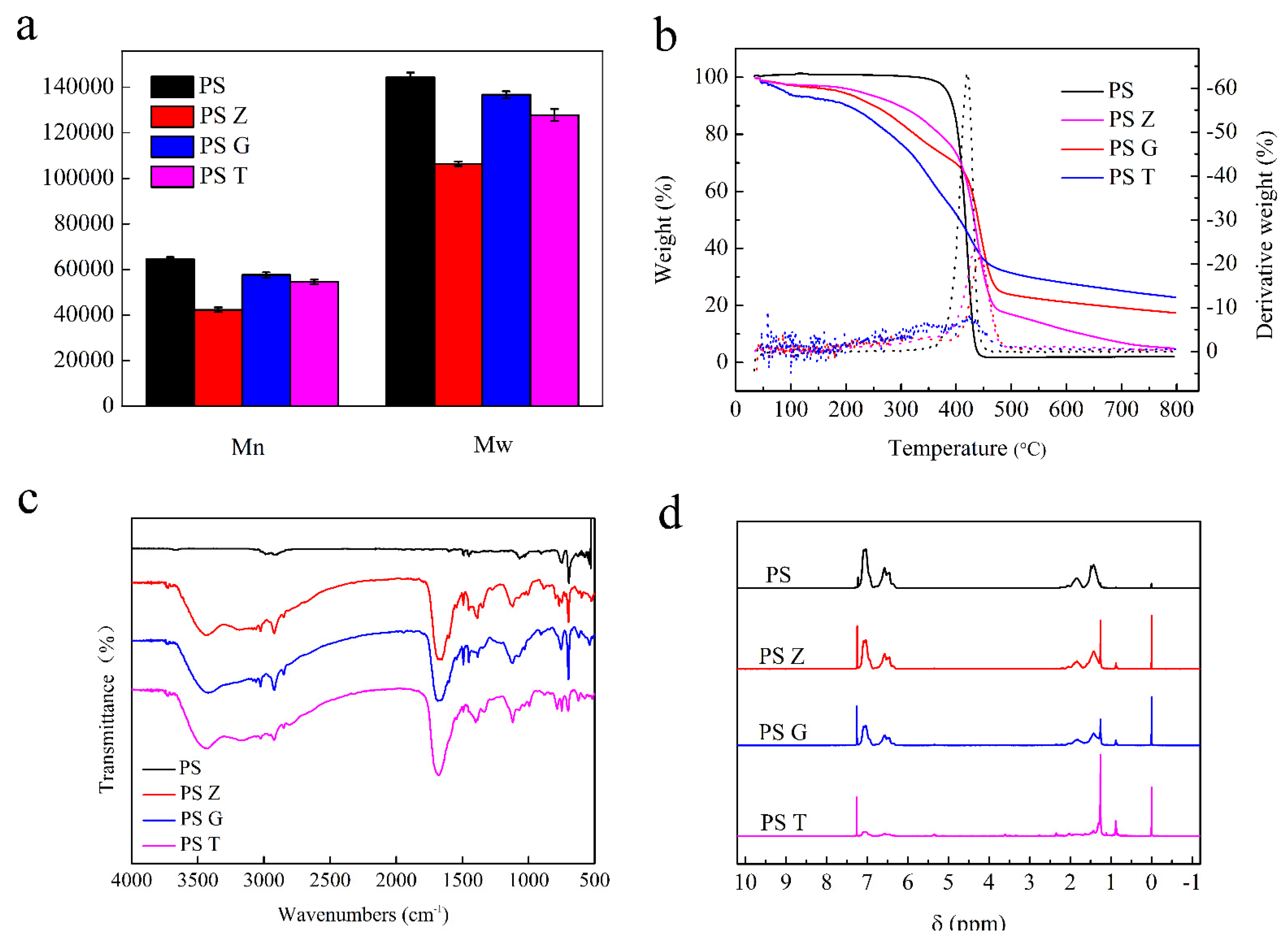
3.2. Evidence and Differences of Biodegradation
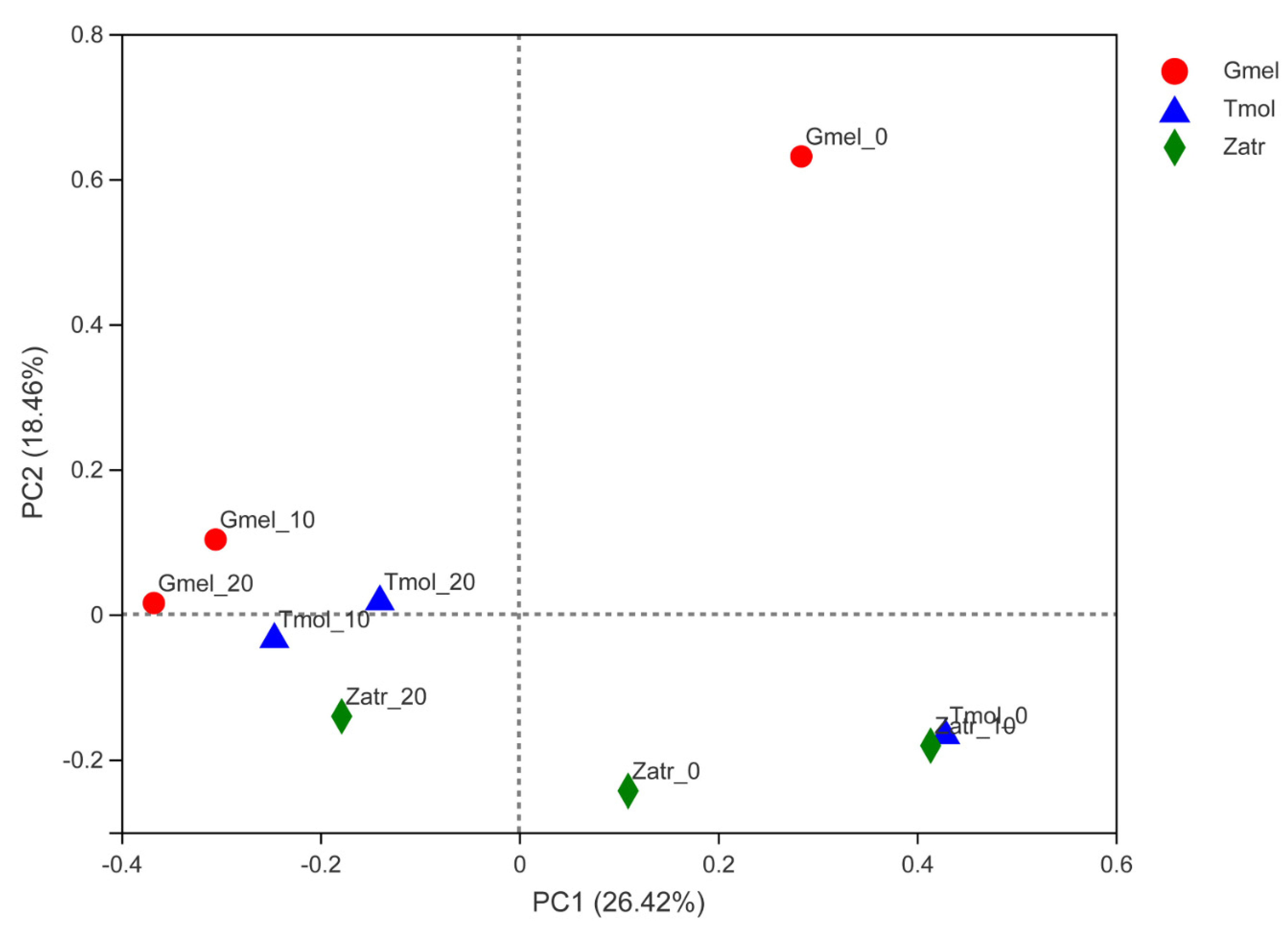
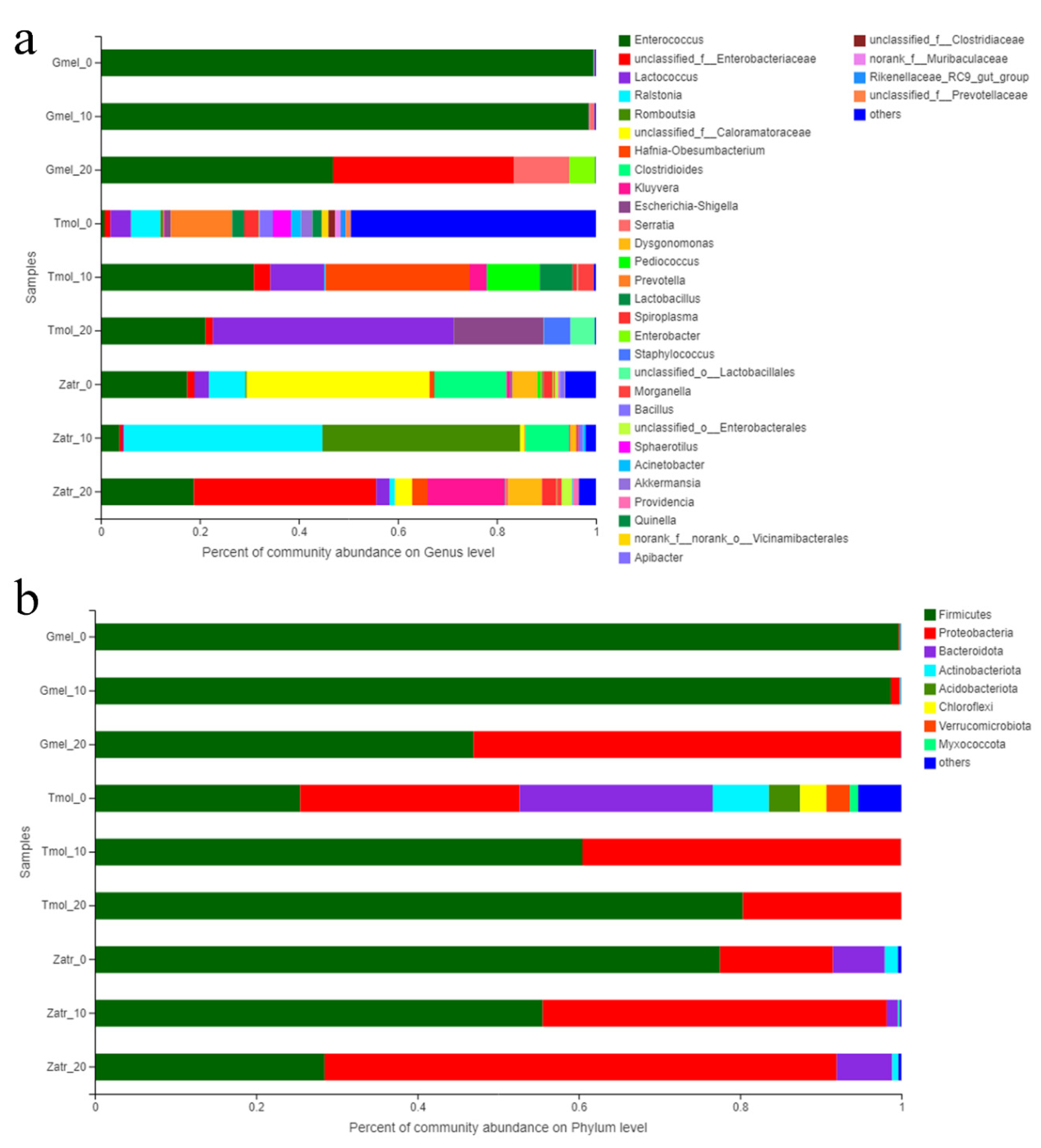
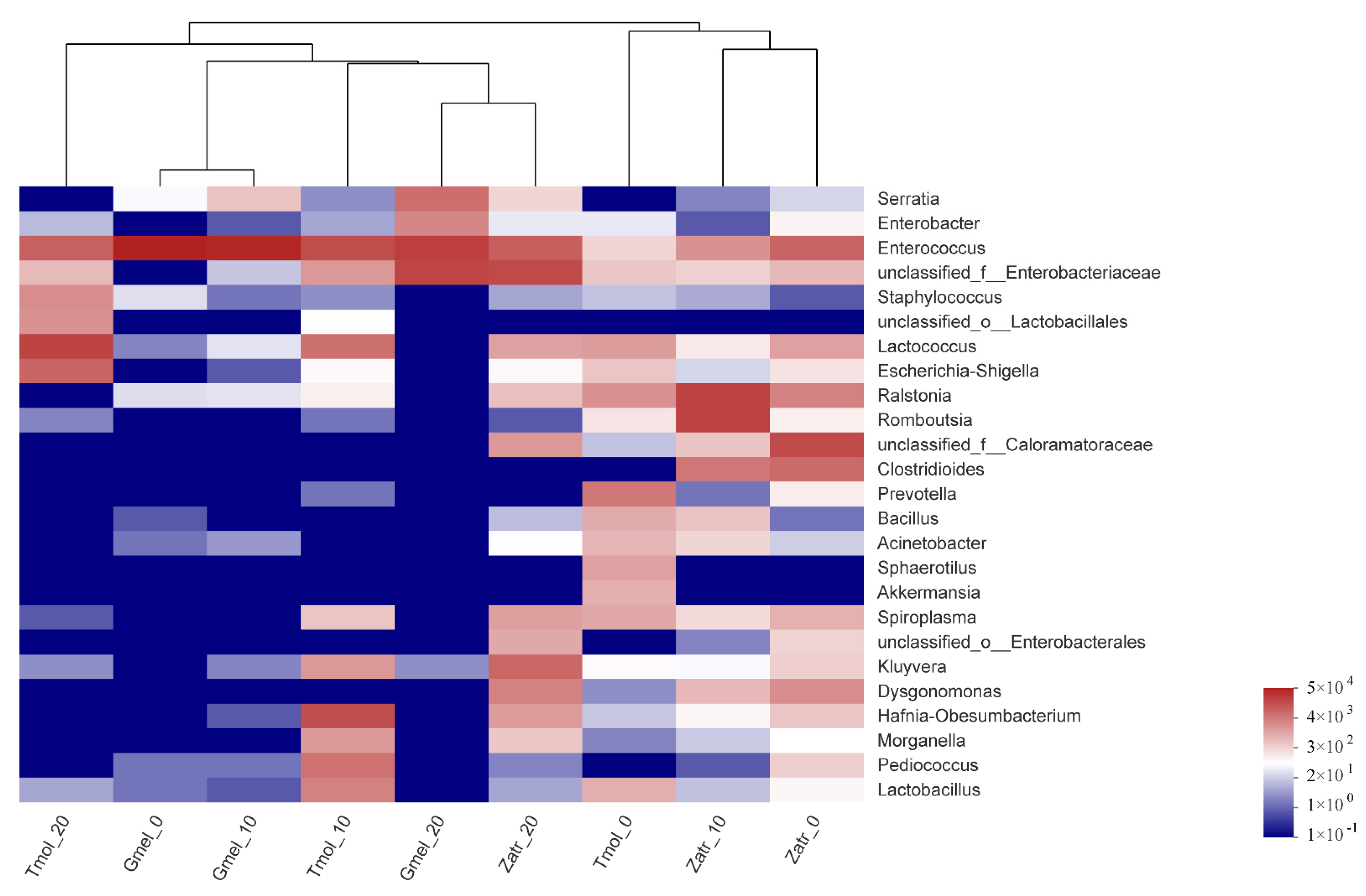
3.3. Comparison of Gut Microbial Diversity
3.4. Analysis of Degradation Products
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mohanan, N.; Montazer, Z.; Sharma, P.K.; Levin, D.B. Microbial and Enzymatic Degradation of Synthetic Plastics. Front. Microbiol. 2020, 11, 2837. [Google Scholar] [CrossRef]
- Veerasingam, S.; Vethamony, P.; Aboobacker, V.M.; Giraldes, A.E.; Al Khayat, J.A. Factors influencing the vertical distribution of microplastics in the beach sediments around the Ras Rakan Island, Qatar. Environ. Sci. Pollut. Res. 2021, 28, 34259–34268. [Google Scholar] [CrossRef]
- Chamas, A.; Moon, H.; Zheng, J.J.; Qiu, Y.; Tabassum, T.; Jang, J.H.; Abu-Omar, M.; Scott, S.L.; Suh, S. Degradation Rates of Plastics in the Environment. ACS Sustain. Chem. Eng. 2020, 8, 3494–3511. [Google Scholar] [CrossRef] [Green Version]
- Carmen, S. Microbial capability for the degradation of chemical additives present in petroleum-based plastic products: A review on current status and perspectives. J. Hazard. Mater. 2021, 402, 123534. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, M.; Salazar Robles, G.; José, M.; Cunill, J. Aiding the Environment: Microorganisms for the Degradation of Plastic in Soil and Water. Indian J. Appl. Microbiol. 2019, 22, 19–37. [Google Scholar]
- Krueger, M.C.; Harms, H.; Schlosser, D. Prospects for microbiological solutions to environmental pollution with plastics. Appl. Microbiol. Biotechnol. 2015, 99, 8857–8874. [Google Scholar] [CrossRef] [PubMed]
- Malachová, K.; Novotn, Č.; Adamus, G.; Lotti, N.; Fava, F. Ability of Trichoderma hamatum Isolated from Plastics-Polluted Environments to Attack Petroleum-Based, Synthetic Polymer Films. Processes 2020, 8, 467. [Google Scholar] [CrossRef] [Green Version]
- Mor, R.; Sivan, A. Biofilm formation and partial biodegradation of polystyrene by the actinomycete Rhodococcus ruber: Biodegradation of polystyrene. Biodegradation 2008, 19, 851–858. [Google Scholar] [CrossRef]
- Sekhar, V.C.; Nampoothiri, K.M.; Mohan, A.J.; Nair, N.R.; Bhaskar, T.; Pandey, A. Microbial degradation of high impact polystyrene (HIPS), an e-plastic with decabromodiphenyl oxide and antimony trioxide. J. Hazard. Mater. 2016, 318, 347–354. [Google Scholar] [CrossRef]
- Riudavets, J.; Salas, I.; Pons, M.J. Damage characteristics produced by insect pests in packaging film. J. Stored Prod. Res. 2007, 43, 564–570. [Google Scholar] [CrossRef]
- Yang, J.; Yang, Y.; Wu, W.M.; Zhao, J.; Jiang, L. Evidence of Polyethylene Biodegradation by Bacterial Strains from the Guts of Plastic-Eating Waxworms. Environ. Sci. Technol. 2014, 48, 13776–13784. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yang, J.; Wu, W.M.; Zhao, J.; Song, Y.L.; Gao, L.C.; Yang, R.F.; Jiang, L. Biodegradation and Mineralization of Polystyrene by Plastic-Eating Mealworms: Part 1. Chemical and Physical Characterization and Isotopic Tests. Environ. Sci. Technol. 2015, 49, 12080–12086. [Google Scholar] [CrossRef]
- Yang, S.S.; Brandon, A.M.; Flanagan, J.C.A.; Yang, J.; Ning, D.; Cai, S.Y.; Fan, H.Q.; Wang, Z.Y.; Ren, J.; Benbow, E.; et al. Biodegradation of polystyrene wastes in yellow mealworms (larvae of Tenebrio molitor Linnaeus): Factors affecting biodegradation rates and the ability of polystyrene-fed larvae to complete their life cycle. Chemosphere 2018, 191, 979–989. [Google Scholar] [CrossRef]
- Peng, B.Y.; Su, Y.M.; Chen, Z.B.; Chen, J.B.; Zhou, X.F.; Benbow, M.E.; Criddle, C.S.; Wu, W.M.; Zhang, Y.L. Biodegradation of Polystyrene by Dark (Tenebrio obscurus) and Yellow (Tenebrio molitor) Mealworms (Coleoptera: Tenebrionidae). Environ. Sci. Technol. 2019, 53, 5256–5265. [Google Scholar]
- Bryan, J.C.; Grove, H.; Elebute, O.; Villanueva, S.; LeMoine, C. Role of the intestinal microbiome in low-density polyethylene degradation by caterpillar larvae of the greater wax moth, Galleria mellonella. Proc. R. Soc. B 2020, 287, 1–8. [Google Scholar]
- Lou, Y.; Ekaterina, P.; Yang, S.S.; Lu, B.Y.; Liu, B.F.; Ren, N.; Corvini, P.; Xing, D.F. Bio-degradation of Polyethylene and Polystyrene by Greater Wax Moth Larvae (Galleria mellonella L.) and the Effect of Co-diet Supplementation on the Core Gut Microbiome. Environ. Sci. Technol. 2020, 54, 2821–2831. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Xin, X.; Shi, X.; Zhang, Y. A polystyrene-degrading Acinetobacter bacterium isolated from the larvae of Tribolium castaneum. Sci. Total Environ. 2020, 726, 138564. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wang, J.; Xia, M. Biodegradation and mineralization of polystyrene by plastic-eating superworms Zophobas atratus. Sci. Total Environ. 2020, 708, 1352331–1352337. [Google Scholar]
- Atiq, N.; Ahmed, S.; Ali, M.I.; Andleeb, S.; Ahmad, B.; Robson, G. Isolation and identification of polystyrene biodegrading bacteria from soil. Afr. J. Microbiol. Res. 2011, 4, 1537–1541. [Google Scholar]
- Kaplan, D.L.; Hartenstein, R.; Sutter, J. Biodegradation of polystyrene, poly(metnyl methacrylate), and phenol formaldehyde. Appl. Environ. Microbiol. 1979, 38, 551–553. [Google Scholar] [CrossRef] [Green Version]
- Skariyachan, S.; Megha, M.; Kini, M.N.; Mukund, K.M.; Rizvi, A.; Vasist, K. Selection and screening of microbial consortia for efficient and ecofriendly degradation of plastic garbage collected from urban and rural areas of Bangalore, India. Environ. Monit. Assess. 2015, 187, 4174. [Google Scholar] [CrossRef] [PubMed]
- Brandon, A.M.; Garcia, A.M.; Khlystov, N.A.; Wu, W.M.; Criddle, C.S. Enhanced Bioavailability and Microbial Biodegradation of Polystyrene in an Enrichment Derived from the Gut Microbiome of Tenebrio molitor (Mealworm Larvae). Environ. Sci. Technol. 2021, 55, 2027–2036. [Google Scholar] [CrossRef] [PubMed]
- Peng, B.Y.; Li, Y.R.; Fan, R.; Chen, Z.B.; Wu, W.M. Biodegradation of low-density polyethylene and polystyrene in superworms, larvae of Zophobas atratus (Coleoptera: Tenebrionidae): Broad and limited extent depolymerization. Environ. Pollut. 2020, 266, 115206. [Google Scholar] [CrossRef]
- Farrelly, T.A.; Shaw, I.C. Polystyrene as Hazardous Household Waste. In Household Hazardous Waste Management; Mmereki, D., Ed.; IntechOpen: London, UK, 2017; pp. 45–60. [Google Scholar]
- Machado, N.C.; De Jesus, L.A.; Pinto, P.S.; De Paula, F.G.; Alves, M.O.; Mendes, K.H.; Mambrini, R.V.; Barrreda, D.; Rocha, V.; Santamaría, R.; et al. Waste-polystyrene foams-derived magnetic carbon material for adsorption and redox supercapacitor applications. J. Clean. Prod. 2021, 313, 127903. [Google Scholar] [CrossRef]
- Albertsson, A.C.; Erlandsson, B.; Hakkarainen, M.; Karlsson, S. Molecular Weight Changes and Polymeric Matrix Changes Correlated with the Formation of Degradation Products in Biodegraded Polyethylene. J. Environ. Polym. Degrad. 1998, 6, 187–195. [Google Scholar] [CrossRef]
- Ali, M.I.; Ahmed, S.; Robson, G.; Javed, I.; Hameed, A. Isolation and molecular characterization of polyvinyl chloride (PVC) plastic degrading fungal isolates. J. Basic Microbiol. 2014, 54, 18–27. [Google Scholar] [CrossRef]
- Brandon, A.M.; Gao, S.H.; Tian, R.M.; Ning, D.L.; Yang, S.S.; Zhou, J.Z.; Wu, W.M.; Criddle, C.S. Biodegradation of Polyethylene and Plastic Mixtures in Mealworms (Larvae of Tenebrio molitor) and Effects on the Gut Microbiome. Environ. Sci. Technol. 2018, 52, 6526–6533. [Google Scholar] [CrossRef]
- Tinker, K.A.; Ottesen, E.A. The Core Gut Microbiome of the American Cockroach, Periplaneta americana, Is Stable and Resilient to Dietary Shifts. Appl. Environ. Microbiol. 2016, 82, 6603–6610. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, Y.L. Investigation of Gut-Associated Bacteria in Tenebrio molitor (Coleoptera: Tenebrionidae) Larvae Using Culture-Dependent and DGGE Methods. Ann. Entomol. Soc. Am. 2015, 108, 941–949. [Google Scholar] [CrossRef]
- Yang, Y.; Yang, J.; Wu, W.M.; Zhao, J.; Song, Y.L.; Gao, L.C.; Yang, R.F.; Jiang, L. Biodegradation and Mineralization of Polystyrene by Plastic-Eating Mealworms: Part 2. Role of Gut Microorganisms. Environ. Sci. Technol. 2015, 49, 12087–12093. [Google Scholar]
- Woo, S.; Song, I.; Cha, H.J. Fast and Facile Biodegradation of Polystyrene by the Gut Microbial Flora of Plesiophthalmus davidis Larvae. Appl. Environ. Microbiol. 2020, 86, e01361–e013620. [Google Scholar] [CrossRef] [PubMed]
- Kong, H.G.; Kim, H.H.; Chung, J.-H.; Jun, J.H.; Soohyun, L.; Kim, H.-M.; Jeon, S.; Park, S.G.; Bhak, J.; Ryu, C.-M. The Galleria mellonella Hologenome Supports Microbiota-Independent Metabolism of Long-Chain Hydrocarbon Beeswax. Cell Rep. 2019, 26, 2451–2464. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Larvae | Initial Weight/Larva (g) | Feed | Weight Change at the End of the Test (%) | Survival Rate (%) | mg PS/Larva/d |
|---|---|---|---|---|---|
| Superworm | 0.86 ± 0.021 | PS | −51.67 ± 1.15 | 90.5 ± 0.5 | 2.78 ± 0.060 a |
| Bran | +12.11 ± 3.37 | 93.67 ± 1.53 | |||
| Greater wax moth | 0.21 ± 0.015 | PS | −43.61 ± 4.67 | 27 ± 2.65 | 1.57 ± 0.066 b |
| Beeswax | +18.89 ± 2.12 | 45.67 ± 3.06 | |||
| Yellow mealworm | 0.08 ± 0.015 | PS | −17.06 ± 5.28 | 75.5 ± 7.4 | 0.07 ± 0.009 c |
| Bran | +25.92 ± 1.84 | 80.83 ± 7.11 |
| Sample | Size | OTUs | Shannon | Simpson | Ace | Chao | Coverage |
|---|---|---|---|---|---|---|---|
| Gmel_0 | 52,347 | 35 | 0.180129 | 0.931893 | 50.41519 | 43.75 | 0.999543 |
| Gmel_10 | 44,132 | 43 | 0.224694 | 0.925592 | 72.07289 | 60.1 | 0.999421 |
| Gmel_20 | 48,381 | 13 | 1.336777 | 0.319691 | 13.375 | 13 | 0.99997 |
| Tmol_0 | 36,796 | 1268 | 5.892504 | 0.009599 | 1269.646 | 1268.35 | 0.999756 |
| Tmol_10 | 47,699 | 37 | 1.964937 | 0.197299 | 60.83233 | 42.6 | 0.999756 |
| Tmol_20 | 43,830 | 27 | 1.605712 | 0.267592 | 38.36263 | 30.75 | 0.999817 |
| Zatr_0 | 41,939 | 148 | 2.399686 | 0.189472 | 179.5835 | 168.6667 | 0.999024 |
| Zatr_10 | 49,021 | 157 | 1.570013 | 0.32704 | 186.085 | 190.4762 | 0.998842 |
| Zatr_20 | 45,519 | 161 | 2.337035 | 0.176703 | 199.1793 | 191.4412 | 0.998598 |
| Larvae | Sample | Chemical Compound |
|---|---|---|
| Greater wax moth | Frass | 2-propenoic acid, behenic alcohol, benzenepropanoic acid, acetic acid, 6-tetradecanesulfonic acid |
| Gut | Hexadecanoic acid, oxalic acid, phthalic acid, benzenepropanoic acid, benzoic acid | |
| Yellow mealworm | Frass | 2-propenoic acid, behenic alcohol, benzenepropanoic acid, silicic acid, [1,2,4]triazo[1,5-a]pyrimidine-6-carboxylic acid |
| Gut | Hexadecanoic acid, 9-octadecenoic acid, z-8-methyl-9-tetradecenoic acid | |
| Superworm | Frass | 2-propenoic acid, behenic alcohol, 9-octadecenoic acid, phthalic acid, hexadecanoic acid, methoxyacetic acid |
| Gut | hexadecanoic acid, stearic acid, phthalic acid, 6-octadecenoic acid | |
| PS | phenol, heptane, toluene, o-xylene, 3-(benzylthio)acrylic acid |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, S.; Su, T.; Zhao, J.; Wang, Z. Biodegradation of Polystyrene by Tenebrio molitor, Galleria mellonella, and Zophobas atratus Larvae and Comparison of Their Degradation Effects. Polymers 2021, 13, 3539. https://doi.org/10.3390/polym13203539
Jiang S, Su T, Zhao J, Wang Z. Biodegradation of Polystyrene by Tenebrio molitor, Galleria mellonella, and Zophobas atratus Larvae and Comparison of Their Degradation Effects. Polymers. 2021; 13(20):3539. https://doi.org/10.3390/polym13203539
Chicago/Turabian StyleJiang, Shan, Tingting Su, Jingjing Zhao, and Zhanyong Wang. 2021. "Biodegradation of Polystyrene by Tenebrio molitor, Galleria mellonella, and Zophobas atratus Larvae and Comparison of Their Degradation Effects" Polymers 13, no. 20: 3539. https://doi.org/10.3390/polym13203539