Catalytic Activities of Multimeric G-Quadruplex DNAzymes

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
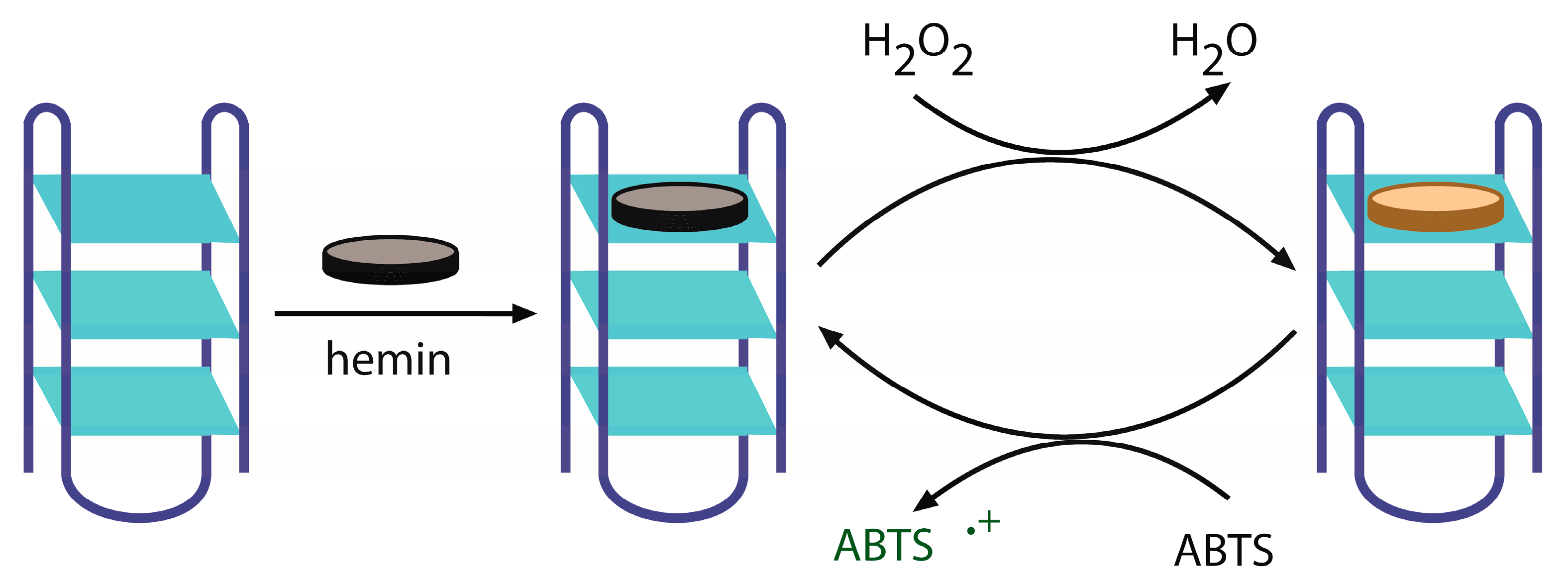
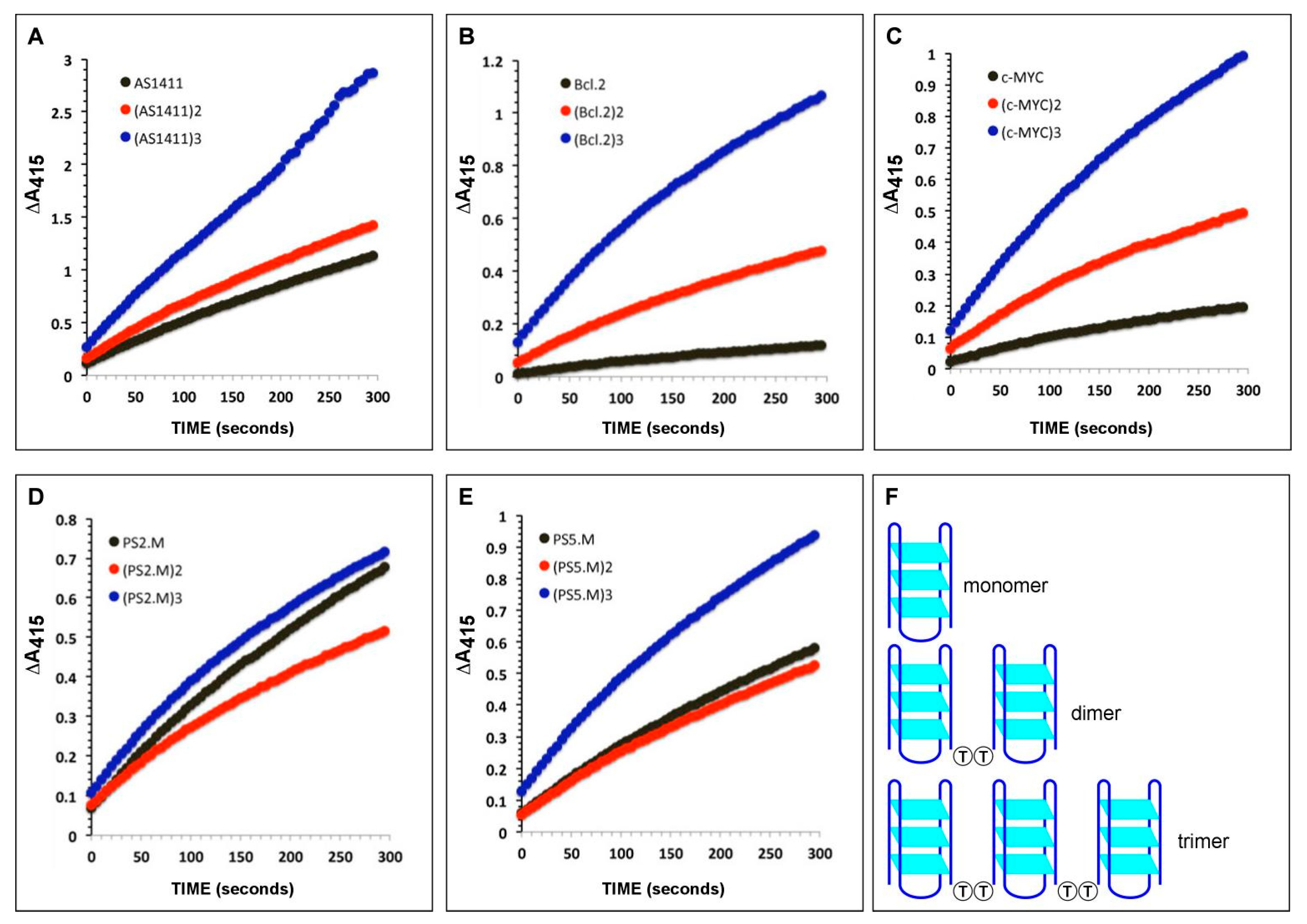
2.1. Design of Multimeric DNAzymes
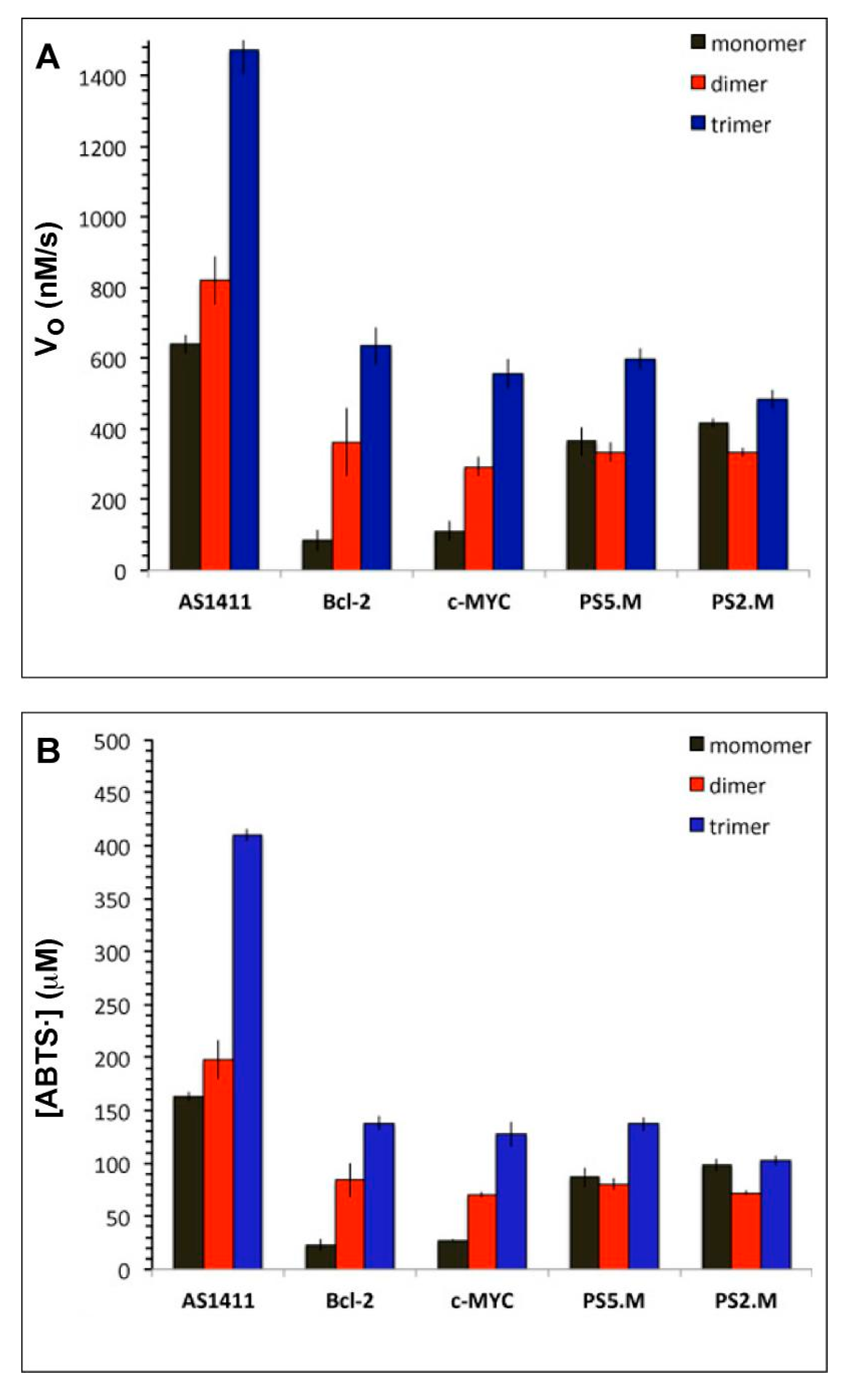
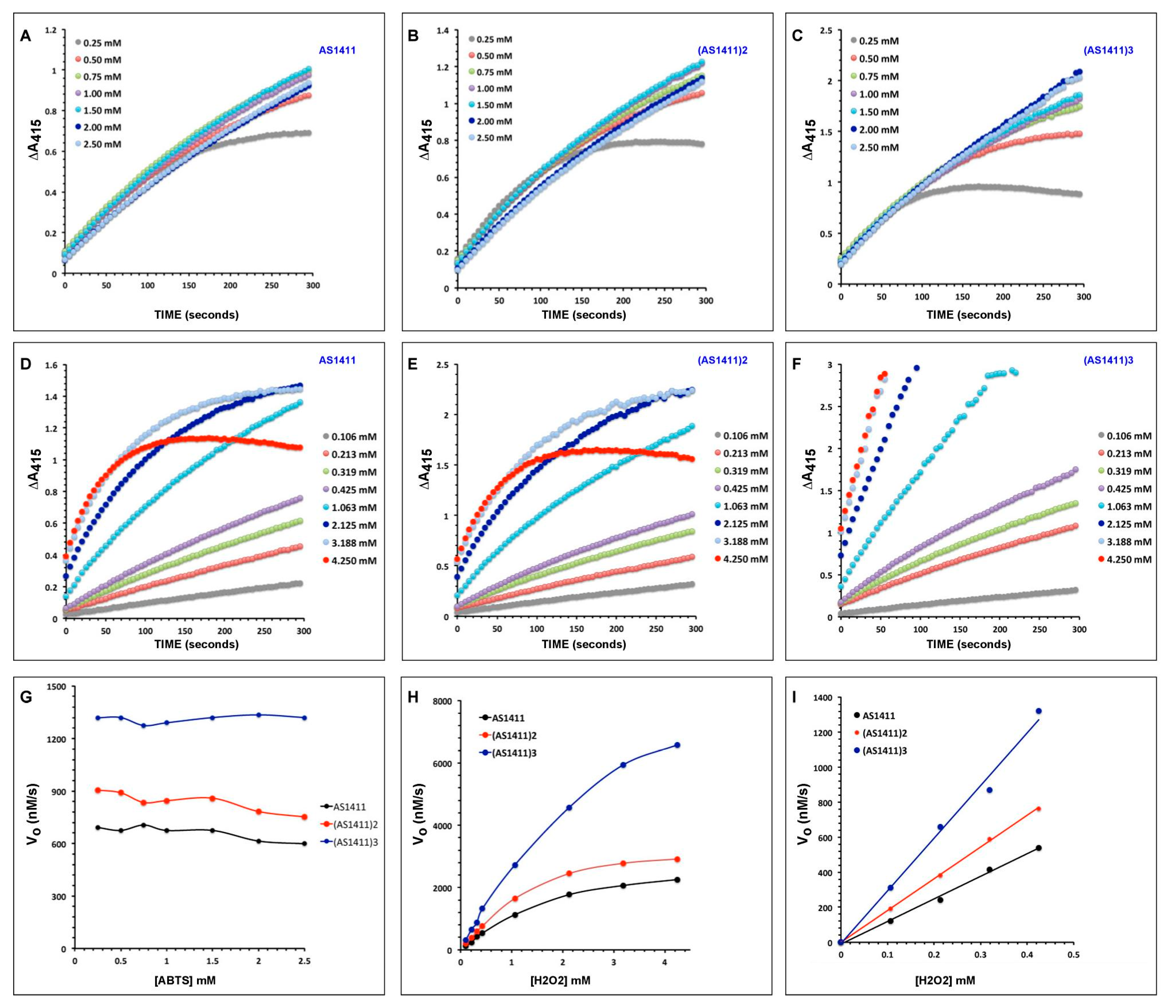
2.2. Catalytic Activities of DNAzymes
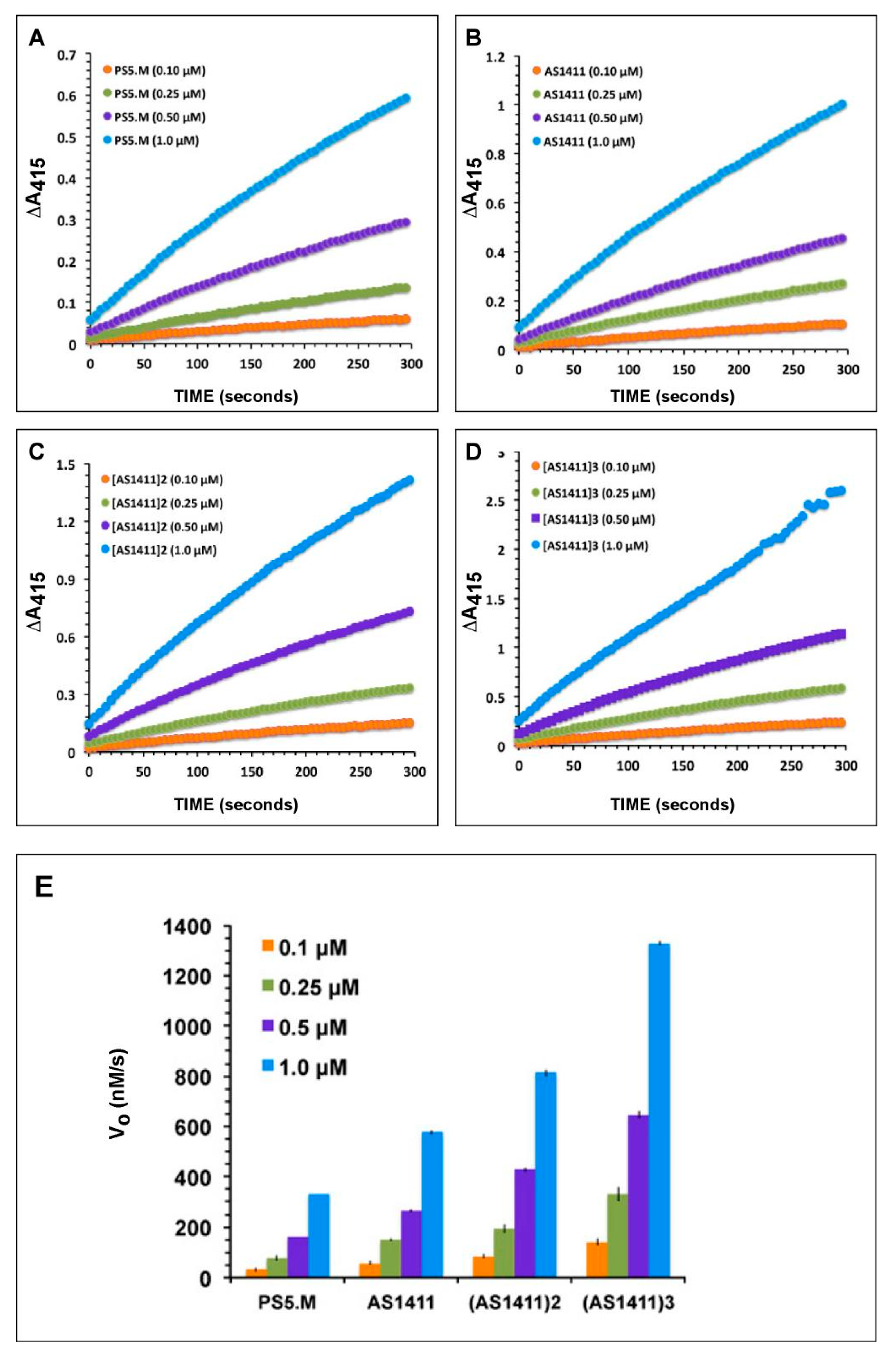
2.3. AS1411 Multimers Display High Activities at Low DNAzyme Concentrations
2.4. Effects of Reactants’ Concentrations on Activities of AS1411 Multimers
2.5. Determinants of Rate Enhancement by Multivalent DNAzymes
3. Materials and Methods
3.1. Reagents and DNA Oligonucleotides
3.2. Preparation of G4/hemin DNAzyme
3.3. Determination of G4/hemin DNAzyme Activity
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Singh, P.K.; Iqbal, N.; Sirohi, H.V.; Bairagya, H.R.; Kaur, P.; Sharma, S.; Singh, T.P. Structural basis of activation of mammalian heme peroxidases. Prog. Biophys. Mol. Biol. 2018, 133, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Sen, D. A catalytic DNA for porphyrin metallation. Nat. Struct. Biol. 1996, 3, 743–747. [Google Scholar] [CrossRef] [PubMed]
- Travascio, P.; Li, Y.; Sen, D. DNA-enhanced peroxidase activity of a DNA aptamer-hemin complex. Chem. Biol. 1998, 5, 505–517. [Google Scholar] [CrossRef] [Green Version]
- Kosman, J.; Juskowwiak, B. Peroxidase-mimicking DNAzymes for biosensing applications: A review. Anal. Chim. Acta 2011, 707, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Ge, B.; Sen, D.; Yu, H.Z. Functional DNA switches: Rational design and electrochemical Signaling. Chem. Soc. Rev. 2014, 43, 518–529. [Google Scholar] [CrossRef] [PubMed]
- Silverman, S.K. Catalytic DNA: Scope, applications, and biochemistry of deoxyribozymes. Trends Biochem. Sci. 2010, 41, 595–609. [Google Scholar] [CrossRef] [PubMed]
- Pelossof, G.; Tel-Vered, R.; Elbaz, J.; Willner, I. Amplified biosensing using the horseradish peroxidase-mimicking DNAzyme as an electrocatalyst. Anal. Chem. 2010, 82, 4396–4402. [Google Scholar] [CrossRef]
- Kong, D.M. Factors influencing the performance of G-quadruplex DNAzyme-based sensors. Methods 2013, 64, 199–204. [Google Scholar] [CrossRef]
- Keniry, M.A. A comparison of the association of spermine with duplex and quadruplex DNA by NMR. FEBS Lett. 2003, 542, 153–158. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Wang, X.; Zhang, J.; Xiao, Y.; Li, W.; Wang, J. Orderly microaggregates of G-/C-Rich oligonucleotides associated with spermine. Biomacromolecules 2011, 12, 747–756. [Google Scholar] [CrossRef]
- Qi, C.; Zhang, N.; Yan, J.; Liu, X.; Bing, T.; Mei, H.; Shangguan, D. Activity enhancement of G-quadruplex/hemin DNAzyme by spermine. RSC Adv. 2014, 4, 1441–1448. [Google Scholar] [CrossRef]
- Kong, D.M.; Xu, J.; Shen, H.X. Positive Effects of ATP on G-quadruplex-Hemin DNAzyme-mediated reactions. Anal. Chem. 2010, 82, 6148–6153. [Google Scholar] [CrossRef]
- Stefan, L.; Denat, F.; Monchaud, D. Insights into how nucleotide supplements enhance the peroxidase-mimicking DNAzyme activity of the G-quadruplex/hemin system. Nucleic Acids Res. 2012, 40, 8759–8772. [Google Scholar] [CrossRef] [Green Version]
- Kosman, J.; Zukowski, K.; Juskowiak, B. Comparison of characteristics and DNAzyme activity of G4/Hemin conjugates obtained via two hemin attachment methods. Molecules 2018, 23, 1400. [Google Scholar] [CrossRef]
- Xiao, L.; Zhou, Z.; Feng, M.; Tong, A.; Xiang, Y. Cationic peptide conjugation enhances the activity of peroxidase-mimicking DNAzymes. Bioconj. Chem. 2016, 27, 621–627. [Google Scholar] [CrossRef]
- Chang, T.; Gong, H.; Ding, P.; Liu, X.; Li, W.; Bing, T.; Cao, Z.; Shangguan, D. Activity enhancement of G-quadruplex/Hemin DNAzyme by flanking d(CCC). Chem. Eur. J. 2016, 22, 4015–4021. [Google Scholar] [CrossRef]
- Chen, J.; Guo, Y.; Zhou, J.; Ju, H. The effect of Adenine repeats on G-quadruplex/hemin peroxidase mimicking DNAzyme activity. Chem. Eur. J. 2017, 23, 4210–4215. [Google Scholar] [CrossRef]
- Li, W.; Li, Y.; Liu, Z.; Lin, B.; Yi, H.; Xu, F.; Nie, Z.; Yao, S. Insight into G-quadruplex-hemin DNAzyme/RNAzyme: Adjacent adenine as the intramolecular species for remarkable enhancement of enzymatic activity. Nucleic Acids Res. 2016, 44, 7373–7384. [Google Scholar] [CrossRef]
- Guo, Y.; Chen, J.; Cheng, M.; Monchaud, D.; Zhou, J.; Ju, H. A thermophilic tetramolecular G-quadruplex/Hemin DNAzyme. Angew. Chem. Int. Ed. 2017, 56, 16636–16640. [Google Scholar] [CrossRef] [PubMed]
- Stefan, L.; Denat, F.; Monchaud, D. Deciphering the DNAzyme activity of multimeric quadruplexes: Insights into their actual role in the telomerase activity evaluation assay. J. Am. Chem. Soc. 2011, 133, 20405–20415. [Google Scholar] [CrossRef]
- Yang, D.K.; Kuo, C.J.; Chen, L.C. Synthetic multivalent DNAzymes for enhanced hydrogen peroxide catalysis and sensitive colorimetric glucose detection. Anal. Chim. Acta 2015, 856, 96–102. [Google Scholar] [CrossRef]
- Liu, S.; Xu, N.; Tan, C.; Fang, W.; Tan, Y.; Jiang, Y. A sensitive colorimetric aptasensor based on trivalent peroxidase mimic DNAzyme and magnetic nanoparticles. Anal. Chim. Acta 2018, 1018, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Dong, S.; Wang, E. G-Quadruplex aptamers with peroxidase-like DNAzyme functions: Which is the best and how does it work? Chem. Asian J. 2009, 4, 918–922. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Li, D.; Shang, H. General peroxidase activity of a parallel G-quadruplex-hemin DNAzyme formed by Pu39WT—A mixed G-quadruplex forming sequence in the Bcl-2 P1 promoter. Chem. Cent. J. 2014, 8, 43. [Google Scholar] [CrossRef] [PubMed]
- Girvan, A.C.; Teng, Y.; Casson, L.K.; Thomas, S.D.; Juliger, S.; Ball, M.W.; Klein, J.B.; Pierce, W.M., Jr.; Barve, S.S.; Bates, J.P. AGRO100 inhibits activation of nuclear factor-κB (NF-κB) by forming a complex with NF-κB essential modulator (NEMO) and nucleolin. Mol. Cancer Ther. 2006, 5, 1790–1799. [Google Scholar] [CrossRef]
- Kong, D.M.; Yang, W.; Wu, J.; Li, C.X.; Shen, H.X. Structure–function study of peroxidase-like G-quadruplex-hemin complexes. Analyst 2010, 135, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Simonsson, T.; Pecinka, P.; Kubista, M. DNA tetraplex formation in the control region of c-myc. Nucleic Acids Res. 1998, 26, 1167–1172. [Google Scholar] [CrossRef] [Green Version]
- Dai, J.; Chen, D.; Jones, R.A.; Hurley, L.H.; Yang, D. NMR solution structure of the major G-quadruplex structure formed in the human BCL2 promoter region. Nucleic Acids Res. 2006, 34, 5133–5144. [Google Scholar] [CrossRef]
- Dai, J.; Dexheimer, T.S.; Chen, D.; Carver, M.; Ambrus, A.; Jones, R.A.; Yang, D. An intramolecular G-quadruplex structure with mixed parallel/antiparallel G-strands formed in the human BCL-2 promoter region in solution. J. Am. Chem. Soc. 2006, 128, 1096–1098. [Google Scholar] [CrossRef]
- Liu, Z.; He, K.; Li, W.; Liu, X.; Xu, X.; Nie, Z.; Yao, S. DNA G-quadruplex-based assay of enzyme activity. Methods Mol. Biol. 2017, 1500, 133–151. [Google Scholar]
- Kwok, C.K.; Merrick, C.J. G-quadruplexes: Prediction, characterization, and biological application. Trends Biotechnol. 2017, 35, 997–1013. [Google Scholar] [CrossRef]
- Kosman, J.; Juskowwiak, B. Hemin/G-quadruplex structure and activity alteration induced by magnesium cations. Int. J. Biol. Macromol. 2016, 85, 555–564. [Google Scholar] [CrossRef]
- Rachwal, P.A.; Brown, T.; Fox, K.R. Effect of G-Tract length on the topology and stability of intramolecular DNA quadruplexes. Biochemistry 2007, 46, 3036–3044. [Google Scholar] [CrossRef]
- Bugaut, A.; Balasubramanian, S. A Sequence-independent study of the influence of short loop lengths on the stability and topology of intramolecular DNA G-Quadruplexes. Biochemistry 2008, 47, 689–697. [Google Scholar] [CrossRef]
- Phan, A.T.; Modi, Y.S.; Patel, D.J. Propeller-type parallel-stranded G-quadruplexes in the human c-myc promoter. J. Am. Chem. Soc. 2004, 126, 8710–8716. [Google Scholar] [CrossRef]
- Li, T.; Shi, L.; Wang, E.; Dong, S. Multifunctional G-quadruplex aptamers and their application to protein detection. Chem. Eur. J. 2009, 15, 1036–1042. [Google Scholar] [CrossRef]
- Liu, W.; Zhu, H.; Zheng, B.; Cheng, S.; Fu, Y.; Li, W.; Lau, T.C.; Liang, H. Kinetics and mechanism of G-quadruplex formation and conformational switch in a G-quadruplex of PS2.M induced by Pb2+. Nucleic Acids Res. 2012, 40, 4229–4236. [Google Scholar] [CrossRef]
- Cheng, X.H.; Liu, X.J.; Bing, T.; Cao, Z.H.; Shangguan, D.H. General peroxidase activity of G-quadruplex–Hemin complexes and its application in ligand screening. Biochemistry 2009, 48, 7817–7823. [Google Scholar] [CrossRef]
- Nakayama, S.; Wang, J.; Sintim, H.O. DNA-based peroxidation catalyst–what is the exact role of topology on catalysis and is there a special binding site for catalysis? Chemistry 2011, 17, 5691–5698. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DNAzyme | Sequence |
|---|---|
| AS1411 | GGTGGTGGTGGTTGTGGTGGTGGTGG |
| Bcl-2 | GGGCGCGGGAGGAAGGGGGCGGG |
| c-MYC | GAGGGTGGGGAGGGTGGGGAAG |
| PS5.M | GTGGGTCATTGTGGGTGGGTGTGG |
| PS2.M | GTGGGTAGGGCGGGTTGG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adeoye, R.I.; Osalaye, D.S.; Ralebitso-Senior, T.K.; Boddis, A.; Reid, A.J.; Fatokun, A.A.; Powell, A.K.; Malomo, S.O.; Olorunniji, F.J. Catalytic Activities of Multimeric G-Quadruplex DNAzymes. Catalysts 2019, 9, 613. https://doi.org/10.3390/catal9070613
Adeoye RI, Osalaye DS, Ralebitso-Senior TK, Boddis A, Reid AJ, Fatokun AA, Powell AK, Malomo SO, Olorunniji FJ. Catalytic Activities of Multimeric G-Quadruplex DNAzymes. Catalysts. 2019; 9(7):613. https://doi.org/10.3390/catal9070613
Chicago/Turabian StyleAdeoye, Raphael I., Dunsin S. Osalaye, Theresia K. Ralebitso-Senior, Amanda Boddis, Amanda J. Reid, Amos A. Fatokun, Andrew K. Powell, Sylvia O. Malomo, and Femi J. Olorunniji. 2019. "Catalytic Activities of Multimeric G-Quadruplex DNAzymes" Catalysts 9, no. 7: 613. https://doi.org/10.3390/catal9070613