
New Insights into Anthelmintic Mechanisms of Action of a Synthetic Peptide: An Ultrastructural and Nanomechanical Approach

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Peptide Design, Characterization by Bioinformatic Analysis, and Chemical Synthesis
2.2. Anthelmintic Assays
2.3. Egg Hatch Test (EHT)
2.4. Larval Development Test (LDT)
2.5. Atomic Force Microscopy (AFM) Analyses
2.6. Hemolytic Potential and Toxicity to Vero Cell Lines of RcAlb-PepIII
3. Results
3.1. Design and Bioinformatics Data of RcAlb-PepIII
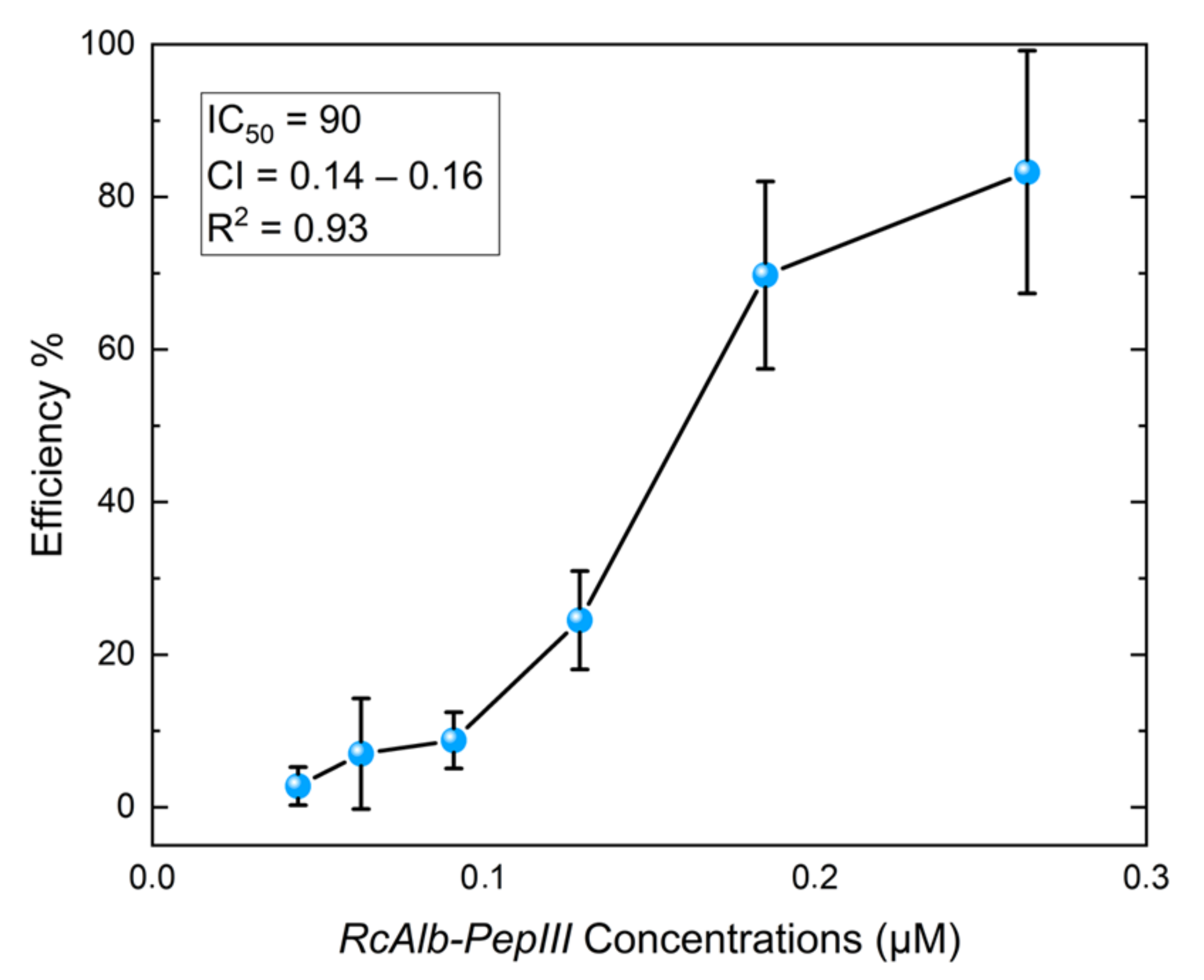
3.2. Anthelmintic Assays
3.3. Atomic Force Microscopy (AFM) Analyses
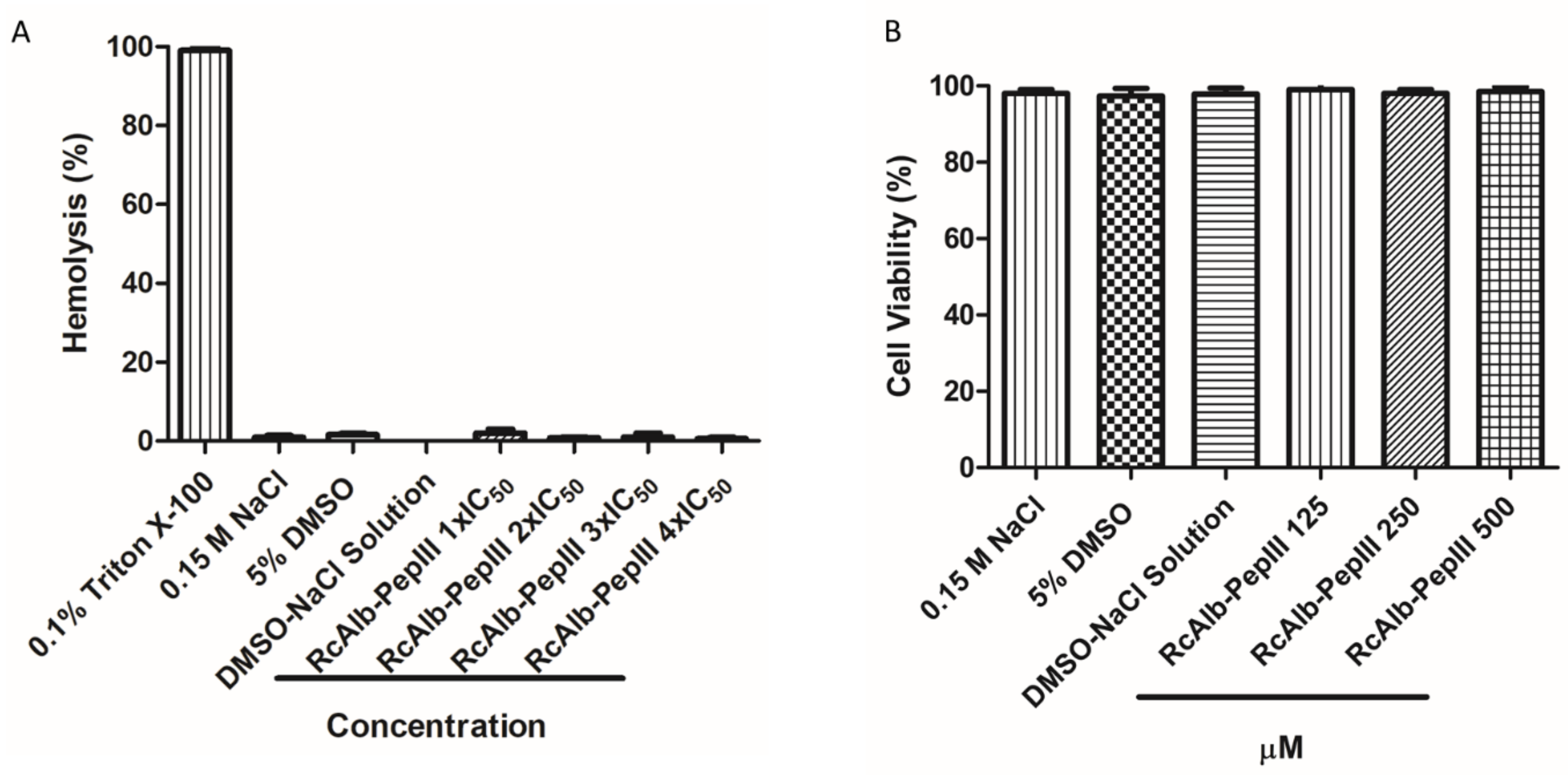
3.4. Toxicity of RcAlb-PepIII
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Ethical Approval
References
- Besier, R.B.; Kahn, L.P.; Sargison, N.D.; Van Wyk, J.A. The Pathophysiology, Ecology and Epidemiology of Haemonchus contortus Infection in Small Ruminants. Adv. Parasitol. 2016, 93, 95–143. [Google Scholar] [CrossRef] [PubMed]
- Emery, D.L.; Hunt, P.W.; Le Jambre, L.F. Haemonchus contortus: The then and now, and where to from here? Int. J. Parasitol. 2016, 46, 755–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplana, R.M.; Vidyashankarb, A.N. An inconvenient truth: Global worming and anthelmintic resistance. Vet. Parasitol. 2012, 186, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Vatta, A.F.; Lindberg, L.E. Managing anthelmintic resistance in small ruminant livestock of resource-poor farmers in South Africa. J. S. Afr. Vet. Assoc. 2006, 77, 2–8. [Google Scholar] [CrossRef] [Green Version]
- Howell, S.B.; Burke, J.M.; Miller, J.E.; Terrill, T.H.; Valencia, E.; Williams, M.J.; Williamson, L.H.; Zajac, A.M.; Kaplan, R.M. Prevalence of anthelmintic resistance on sheep and goat farms in the southeastern United States. J. Am. Vet. Med. Assoc. 2008, 233, 1913–1919. [Google Scholar] [CrossRef] [Green Version]
- Kotze, A.C.; Prichard, R.K. Anthelmintic Resistance in Haemonchus contortus. History, Mechanisms and Diagnosis. Adv. Parasitol. 2016, 93, 397–428. [Google Scholar] [CrossRef]
- Nixon, S.A.; Welz, C.; Woods, D.J.; Costa-Junior, L.; Zamanian, M.; Martin, R.J. Where are all the anthelmintics? Challenges and opportunities on the path to new anthelmintics. Int. J. Parasitol. Drugs Drug Resist. 2020, 14, 8–16. [Google Scholar] [CrossRef]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef]
- da Costa, J.P.; Cova, M.; Ferreira, R.; Vitorino, R. Antimicrobial peptides: An alternative for innovative medicines? Appl. Microbiol. Biotechnol. 2015, 99, 2023–2040. [Google Scholar] [CrossRef]
- Marr, A.K.; Gooderham, W.J.; Hancock, R.E. Antibacterial peptides for therapeutic use: Obstacles and realistic outlook. Curr. Opin. Pharmacol. 2006, 6, 468–472. [Google Scholar] [CrossRef]
- Colgrave, M.L.; Kotze, A.C.; Kopp, S.; McCarthy, J.S.; Coleman, G.T.; Craik, D.J. Anthelmintic activity of cyclotides: In vitro studies with canine and human hookworms. Acta Trop. 2009, 109, 163–166. [Google Scholar] [CrossRef]
- Colgrave, M.L.; Kotze, A.C.; Ireland, D.C.; Wang, C.K.; Craik, D.J. The anthelmintic activity of the cyclotides: Natural variants with enhanced activity. Chembiochem 2008, 9, 1939–1945. [Google Scholar] [CrossRef]
- Colgrave, M.L.; Kotze, A.C.; Huang, Y.-H.; O’Grady, J.; Simonsen, S.M.; Craik, D.J. Cyclotides: Natural, Circular Plant Peptides that Possess Significant Activity against Gastrointestinal Nematode Parasites of Sheep. Biochemistry 2008, 47, 5581–5589. [Google Scholar] [CrossRef]
- Souza, P.F.N.; Marques, L.S.M.; Oliveira, J.T.A.; Lima, P.G.; Dias, L.P.; Neto, N.A.S.; Lopes, F.E.S.; Sousa, J.S.; Silva, A.F.B.; Caneiro, R.F.; et al. Synthetic antimicrobial peptides: From choice of the best sequences to action mechanisms. Biochimie 2020, 175, 132–145. [Google Scholar] [CrossRef]
- Souza, P.F.N.; Vasconcelos, I.M.; Silva, F.D.A.; Moreno, F.B.; Monteiro-Moreira, A.C.O.; Alencar, L.M.R.; Abreu, A.S.G.; Sousa, J.S.; Oliveira, J.T.A. A 2S albumin from the seed cake of ricinus communis inhibits trypsin and has strong antibacterial activity against human pathogenic bacteria. J. Nat. Prod. 2016, 79. [Google Scholar] [CrossRef]
- Dias, L.P.; Souza, P.F.N.; Oliveira, J.T.A.; Vasconcelos, I.M.; Araújo, N.M.S.; Tilburg, M.F.V.; Guedes, M.I.F.; Carneiro, R.F.; Lopes, J.L.S.; Sousa, D.O.B. RcAlb-PepII, a synthetic small peptide bioinspired in the 2S albumin from the seed cake of Ricinus communis, is a potent antimicrobial agent against Klebsiella pneumoniae and Candida parapsilosis. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183092. [Google Scholar] [CrossRef]
- Sharma, A.; Singla, D.; Rashid, M.; Raghava, G.P.S. Designing of peptides with desired half-life in intestine-like environment. BMC Bioinform. 2014, 15, 282. [Google Scholar] [CrossRef] [Green Version]
- Thévenet, P.; Shen, Y.; Maupetit, J.; Guyon, F.; Derreumaux, P.; Tufféry, P. PEP-FOLD: An updated de novo structure prediction server for both linear and disulfide bonded cyclic peptides. Nucleic Acids Res. 2012, 40. [Google Scholar] [CrossRef] [Green Version]
- Coles, G.C.; Bauerb, C.; Borgsteedec, F.H.; Geertsd, S.; Kleie, T.R.; Taylora, M.A.; Wallerf, P.J. World Association for the Advancement of Veterinary Parasitology (W.A.A.V.P.) methods for the detection of anthelmintic resistance in nematodes of veterinary importance. Vet. Parasitol. 1992, 44, 35–44. [Google Scholar] [CrossRef]
- Demeler, J.; Küttler, I.; Samson-Himmelstjerna, G. Adaptation and evaluation of three different in vitro tests for the detection of resistance to anthelmintics in gastro intestinal nematodes of cattle. Vet. Parasitol. 2010, 170, 61–70. [Google Scholar] [CrossRef]
- Costa-Junior, L.M.; Silva, C.R.; Soares, A.M.S.; Menezes, A.S.; Silva, M.R.L.; Amarante, A.F.T.; Costa, E.F.; Alencar, L.M.R. Assessment of biophysical properties of Haemonchus contortus from different life cycle stages with atomic force microscopy. Ultramicroscopy 2020, 209, 112862. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, J.T.A.; Souza, P.F.N.; Vasconcelos, I.M.; Dias, L.P.; Martins, T.F.; Van Tilburg, M.F.; Guedes, M.I.F.; Sousa, D.O.B. Mo-CBP3-PepI, Mo-CBP3-PepII, and Mo-CBP3-PepIII are synthetic antimicrobial peptides active against human pathogens by stimulating ROS generation and increasing plasma membrane permeability. Biochimie 2019, 157, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Gilleard, J.S.; Redman, E. Genetic Diversity and Population Structure of Haemonchus contortus. Adv. Parasitol. 2016, 93, 31–68. [Google Scholar] [CrossRef] [PubMed]
- Šimpraga, M.; Ljubičić, I.; Hlede, J.P.; Vugrovečki, A.S.; Marinculić, A.; Tkalčić, S. Alternative approaches for the control of gastrointestinal nematodes in sheep farming: A review. Berl. Munch. Tierarztl. Wochenschr. 2015, 128, 257–270. [Google Scholar]
- Alonso-Díazab, M.A.; Torres-Acostaa, J.F.J.; Sandoval-Castroa, C.A.; Capetillo-Leala, L.; Brunet, S.; Hostec, H. Effects of four tropical tanniniferous plant extracts on the inhibition of larval migration and the exsheathment process of Trichostrongylus colubriformis infective stage. Vet. Parasitol. 2008, 153, 187–192. [Google Scholar] [CrossRef]
- Hernández-Villegasa, M.M.; Borges-Argáeza, R.; Rodriguez-Vivasb, R.I.; Torres-Acostab, J.F.J.; Méndez-Gonzaleza, M.; Cáceres-Farfana, M. Ovicidal and larvicidal activity of the crude extracts from Phytolacca icosandra against Haemonchus contortus. Vet. Parasitol. 2011, 179, 100–106. [Google Scholar] [CrossRef]
- Jennings, C.; West, J.; Waine, C.; Craik, D.; Anderson, M. Biosynthesis and insecticidal properties of plant cyclotides: The cyclic knotted proteins from Oldenlandia affinis. Proc. Natl. Acad. Sci. USA 2001, 98, 10614–10619. [Google Scholar] [CrossRef] [Green Version]
- Malagón, D.; Botterill, B.; Gray, D.J.; Lovas, E.; Duke, M.; Gray, C.; Kopp, S.R.; Knott, L.M.; McManus, D.P.; Daly, N.L.; et al. Anthelminthic activity of the cyclotides (kalata B1 and B2) against schistosome parasites. Biopolymers 2013, 100, 461–470. [Google Scholar] [CrossRef]
- Holden-Dye, L.; Walker, R.J. Anthelmintic drugs and nematicides: Studies in Caenorhabditis elegans. Wormbook 2014, 1. [Google Scholar] [CrossRef]
- Wanderley, L.F.; Soares, A.M.; Silva, C.R.; Figueiredo, I.M.; Ferreira, A.T.; Perales, J.; Mota, H.R.; Oliveira, J.T.; Costa, L.M. A cysteine protease from the latex of Ficus benjamina has in vitro anthelmintic activity against Haemonchus contortus. Rev. Bras. Parasitol. Veterinária 2018, 27, 473–480. [Google Scholar] [CrossRef]
- Soares, A.M.S.; Oliveira, J.T.A.; Rocha, C.Q.; Ferreira, A.T.S.; Perales, J.; Zanatta, A.C.; Vilegas, W.; Silva, C.R.; Costa-Junior, L.M. Myracrodruon urundeuva seed exudates proteome and anthelmintic activity against Haemonchus contortus. PLoS ONE 2018, 13, e0200848. [Google Scholar] [CrossRef]
- Svangård, E.; Burman, R.; Gunasekera, S.; Lövborg, H.; Gullbo, J.; Göransson, U. Mechanism of action of cytotoxic cyclotides: Cycloviolacin O2 disrupts lipid membranes. J. Nat. Prod. 2007, 70, 643–647. [Google Scholar] [CrossRef]
- Butt, H.J.; Cappella, B.; Kappl, M. Force measurements with the atomic force microscope: Technique, interpretation and applications. Surf. Sci. Rep. 2005, 59, 1–152. [Google Scholar] [CrossRef] [Green Version]
- Henriques, S.T.; Huang, Y.-H.; Chaousis, H.; Wang, C.K.; Craik, C.J. Anticancer and toxic properties of cyclotides are dependent on phosphatidylethanolamine phospholipid targeting. Chembiochem 2014, 15, 1956–1965. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Properties | RcAlb-PepIII | Kalata B2 |
|---|---|---|
| Physicochemical | ||
| Sequence | AKLIPTIA | GLPVCGETCFGGTCNTPGCSCTWPICTRD |
| 1 Calculated molecular mass (Da) | 826.05 | 2979.81 |
| 2 Isoeletric point (pI) | 8.80 | 4.71 |
| 1 Hydrophobicity | 0.864 | 0.634 |
| 1 Net charge | +1 | −1 |
| 4 Ramachandran Plot (%) | 98 | 34 |
| 5 Tm | 0.56 | - |
| 5 sOPEP | −5.73 | - |
| Biological Properties | ||
| 6 CPP | No | Yes |
| 7 Allergic potential | No | Yes |
| Hemolytic potential | No | Yes |
| Toxic potential | No | Yes |
| 9 Cleavage sites | ||
| Trypsin (high pH) | 1 | 1 |
| Pepsin (pH 1.3) | 1 | 3 |
| Pepsin (pH > 2) | 1 | 4 |
| 10 Half-life time | 2.21 | 0.961 |
| 10 Stability | High | Normal |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soares, A.M.S.; Alencar, L.M.R.; Dias, L.P.; Lima, R.C.; Rocha Silva, C.; Santos-Oliveira, R.; Oliveira, J.T.A.; Junior, L.M.C.; Souza, P.F.N. New Insights into Anthelmintic Mechanisms of Action of a Synthetic Peptide: An Ultrastructural and Nanomechanical Approach. Polymers 2021, 13, 2370. https://doi.org/10.3390/polym13142370
Soares AMS, Alencar LMR, Dias LP, Lima RC, Rocha Silva C, Santos-Oliveira R, Oliveira JTA, Junior LMC, Souza PFN. New Insights into Anthelmintic Mechanisms of Action of a Synthetic Peptide: An Ultrastructural and Nanomechanical Approach. Polymers. 2021; 13(14):2370. https://doi.org/10.3390/polym13142370
Chicago/Turabian StyleSoares, Alexandra M. S., Luciana M. R. Alencar, Lucas P. Dias, Ruana C. Lima, Carolina Rocha Silva, Ralph Santos-Oliveira, Jose T. A. Oliveira, Livio M. C. Junior, and Pedro F. N. Souza. 2021. "New Insights into Anthelmintic Mechanisms of Action of a Synthetic Peptide: An Ultrastructural and Nanomechanical Approach" Polymers 13, no. 14: 2370. https://doi.org/10.3390/polym13142370