Aqueous Liquid-Liquid Phase Separation of Natural and Synthetic Polyguanidiniums

Abstract
:1. Introduction
2. Materials and Methods
2.1. Salmine Sulfate and Salmine Chloride
2.2. Coacervate Characterization
2.3. Phase Transition Temperature Determination by Turbidity
2.4. Isothermal Titration Calorimetry (ITC)
2.5. Synthesis of N-(3-methacrylamidopropyl)guanidinium Chloride
2.6. Synthesis and Characterization of Poly(3-guanidinopropyl methacrylamide-co-acrylamide) HCl
2.7. Preparation of Poly(3-guanidinopropyl methacrylamide-co-acrylamide) Sulfate
3. Results
3.1. Condensation of Salmine Sulfate
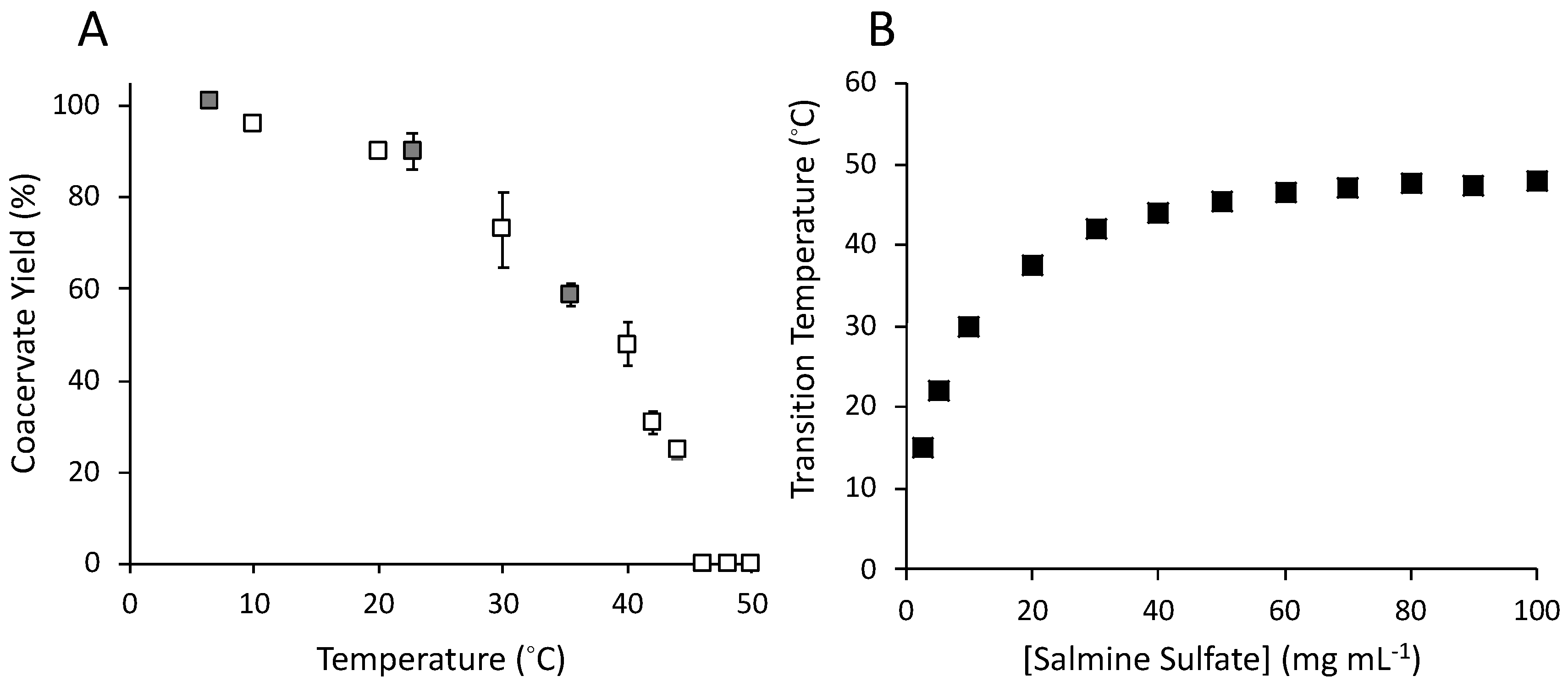
3.2. Temperature (T) and Concentration Dependence of Salmine Sulfate Condensation
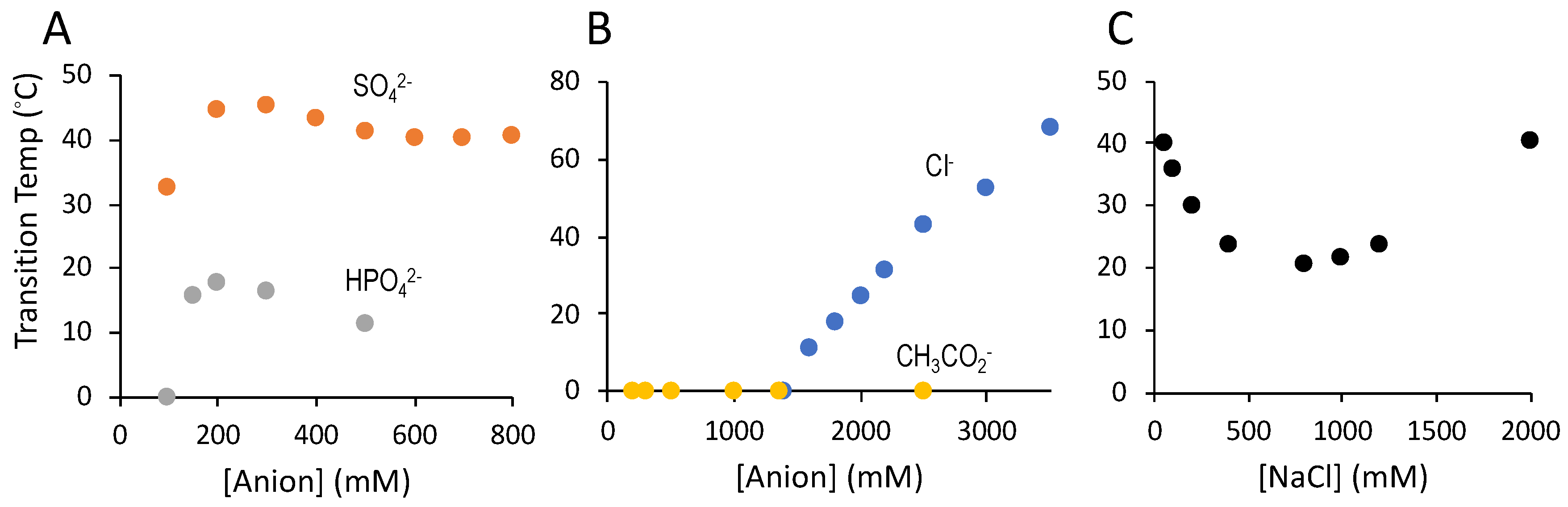
3.3. Counter-Anion Dependence of Salmine Condensation
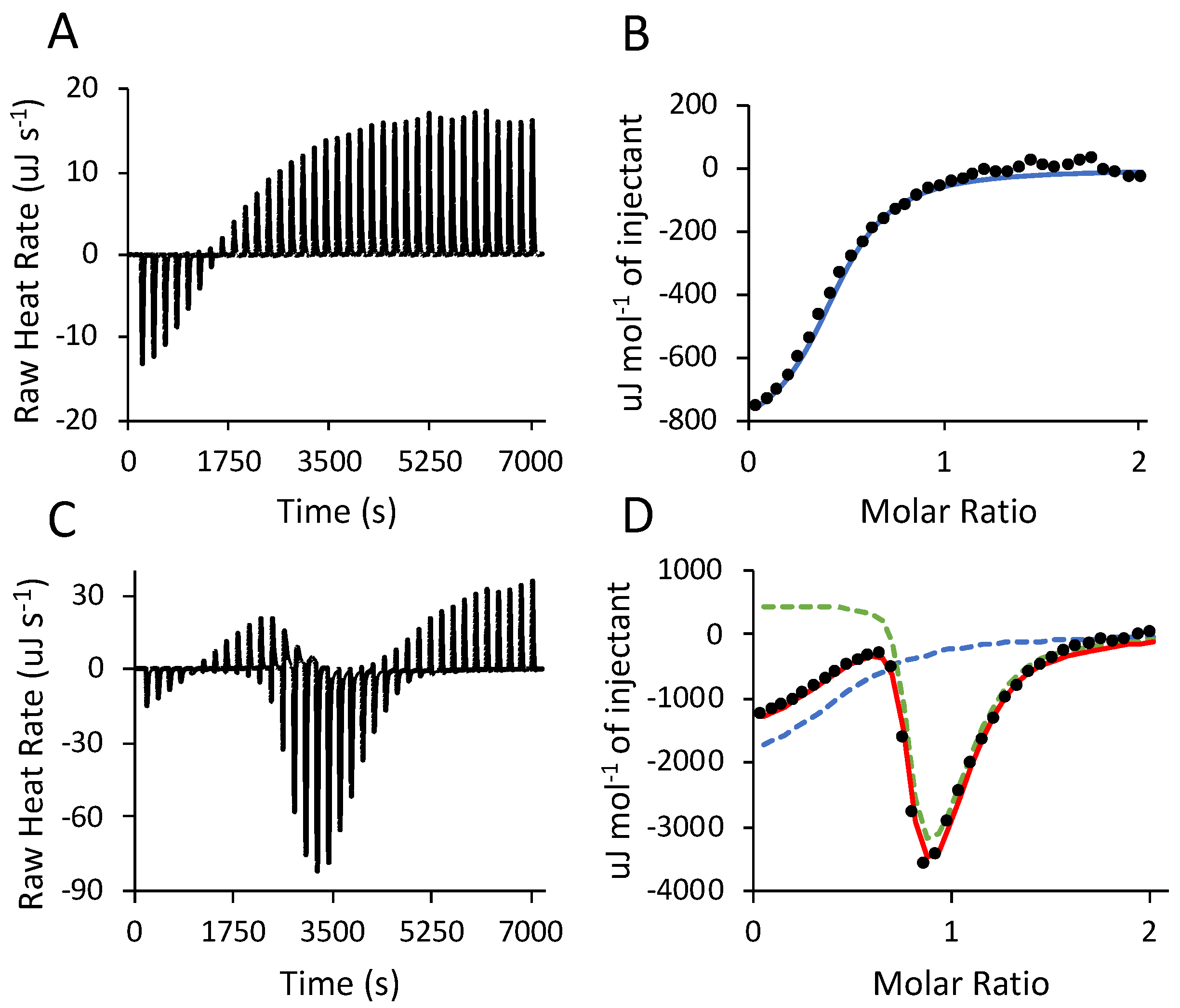
3.4. Isothermal Titration Calorimetry (ITC)
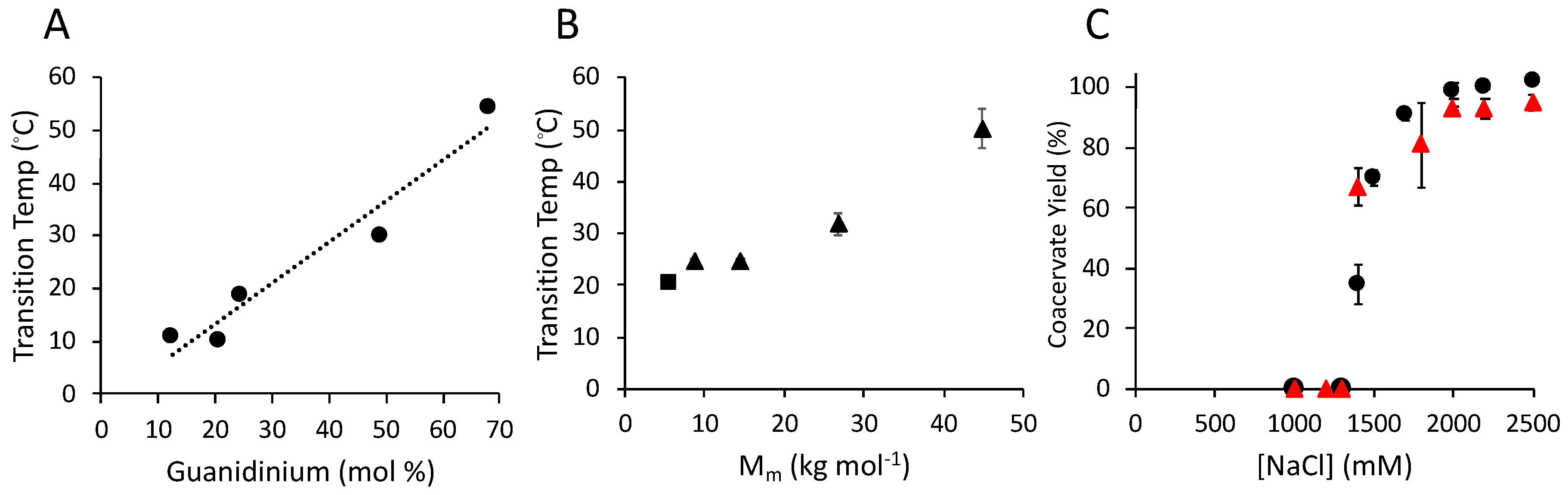
3.5. Condensation of Salmine Synthetic Analogs
4. Discussion
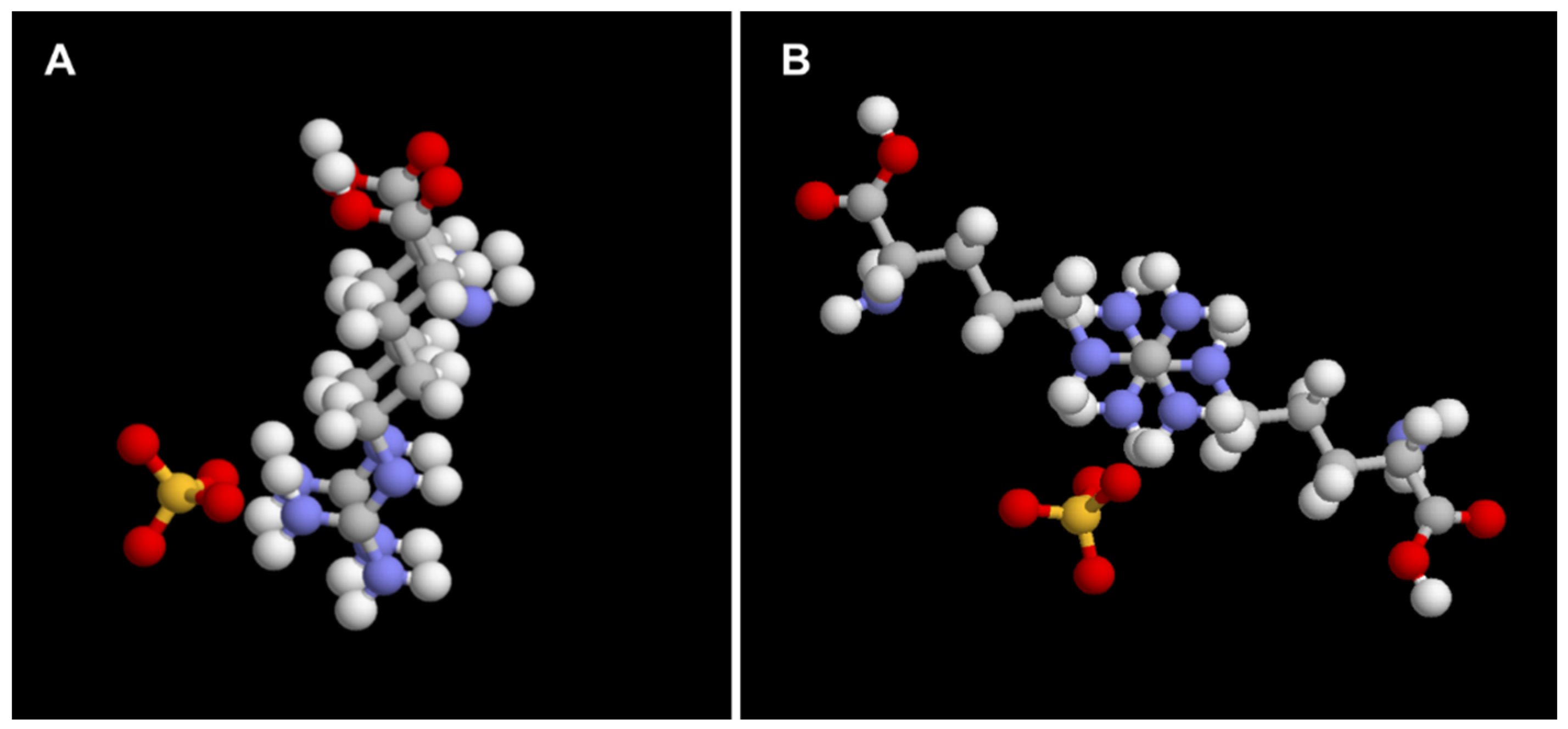
The Molecular Mechanism of Polyguanidinium LLPS
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Yevreinova, T.N. Concentration of Matter and Action of Enzymes in Coacervates; Nauka Press: Moscow, Russia, 1966. [Google Scholar]
- De Kruif, C.G.; Weinbreck, F.; De Vries, R. Complex Coacervation of Proteins and Anionic Polysaccharides. Curr. Opin. Colloid Interface Sci. 2004, 9, 340–349. [Google Scholar] [CrossRef]
- Brangwynne, C.P.; Tompa, P.; Pappu, R.V. Polymer physics of intracellular phase transitions. Nat. Phys. 2015, 11, 899–904. [Google Scholar] [CrossRef]
- Song, I.T.; Stewart, R.J. Complex coacervation of Mg(II) phospho-polymethacrylate, a synthetic analog of sandcastle worm adhesive phosphoproteins. Soft Matter 2018, 14, 379–386. [Google Scholar] [CrossRef]
- Hyman, A.A.; Weber, C.A.; Jülicher, F. Liquid-Liquid Phase Separation in Biology. Annu. Rev. Cell Dev. Biol. 2014, 30, 39–58. [Google Scholar] [CrossRef]
- Sear, R.P. Phase separation of equilibrium polymers of proteins in living cells. Faraday Discuss. 2008, 139, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Mitrea, D.M.; Kriwacki, R.W. Phase separation in biology; functional organization of a higher order. Cell Commun. Signal. 2016, 14, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.; Brangwynne, C.P. Liquid phase condensation in cell physiology and disease. Science 2017, 357, eaaf4382. [Google Scholar] [CrossRef] [PubMed]
- Aguzzi, A.; Altmeyer, M. Phase Separation: Linking Cellular Compartmentalization to Disease. Trends Cell Biol. 2016, 26, 547–558. [Google Scholar] [CrossRef]
- Elbaum-Garfinkle, S.; Kim, Y.; Szczepaniak, K.; Chen, C.C.-H.; Eckmann, C.R.; Myong, S.; Brangwynne, C.P. The disordered P granule protein LAF-1 drives phase separation into droplets with tunable viscosity and dynamics. Proc. Natl. Acad. Sci. USA 2015, 112, 7189–7194. [Google Scholar] [CrossRef] [PubMed]
- Nott, T.J.; Petsalaki, E.; Farber, P.; Jervis, D.; Fussner, E.; Plochowietz, A.; Craggs, T.D.; Bazett-Jones, D.P.; Pawson, T.; Forman-Kay, J.D.; et al. Phase Transition of a Disordered Nuage Protein Generates Environmentally Responsive Membraneless Organelles. Mol. Cell 2015, 57, 936–947. [Google Scholar] [CrossRef]
- Qamar, S.; Wang, G.; Randle, S.J.; Ruggeri, F.S.; Varela, J.A.; Lin, J.Q.; Phillips, E.C.; Miyashita, A.; Williams, D.; Ströhl, F.; et al. FUS Phase Separation Is Modulated by a Molecular Chaperone and Methylation of Arginine Cation-π Interactions. Cell 2018, 173, 720–725. [Google Scholar] [CrossRef]
- Varadi, M.; Zsolyomi, F.; Guharoy, M.; Tompa, P. Functional Advantages of Conserved Intrinsic Disorder in RNA-Binding Proteins. PLoS ONE 2015, 10, e0139731. [Google Scholar] [CrossRef] [PubMed]
- Burke, K.A.; Janke, A.M.; Rhine, C.L.; Fawzi, N.L. Residue-by-Residue View of In Vitro FUS Granules that Bind the C-Terminal Domain of RNA Polymerase II. Mol. Cell 2015, 60, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Rhoads, S.; Monahan, Z.; Yee, D.; Shewmaker, F. The Role of Post-Translational Modifications on Prion-Like Aggregation and Liquid-Phase Separation of FUS. Int. J. Mol. Sci. 2018, 19, 886. [Google Scholar] [CrossRef] [PubMed]
- Hofweber, M.; Hutten, S.; Bourgeois, B.; Spreitzer, E.; Niedner-Boblenz, A.; Schifferer, M.; Ruepp, M.D.; Simons, M.; Niessing, D.; Madl, T.; et al. Phase Separation of FUS Is Suppressed by Its Nuclear Import Receptor and Arginine Methylation. Cell 2018, 173, 706–719. [Google Scholar] [CrossRef]
- DeJesus-Hernandez, M.; Mackenzie, I.R.; Boeve, B.F.; Boxer, A.L.; Baker, M.; Rutherford, N.J.; Nicholson, A.M.; Finch, N.A.; Flynn, H.; Adamson, J.; et al. Expanded GGGGCC Hexanucleotide Repeat in Noncoding Region of C9ORF72 Causes Chromosome 9p-Linked FTD and ALS. Neuron 2011, 72, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Kwon, I.; Xiang, S.; Kato, M.; Wu, L.; Theodoropoulos, P.; Wang, T.; Kim, J.; Yun, J.; Xie, Y.; McKnight, S.L. Poly-dipeptides encoded by the C9orf72 repeats bind nucleoli, impede RNA biogenesis, and kill cells. Science 2014, 345, 1139–1145. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.H.; Zhang, P.; Kim, H.J.; Mitrea, D.M.; Sarkar, M.; Freibaum, B.D.; Cika, J.; Coughlin, M.; Messing, J.; Molliex, A.; et al. C9orf72 Dipeptide Repeats Impair the Assembly, Dynamics, and Function of Membrane-Less Organelles. Cell 2016, 167, 774–788. [Google Scholar] [CrossRef]
- Balhorn, R. The protamine family of sperm nuclear proteins. Genome Biol. 2007, 8, 227. [Google Scholar] [CrossRef]
- Kossel, A. The Protamines and Histones; Longmans Green and Co.: London, UK, 1928. [Google Scholar]
- Sautière, P.; Briand, G.; Gusse, M.; Chevaillier, P. Primary structure of the protamine isolated from the sperm nuclei of the dog-fish Scylliorhinus caniculus. Eur. J. Biochem. 1981, 119, 251–255. [Google Scholar] [CrossRef]
- Watanabe, R.; Suzuki, T.; Oshima, Y. Development of quantitative NMR method with internal standard for the standard solutions of paralytic shellfish toxins and characterisation of gonyautoxin-5 and gonyautoxin-6. Toxicon 2010, 56, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Freire, E.; Mayorga, O.L.; Straume, M. Isothermal Titration. Anal. Chem. 1990, 62, 950–959. [Google Scholar] [CrossRef]
- Bernatowicz, M.S.; Wu, Y.; Matsueda, G.R. 1H-Pyrazole-1-carboxamidine hydrochloride an attractive reagent for guanylation of amines and its application to peptide synthesis. J. Org. Chem. 1992, 57, 2497–2504. [Google Scholar] [CrossRef]
- Yonamine, Y.; Yoshimatsu, K.; Lee, S.H.; Hoshino, Y.; Okahata, Y.; Shea, K.J. Polymer Nanoparticle−Protein Interface. Evaluation of the Contribution of Positively Charged Functional Groups to Protein Affinity. ACS Appl. Mater. Interfaces 2013, 5, 374–379. [Google Scholar] [CrossRef]
- Kayitmazer, A.B. Thermodynamics of complex coacervation. Adv. Colloid Interface Sci. 2017, 239, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Aberkane, L.; Jasniewski, J.; Gaiani, C.; Scher, J.; Sanchez, C. Thermodynamic Characterization of Acacia Gum-β-Lactoglobulin Complex Coacervation. Langmuir 2010, 26, 12523–12533. [Google Scholar] [CrossRef] [PubMed]
- Neves, M.A.C.; Yeager, M.; Abagyan, R. Unusual Arginine Formations in Protein Function and Assembly: Rings, Strings, and Stacks. J. Phys. Chem. B 2012, 116, 7006–7013. [Google Scholar] [CrossRef]
- Spruijt, E. Strength, Structure and Stability of Polyelectrolyte Complex Coacervates. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 2012. [Google Scholar]
- Wang, Q.; Schlenoff, J.B. The Polyelectrolyte Complex/Coacervate Continuum. Macromolecules 2014, 47, 3108–3116. [Google Scholar] [CrossRef]
- Magalhaes, A.; Maigret, B.; Hoflack, J.; Gomes, J.N.; Scheraga, H.A. Contribution of unusual arginine-arginine short-range interactions to stabilization and recognition in proteins. J. Protein Chem. 1994, 13, 195–215. [Google Scholar] [CrossRef]
- Vondrášek, J.; Mason, P.E.; Heyda, J.; Collins, K.D.; Jungwirth, P. The Molecular Origin of Like-Charge Arginine−Arginine Pairing in Water. J. Phys. Chem. B 2009, 113, 9041–9045. [Google Scholar] [CrossRef]
- No, K.T.; Nam, K.-Y.; Scheraga, H.A. Stability of Like and Oppositely Charged Organic Ion Pairs in Aqueous Solution. J. Am. Chem. Soc. 1997, 119, 12917–12922. [Google Scholar] [CrossRef]
- Vazdar, M.; Uhlig, F.; Jungwirth, P. Like-Charge Ion Pairing in Water: An Ab Initio Molecular Dynamics Study of Aqueous Guanidinium Cations. J. Phys. Chem. Lett. 2012, 3, 2021–2024. [Google Scholar] [CrossRef]
- Mason, P.E.; Neilson, G.W.; Enderby, J.E.; Saboungi, M.-L.; Dempsey, C.E.; MacKerell, A.D.; Brady, J.W. The Structure of Aqueous Guanidinium Chloride Solutions. J. Am. Chem. Soc. 2004, 126, 11462–11470. [Google Scholar] [CrossRef] [PubMed]
- Boudon, S.; Wipff, G.; Maigret, B. Monte Carlo Simulations on the Like-Charged Guanidinium-Guanidinium Ion Pair in Water. J. Phys. Chem. 1990, 94, 6056–6061. [Google Scholar] [CrossRef]
- Mason, P.E.; Neilson, G.W.; Dempsey, C.E.; Brady, J.W. Nanometer-Scale Ion Aggregates in Aqueous Electrolyte Solutions: Guanidinium Sulfate and Guanidinium Thiocyanate. J. Phys. Chem. B 2005, 109, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.P.; Shukla, D.; Trout, B.L. Arginine and the Hofmeister Series: The Role of Ion–Ion Interactions in Protein Aggregation Suppression. J. Phys. Chem. B 2011, 115, 7447–7458. [Google Scholar] [CrossRef] [PubMed]
- Gund, P. Guanidine, trimethylenemethane, and” Y-delocalization.” Can acyclic compounds have “aromatic” stability? J. Chem. Educ. 1972, 49, 100–103. [Google Scholar] [CrossRef]
- Gobbi, A.; Frenking, G. Y-Conjugated Compounds: The Equilibrium Geometries and Electronic Structures of Guanidine, Guanidinium Cation, Urea, and 1,l-Diaminoethylene. J. Am. Chem. Soc. 1993, 115, 2362–2372. [Google Scholar] [CrossRef]
- Mason, P.E.; Neilson, G.W.; Dempsey, C.E.; Barnes, A.C.; Cruickshank, J.M. The hydration structure of guanidinium and thiocyanate ions: Implications for protein stability in aqueous solution. Proc. Natl. Acad. Sci. USA 2003, 100, 4557–4561. [Google Scholar] [CrossRef]
- Hunger, J.; Niedermayer, S.; Buchner, R.; Hefter, G. Are Nanoscale Ion Aggregates Present in Aqueous Solutions of Guanidinium Salts? J. Phys. Chem. B 2010, 114, 13617–13627. [Google Scholar] [CrossRef] [PubMed]
- Vazdar, M.; Heyda, J.; Mason, P.E.; Tesei, G.; Allolio, C.; Lund, M.; Jungwirth, P. Arginine “Magic”: Guanidinium Like-Charge Ion Pairing from Aqueous Salts to Cell Penetrating Peptides. Acc. Chem. Res. 2018, 51, 1455–1464. [Google Scholar] [CrossRef] [PubMed]
- Boeynaems, S.; Bogaert, E.; Kovacs, D.; Konijnenberg, A.; Timmerman, E.; Volkov, A.; Guharoy, M.; De Decker, M.; Jaspers, T.; Ryan, V.H.; et al. Phase Separation of C9orf72 Dipeptide Repeats Perturbs Stress Granule Dynamics. Mol. Cell 2017, 65, 1044–1055. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total [anion] (mM) | Salmine (mg mL−1) | Water (wt%) | Molar Ratio SO42− to Gdm+ |
|---|---|---|---|
| Salmine Sulfate | |||
| 99 | 617 ± 33 | 49.8 ± 2.2 | 0.43 ± 0.03 |
| 200 | 634 ± 68 | 50.0 ± 4.0 | 0.44 ± 0.09 |
| 400 | 602 ± 11 | 47.8 ± 0.5 | 0.47 ± 0.00 |
| 800 | 643 ± 46 | 47.0 ± 1.5 | 0.47 ± 0.01 |
| Salmine Chloride | |||
| 1400 | 465 ± 58 | 66.8 ± 4.3 | -- |
| 1500 | 511 ± 11 | 60.9 ± 2.7 | -- |
| 1700 | 553 ± 27 | 56.0 ± 0.8 | -- |
| 2000 | 616 ± 17 | 55.3 ± 2.7 | -- |
| 2200 | 626 ± 6 | 52.2 ± 1.4 | -- |
| 2500 | 637 ± 6 | 48.5 ± 2.4 | -- |
| Temperature (°K) | N | Ka (M−1) | H (kJ mol−1) | −TΔS (kJ mol−1) | G (kJ mol−1) |
|---|---|---|---|---|---|
| 298.15 * | 0.45 | 3.96 × 102 | −2.18 | −12.65 | −14.83 |
| 283.15 | 0.49 | 2.09 × 102 | −5.45 | −7.13 | −12.58 |
| 283.15 | 0.76 | 1.15 × 105 | 1.15 | −28.60 | −27.45 |
| 283.15 | 0.33 | 4.90 × 102 | −11.71 | −2.87 | −14.58 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prather, L.J.; Weerasekare, G.M.; Sima, M.; Quinn, C.; Stewart, R.J. Aqueous Liquid-Liquid Phase Separation of Natural and Synthetic Polyguanidiniums. Polymers 2019, 11, 649. https://doi.org/10.3390/polym11040649
Prather LJ, Weerasekare GM, Sima M, Quinn C, Stewart RJ. Aqueous Liquid-Liquid Phase Separation of Natural and Synthetic Polyguanidiniums. Polymers. 2019; 11(4):649. https://doi.org/10.3390/polym11040649
Chicago/Turabian StylePrather, Leland J., G. Mahika Weerasekare, Monika Sima, Colette Quinn, and Russell J. Stewart. 2019. "Aqueous Liquid-Liquid Phase Separation of Natural and Synthetic Polyguanidiniums" Polymers 11, no. 4: 649. https://doi.org/10.3390/polym11040649