3.1. Shape of Bulk COM Crystals Formed in the Supersaturated Pure Solution and Solution Containing L-Aspartic Acid 6mer (L-Asp6)
The COM crystal has a monoclinic crystal system with unit cell parameters of a = 9.976 Å, b = 14.588 Å, c = 6.291 Å, and β = 107.05° [
26]. It typically forms with a hexagonal crystal morphology in the pure supersaturated solution, although the shape can be somewhat different depending on the synthesis conditions, such as ionic strength, pH, temperature, etc. [
16,
24].
Figure 1a shows a COM crystal with a hexagonal crystal morphology, synthesized in a pure calcium oxalate solution (0.7 mM CaCl
2, 0.7 mM K
2C
2O
4, 46.6 mM KCl), with its broadest face, (
01) placed on the carbon tape of the substrate.
As seen, the COM crystal is the twin crystal (indicated by Crystal 1 and Crystal 2, and also another, Crystal 3, is seen), and all or nearly all of the observed crystals had this characteristic in this synthesis condition. As already reported in previous papers, the broadest COM face (
01) is known to have many growth hillocks, as shown in
Figure 1c, resembling the triangle morphology during their growth. These hillocks consist of many spiral turns with molecular steps (indicated by a dashed white arrow in
Figure 1c) 0.57 nm high [
16,
17], of which generation from the screw dislocation outcrop is the main topic of this paper. The shape of the growth hillocks and location on the (
01) where they were frequently found during in situ liquid-phase AFM observation is indicated by placing a schematic (called Hillock A) on that location which is the left side of the crystal shown in
Figure 1a. In this orientation of the placement of the crystal, the hillocks (called Hillock B), which had mirror-symmetrical relationships, were frequently observed in the places close to the right side of the whole crystal (or Crystal 2) seen by a schematic placed on that location due to the twin structure of the bulk crystal.
When the synthesis solution included 20 μM L-aspartic acid 6mer (Asp
6)—a model peptide—which mimics the acidic functional group (COOH) of molecules regulating the COM formation, the synthesized crystals had such morphologies as one seen in
Figure 1b. This morphology is similar to those synthesized in the solution containing citrate with carboxylic moieties, a regulating peptide for the COM crystal formation [
13]. A comparison of the crystal morphologies formed from the pure solution and solution containing 20 μM L-Asp
6 shows that growth along the direction of length (L) was inhibited relatively more to the direction of width (W), given by the value of W/L = 0.38 for the pure solution (
Figure 1a) versus W/L = 0.55 for the solution containing 20 μM L-Asp
6 (
Figure 1b).
As seen in the results of in situ liquid-phase AFM investigation of the effect of citrate on the growth of the hillocks, which explained the shape modification of COM crystals [
11,
13], the AFM observation of the effect of L-Asp
6 on the growth of the hillocks can account for the morphological modification seen in
Figure 1b. This will be dealt with later while the effect of L-Asp
6 on the step generation from the screw dislocation outcrop on the (
01) surface is explained.
3.2. Spiral Generation from the Screw Dislocation Outcrop at the (01) Crystal Surface in the Supersaturated Pure Solution
The COM crystal grows by the growth of molecular steps generated from the screw dislocation outcrop [
27] at the crystal surface in the solution at low to moderate supersaturations like many other crystals, such as calcite [
28,
29,
30] and brushite [
31]. The inset of
Figure 2b shows an AFM tip scanning on the growing (
01) surface of a COM crystal grown by the gel method [
24]—much larger than that in
Figure 1a—in a supersaturated pure calcium/oxalate solution. During this in situ observation of the growing (
01) surface, molecular steps generated from the aforementioned screw dislocation outcrops were seen to grow and form the growth hillock such as one shown in
Figure 1c [
16,
17,
19,
20].
Consecutive in situ liquid-phase AFM images (
Figure 2) capturing the growth events occurring in a location as an area within a dashed white box in
Figure 1c show how a molecular step spiral at the screw dislocation outcrop on the (
01) surface was generated and subsequently how the growth hillock could form. In
Figure 2a, the location of a screw dislocation outcrop is indicated by a dashed white arrow and is seen in more detail in
Figure 2e, which is a magnified view of a white box in
Figure 2a. In
Figure 2e, not yet a new spiral did form and propagate because a step segment (named
) with a length of L, ~26 nm, which would propagate along [
0
] direction at exceeding a critical L
c did not reach a critical L
c; the step segment starts to advance when its length L exceeds L
c because, then, the volume-free energy release accompanied by the step movement, i.e., solid formation begins to be larger than the step edge energy created by the creation of the step edges at both ends of the step line. See ref. [
29] for a more detailed explanation of this.
In
Figure 2b, still, a new spiral did not form and propagate, meaning a step segment,
did not reach L
c. However, as seen in
Figure 2f, which is a magnified view of a white box in
Figure 2b, steps grew along [1
0] and [120] a little bit, as shown by a comparison of them with those in
Figure 2e; the [1
0] and [120] steps in
Figure 2f are shown to have moved to the left a little compared to those in
Figure 2e due to their growth. At this time,
was accordingly also slightly lengthened by the growth of those steps along [1
0] and [120];
= ~29 nm in
Figure 2f versus ~26 nm in
Figure 2e. Then, in an image corrected in 18 s, as seen in
Figure 2c, a new spiral was seen to have been generated and grown. This suggests that the step segment with
in
Figure 2f almost reached L
c. (Note that the captured morphology of the [
0
] step line during the upward scanning in
Figure 2c is slanted clockwise, although the true orientation of the [
0
] step line is (almost) vertical, as represented by a schematic located in the left corner of the bottom of the figure. The so-called step orientation distortion in the in situ AFM images recording moving steps occurs with its magnitude proportional to the step-growth speed for a given scan rate because the whole parts of the steps are imaged serially from top to bottom (down scan) or from bottom to top (up scan) at a finite scan rate, rather than imaged at once for AFM [
19,
32] (especially see Supplementary Material of ref. [
19] for detailed explanation)). In
Figure 2d, the spiral seen in
Figure 2c had grown further, and the [
0
] step line was slanted counter-clockwise because it was captured by the down scan.
As is known now, the process seen in
Figure 2a–d is repeated (a generation or one full rotation time (t
r) of this spiral shown in the figure was 69 ± 2 s), and this process leads to the formation of the growth hillock seen in
Figure 1c.
The results of the generation of the molecular step spiral are schematically summarized in
Figure 2g. At Stage 1, a step segment has a length,
smaller than L
c. Then, as the [120] and [1
0] steps grow, the step segment accordingly lengthens and becomes to have
reach L
c (Stage 2). From this time, as seen in Stage 3, the segment starts to advance toward the [
0
] direction, and in the meantime, other steps move along their respective directions to be a new spiral. Although little expressed, [
20] and [
0] steps are observed in the spiral [
19]. While the new spiral grows, the step segment S connected to the emergence point of the screw dislocation at the crystal surface is turning around the point; thus, its orientation varies while the spiral grows, as is seen by looking at the orientation of segment S at Stage 4 in comparison to it at Stage 3. As the spiral grows further, the turning step segment S becomes one with
reaching L
c (Stage 5), completing one full rotation. Then, it starts to advance, and in the meantime, the already-formed spiral continues to grow (Stage 6). Again, although little expressed, the components of the mirror-symmetry related [
20]/[
0] steps and [010]/[0
0] steps are observed in the spiral (at Stages 5 and 6).
As now understood by the schematic (
Figure 2g), one full spiral rotation time, t
r, is influenced by the growth rate (V
S) of step segments around the emergence point of the screw dislocation, or V of large steps (i.e., L >> L
c), with a relationship of t
r 1/V, because V
S is proportional to V [
29]. Thus, one full spiral rotation time is affected by supersaturation (σ) and temperature, which determines V, like the bulk nucleation process [
33]. As previously reported in the calcite system [
29], L
c is inversely proportional to σ, and V is proportional to σ at a given temperature, and consequently, this relationship of t
r 1/V
1/σ exists.
3.3. Effect of L-Asp6 on the Birth and Growth of the Step Spiral on the (01) Face
Citrate [
11,
13,
14,
18,
22] and aspartic acid 6mer (L-Asp
6) [
17,
19] with the functional group of carboxylic acids had the strongest inhibitory effect on the growth of the molecular step spiral on the broadest (
01) face of the COM crystal, as shown in
Figure 1a. For example, citrate changed the step spiral resembling a triangle shape into a round disk-shaped one by adsorbing to and roughening the growing steps of the spiral, especially with dropping the fastest growing [
0
] step speed by a factor of 25 at a supersaturation (σ) of 0.7, which led to similar growth speed over all the growth directions of the step spiral. Although those studies well demonstrated the effect of those molecules on the spiral growth in terms of morphological and step speed changes, that is, the inhibition of the step-growth, they lacked in demonstrating in detail how they affect the birth of a new spiral and its subsequent temporal growth compared to those occurring in the pure solution.
Figure 3 shows how the generation of a new step spiral at the dislocation outcrop on the (
01) face and its subsequent temporal growth were affected when the step spirals growing in the pure solution were exposed to the solution containing 2 µM L-Asp
6 at σ
0.93.
Figure 3a shows a triangle-resembling topmost molecular step spiral with its [
0
] step growing at a growth rate of 27 nm/s in the pure calcium/oxalate solution at σ = 0.93. (As already explained in dealing with
Figure 2, the true orientation of the [
0
] step line is (almost) vertical as expressed in the schematic of
Figure 2c, although it appears slanted counter-clockwise.)
Figure 3b shows the morphology of the step spirals shown in
Figure 3a when they had grown in the supersaturated calcium/oxalate solution (σ
0.93) containing 2 µM L-Asp
6 for about 4 min and 30 s; this image of
Figure 3b was collected by the down scan with an acquisition time of 18 s; thus, the top and bottom of the image are arbitrarily set to 0 s and 18 s, and subsequent images (
Figure 3c–e) have times marked accordingly.
Compared to the step spirals growing in the pure solution at σ = 0.93, the [120] and [1
0] step lines became rough, as seen simply by a comparison of the step lines within yellow boxes in
Figure 3b with those in
Figure 3a, and cusps in the [
0
] step line developed. This suggests that L-Asp
6 was bound to the step lines. At this moment, the [
0
] step speed was dramatically decreased to ~0.8 nm/s, indicating that the additive adsorbed to the step line exerted a great effect on inhibiting the step growth. Also, note that unlike the fast-growing [
0
] step appearing slanted, that is, appearing with the step line orientation distorted in the pure supersaturated solution shown in
Figure 3a, the step orientations along all the directions in
Figure 3b represent almost true ones because the speed of steps was so slow with ~0.8 nm that the orientation distortion in the captured moving steps was minimal.
To help explain the effect of L-Asp
6 on the birth of a new spiral and temporal growth of the topmost and second topmost spirals, their morphologies are shown in
Figure 3b is drawn schematically in
Figure 4b, together with those in the pure solution without L-Asp
6 such as one expressed in
Figure 4a.
Although the cusps developed due to the adsorption of L-Asp
6 to the step lines of the second topmost spiral, as seen in
Figure 4b, the overall morphology of this second topmost spiral is still similar to the triangle-resembling morphology of the spiral from the pure solution seen in
Figure 4a at this moment. This happens because the second topmost spiral resembling triangle morphology was first generated from the pure solution before the introduction of 2 µM L-Asp
6-containing solution and got the adsorption of L-Asp
6 on the way of its propagation (see
Materials and Methods for details on the AFM imaging).
Unlike this morphology of the second topmost spiral, a newly-born topmost spiral does not have a pseudo-triangle shape, which is the morphology achieved when a spiral newly forms in the pure supersaturated solution; rather, it has a morphology similar to the hexagonal bulk crystal. This indicates that the adsorption of L-Asp
6 to step segments just emanating from the dislocation outcrop has a dramatic effect on the step growth orientation and, subsequently, the spiral morphology from the time of generation of a new spiral. Since the birth of new step segments with the critical step lengths and the orientation of the segments at the dislocation outcrop are controlled by the step edge free energy rather than the kinetic factor, the adsorption of L-Asp
6 to the step segments emanating from the dislocation outcrop is considered to have induced new equilibrium step directions like the case of step spirals of calcite emanating from the dislocation outcrop with adsorption of aspartic acid to them [
29].
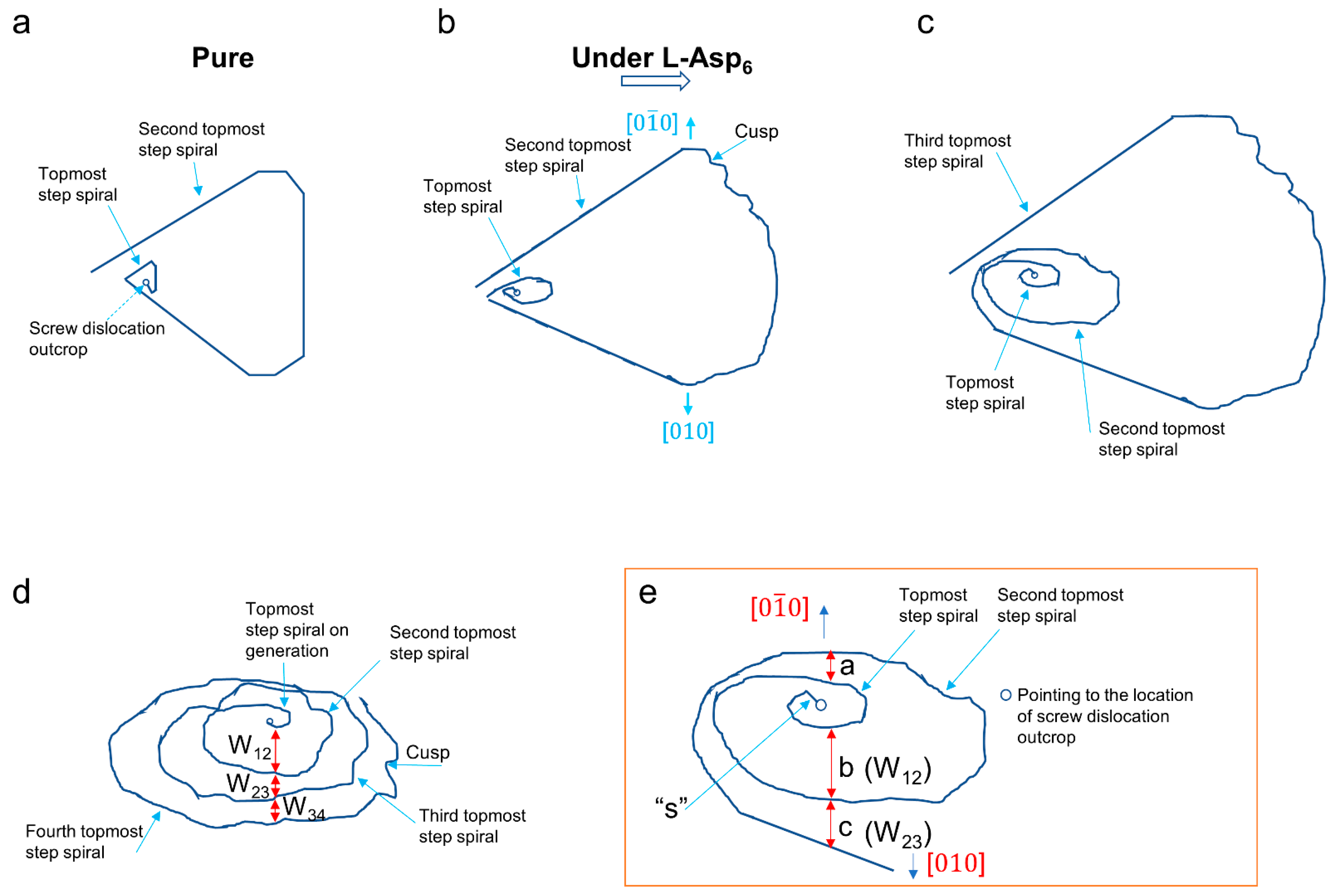
Figure 3c shows the morphological evolution of molecular steps at the dislocation hillock shown in
Figure 3b, captured 367 s later. Again, for an easier explanation of the effect of adsorbed L-Asp
6 on the spiral generation and growth for this time interval, the morphologies of the first to third topmost spirals shown in
Figure 3c are expressed schematically in
Figure 4c, and the morphologies of those first and second topmost spirals in
Figure 4c are magnified in
Figure 4e. As seen in
Figure 4e, the step width (W
12) along [0
0] direction between the topmost and second topmost spiral (i.e., the distance of “a”: 62 nm for
Figure 3c) is smaller than W
12 along [010] direction (i.e., the distance of “b”: 112 nm for
Figure 3c). Since the step speed (V) is proportional to the distance of the movement for a given time, the [0
0] step had about twice smaller V than the [010] step, indicating L-Asp
6 had about twice the stronger inhibition effect on the growth of the [0
0] over [010] step. This kind of characteristic of chiral aspartic acid 6mers inhibiting one of two symmetrical steps of the COM step spiral preferentially and of aspartic acid doing on calcite, which induced chiral morphology of the step spirals was already reported [
16,
20,
34].
Now let us see the characteristic of the step generation under the adsorbed L-Asp
6 further beyond the morphological changes from the pseudo triangle to hexagon-like morphology, compared to the case from the pure solution. For the pure solution (see
Figure 3a), once a new step spiral resembling a triangle was generated from the dislocation outcrop in the dislocation hillock, as reported recently by Cho et al. [
20], the symmetrical [120] and [1
0] steps (<120> steps) of the topmost spiral, such as one indicated by a yellow arrow, which had a wider step width (with respect to the second topmost [120] step) than those already formed <120> steps such as the second topmost <120> step and <120> steps indicated by a circle in the periphery of the hillock, for a brief moment, moved faster than them. As a result, the <120> steps of the newly born topmost spiral caught up with the second topmost <120> step and then moved at the same speed, keeping the same or similar step widths, all together with the <120> steps indicated by a white circle in the periphery afterward; the [120] step of the topmost spiral with the step width (W
12) between the topmost and second topmost [120] step of 82.1 ± 6.0 nm grew at 6.7 ± 0.8 nm/s and caught up with the [120] step of the second topmost spiral with a step width (W
23) of 38.9 ± 6.0 nm respect to the [120] step of the third topmost spiral (see ref. [
20] for details). The strong dependence of V of the topmost <120> step on W until it reached those in the periphery happened because calcium and oxalate ions over the terrace between the topmost and second topmost <120> step were the main supply source, which had a much smaller ion concentration than bulk solution volume facing the terrace, for the growth of the newly born topmost <120> step; in other words, the step grew by the so-called surface diffusion growth mechanism [
20,
28].
When the topmost spiral in
Figure 3b grew for about 367 s, it became the second topmost spiral shown in
Figure 3c, and a new spiral, i.e., the topmost spiral, just formed. The spiral generation was greatly delayed with a generation time (t
r) of about 7 min at this moment, compared to t
r of 69 ± 2 s in the pure solution; later, when exposed to the solution for a longer time, t
r was around 13 min. This is natural because a new spiral is generated once a step segment (marked as “S” in
Figure 4e) originating from the point of the screw dislocation outcrop at the crystal surface makes one complete turn around the point, and the speed of this movement is in the similar order of the step-growth rate of the topmost spiral in a direction such as [
0
], 0.4 nm/s, given that the ellipse-like hexagonal morphology of the topmost spiral has small anisotropy in the dependence of the growth rate on the step propagation directions. At this moment, b, i.e., W
12 along [010], is larger than c, i.e., W
23 in
Figure 4e.
Figure 3d shows the morphology of the spirals shown in
Figure 3c after 36 s. Due to so slow growth rate of the spirals, little morphological changes happened during this interval. Then, after relatively sufficient time passed, several spirals were seen to have developed from the origin of the dislocation outcrop, as seen in
Figure 3e, which was collected 659 s after
Figure 3d. Several spirals from the screw dislocation outcrop shown in
Figure 3e are magnified in
Figure 3f. Like before, a schematic (
Figure 4d) of the spirals is drawn to explain the character of the step generation under the adsorbed L-Asp
6 at this stage effectively. As seen,
Figure 4d expresses four spiral turns of
Figure 3e from the topmost to the fourth topmost spirals. The step width (W
12) between the topmost and second topmost spirals is larger than the subsequent step widths, W
23 and W
34. That W
12 is larger than the subsequent ones is a phenomenon that is also seen in
Figure 3c (or
Figure 4e). Given that the current topmost and second topmost spirals in
Figure 4d become the third and fourth topmost spirals, respectively, when new topmost and second topmost spirals form, this suggests that the step width (W
12) decreases while the topmost and second topmost spirals grow. This is achieved because the topmost spiral grows faster than the second topmost spiral when W
12 is larger than W
23 and then moves at the same or similar speed as the second topmost spiral after reaching the same or similar step width as W
23. Thus, this indicates that the steps of the spirals grow predominantly by incorporating calcium and oxalate ions diffusing over the terrace, with the same growth mechanism via surface diffusion-controlled growth as in the pure supersaturated solution [
20], while their growth is highly inhibited by L-Asp
6 adsorbed to the terraces and steps.
The ellipse-like hexagonal morphology of the step spirals developed under L-Asp
6 in
Figure 3e (or
Figure 3f) parallels the bulk COM crystal morphology produced in the supersaturated solution containing L-Asp
6 shown in
Figure 1b. This result indicates that the modified morphology of the molecular step spirals on the broadest COM crystal face (
01) is mainly transferred to the bulk crystal morphology, as in the case of COM crystal growth in the solution containing citrate [
11,
13]. It is natural because the COM crystal grows by the growth of molecular spirals on its faces, and the morphological modification of the broadest face plays the most important role in shaping the bulk crystal morphology.


{kind=link}
{kind=link}
{kind=link}
{kind=link}



