Appendix A. Model Development
Appendix A1. Single-Substrate Kinetic Model
In an earlier work [
5], a model was developed to predict the time course of the laccase-catalyzed oxidation of phenol at low concentrations. It was developed based on the reactions of laccase illustrated in
Figure A1.
Figure A1.
The catalytic cycle of laccase in which enzyme in the native state, E, is oxidized by aqueous oxygen, O
2, to produce activated enzyme, E*, which reacts with substrate, S, to produce phenoxy radicals, S·. Catalytically slow (E
RO) or non-productive (E
P, E
X) states of laccase involved in side reactions are also shown (Source: [
5]).
Figure A1.
The catalytic cycle of laccase in which enzyme in the native state, E, is oxidized by aqueous oxygen, O
2, to produce activated enzyme, E*, which reacts with substrate, S, to produce phenoxy radicals, S·. Catalytically slow (E
RO) or non-productive (E
P, E
X) states of laccase involved in side reactions are also shown (Source: [
5]).
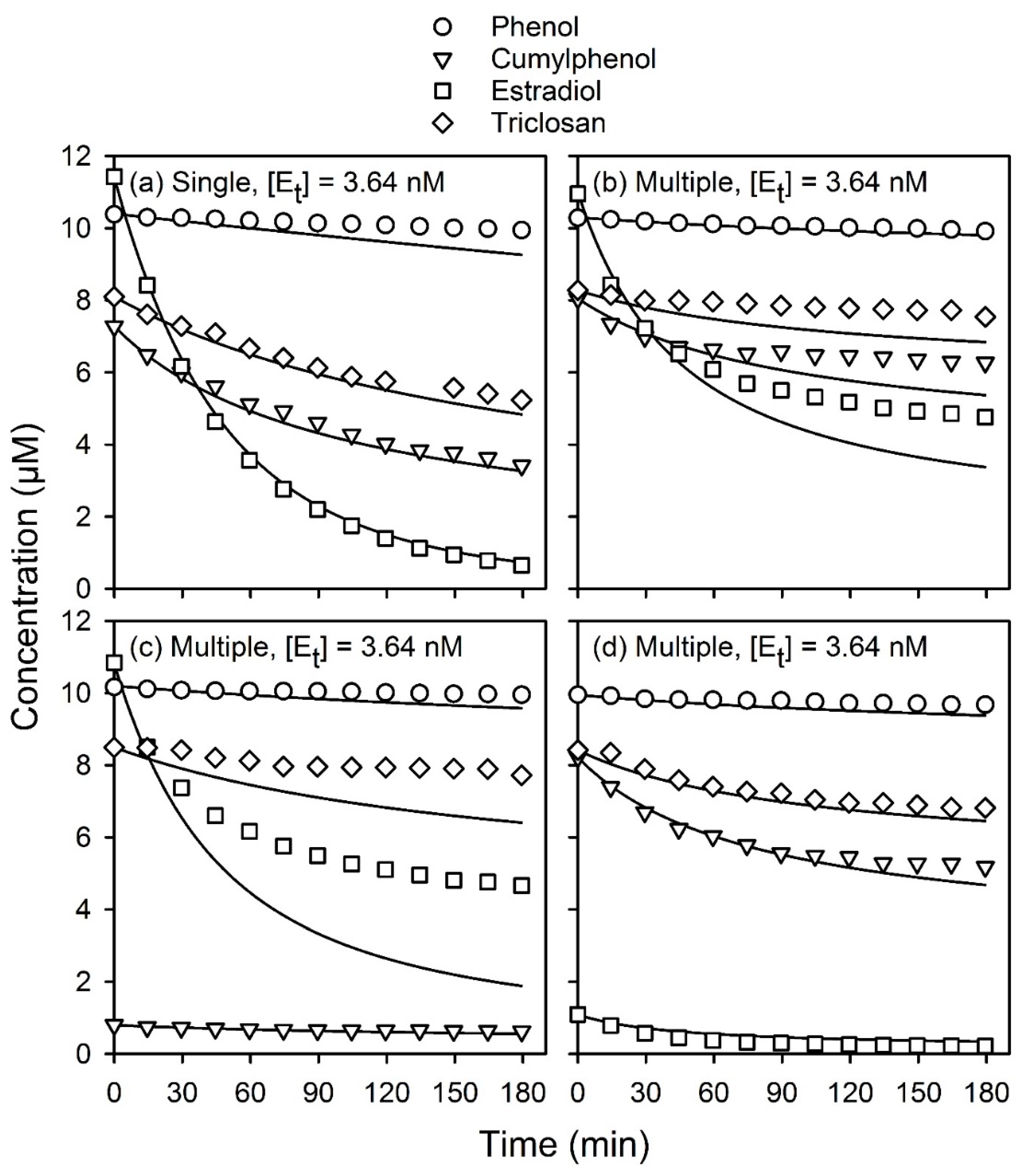
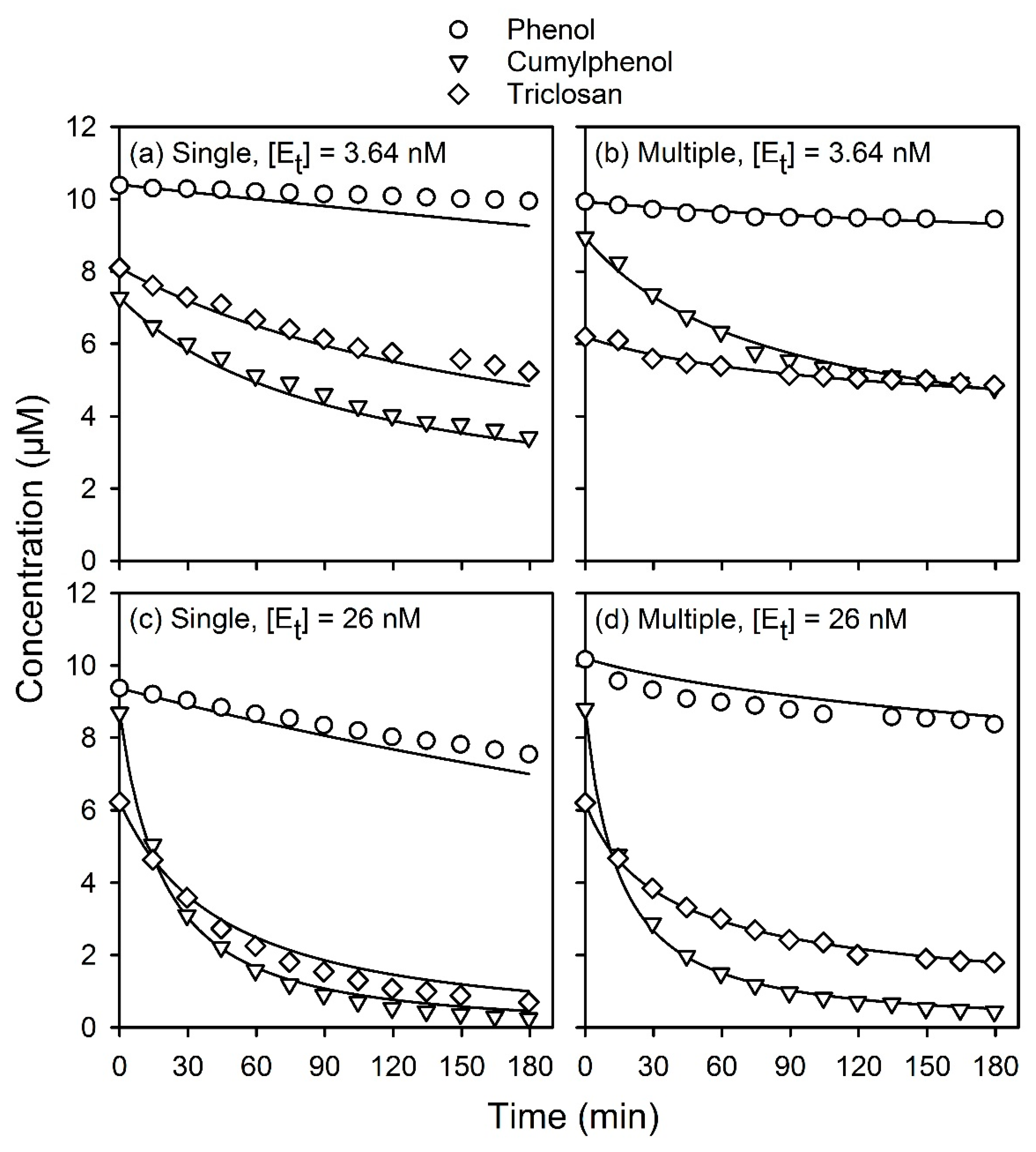
In a subsequent work, the general applicability of the model was demonstrated by applying it to three other phenolic substrates at micromolar and sub-micromolar concentrations. For these three substrates, including estradiol, cumylphenol, and triclosan, it was necessary to incorporate an expression accounting for enzyme inactivation into the model [
6], where it was assumed that inactivation results from the interaction of substrate radicals produced during the reaction, S, with the active enzyme. This model was used to predict the time course of reactions of reaction mixtures containing a single phenolic substrate and, hence, it is termed here the “single-substrate” model.
The general model consists of four differential and one mass balance equations, as follows:
These equations describe the transient kinetics of laccase-catalyzed oxidation of a phenolic substrate in an aqueous reaction mixture with initial concentrations of substrate [S]0, oxygen [O2]0, and enzyme [Et]. Equation (A1) describes the rate of change of oxygen concentration due to its consumption in the oxidation reaction and its replenishment by mass transfer for situations where the system is open to the air. Equation (A2) describes the kinetics of phenolic substrate oxidation, and Equation (A3) describes the rate of concentration change of enzyme in the reduced form, E. The rate of formation of inactive enzyme is described by Equation (A4), where the term [Et] − [Ei] represents the total active enzyme in the mixture at any instant. Finally, the enzyme mass balance, Equation (A5), is used to estimate the concentration of enzyme in the oxidized state, E*, at any instant. Variable definitions and their dimensions are listed in the Nomenclature above. As reported previously, when solving these equations, it can be assumed that [O2]0 = [O2]sat for reaction mixtures that were in equilibrium with ambient air before reaction initiation at t = 0. Furthermore, it can be assumed that all enzyme is in the oxidized form at the start of the reaction (i.e., [E] = 0 and [E*] = [Et]).
As explained elsewhere [
6], the model has been calibrated for single-substrate reactions of phenol, estradiol, cumylphenol, and triclosan. The kinetic parameters for each of these substrates are summarized in
Table A1. The model was shown to be able to predict the time course of substrate concentrations for reactions conducted under conditions that were significantly outside of the range of its calibration. Overall, the model very accurately described the time course of reactions over several orders of magnitude of enzyme and substrate concentrations for reactions conducted over 3-h periods and longer. The kinetic parameters listed in
Table A1 were used in the present study when modeling reactions of individual or mixed substrates.
Table A1.
Modeled average rates of substrate oxidation (and relative rates with respect to phenol) during the first minute of reactions conducted at high and low enzyme concentrations at pH 5.0 and 25 °C. Also shown is the estimated percentage of enzyme inactivated over the first minute of the reaction modeled under conditions of high substrate and high enzyme concentrations where inactivation would be greatest.
Table A1.
Modeled average rates of substrate oxidation (and relative rates with respect to phenol) during the first minute of reactions conducted at high and low enzyme concentrations at pH 5.0 and 25 °C. Also shown is the estimated percentage of enzyme inactivated over the first minute of the reaction modeled under conditions of high substrate and high enzyme concentrations where inactivation would be greatest.
| Parameters | Parameter Values Used to Model Substrate Kinetics |
|---|
| Estradiol | Cumylphenol | Triclosan | Phenol |
|---|
| (min−1) | 1.05 × 10−2 |
| (μM−1·min−1) | 5.44 × 10−1 |
| (μM1−α−β·min−1) | 6.75 | 0.879 | 0.271 | 4.17 × 10−3 |
| (dimensionless) | 1.00 | 0.88 | 0.87 | 0.51 |
| (dimensionless) | 1.18 | 1.23 | 1.39 | 1.44 |
| ki (μM−0.5·min−0.5) | 1.87 × 10−2 | 5.11 × 10−2 | 2.99 × 10−2 | 0 |
Appendix A2. Multi-Substrate Kinetic Model
The single-substrate kinetic model presented above was adapted to predict the time course of reactions of multiple substrates in a reaction mixture, as will be described below. Note that Equations (A1) and (A5) do not require adaptation because they are substrate-independent.
For a mixture of multiple substrates, equations must be written and simultaneously solved for each substrate to evaluate their respective rates of concentration change with time. That is, for each individual substrate,
n, in a mixture of
N substrates, Equation (A2) can be adapted as follows to calculate the rate of concentration change of substrate
n,
[Sn]:
Therefore, for a mixture of N substrates, there will be N independent equations where each substrate, Sn (i.e., where n = 1 to N), will have its own rate equation and kinetic parameters, , , and .
Equation (A3) can then be adapted to account for the formation of reduced enzyme, E, in an additive manner through the parallel reactions of E* with each substrate, S
n, as follows:
Similarly, the rate of enzyme inactivation in a mixture can be calculated as the sum of the rates of inactivation arising from the oxidation of each substrate. In doing this, it is assumed that the free-radical products, S
n, arising from the reactions of the different substrates, S
n, do not interact with each other in such a way that inactivation is reduced (through radical–radical reactions) or increased (through product formation). The validity of this assumption is tested in the present work. The rate of change of concentration of inactive enzyme,
[Ei], may be expressed as
Therefore, the kinetic model for the laccase-catalyzed oxidation of a mixture of N substrates consists of Equations (A1), (A5), (A7), and (A8), along with Equation (A6) adapted N times for the N substrates. The initial conditions for the numerical solution of model equations are [O2] = [O2]0, [Sn] = [Sn]0, [E*] = [Et], [E] = 0, and [Ei]0 = 0 at t = 0.
For the purposes of this study, the model coefficients used included the kinetic constants listed in
Table A1 for the four phenolic substrates [
6], an oxygen mass-transfer coefficient,
, of 1.05 × 10
−2 min
−1, and
[O2]0 equal to the saturation concentration,
[O2]sat, of 250 µM [
5]. The methods used to calibrate these parameters are reported elsewhere [
5,
6].
Appendix B. Classification of Substrates
For the purposes of this study, substrates were classified as either fast, intermediate, or slow with respect to their relative rates of oxidation. Note that, due to the non-linear nature of Equation (2), it was not possible to directly compare the kinetic parameters shown in
Table A1 in order to carry out such a classification. Instead, they were classified based on estimates of the rate of substrate oxidation,
(μM·min
−1), and modeled under fixed reaction conditions of high and low substrate and enzyme concentrations. Thus, the model equations summarized in
Section A1 were solved using the initial conditions listed in the first column of
Table A2 in order to determine the average reaction rate in the first minute of the reaction. Note that these modeled initial conditions of substrate and enzyme concentrations were the same as those used in kinetic experiments in the present study (i.e., see
Figure 4,
Figure 5 and
Figure 6 in the main body).
Table A2.
Modeled average rates of substrate oxidation (and relative rates with respect to phenol) during the first minute of reactions conducted at high and low enzyme concentrations at pH 5.0 and 25 °C. Also shown is the estimated percentage of enzyme inactivated over the first minute of the reaction modeled under conditions of high substrate and high enzyme concentrations where inactivation would be greatest.
Table A2.
Modeled average rates of substrate oxidation (and relative rates with respect to phenol) during the first minute of reactions conducted at high and low enzyme concentrations at pH 5.0 and 25 °C. Also shown is the estimated percentage of enzyme inactivated over the first minute of the reaction modeled under conditions of high substrate and high enzyme concentrations where inactivation would be greatest.
| Reaction Conditions or Classification | Substrate |
|---|
| Estradiol | Cumylphenol | Triclosan | Phenol |
|---|
Rate of Substrate Oxidation (μM·min−1)
(Relative Rate with Respect to Phenol; Dimensionless) |
|---|
| [S]0 = 1 μM, [Et] = 3.64 nM | 2.31 × 10−2 (85) | 6.17 × 10−3 (23) | 2.04 × 10−3 (7.5) | 2.73 × 10−4 (1) |
| [S]0 = 1 μM, [Et] = 26.0 nM | 1.52 × 10−1 (235) | 3.42 × 10−2 (53) | 1.12 × 10−2 (17) | 6.48 × 10−4 (1) |
| [S]0 = 10 μM, [Et] = 3.64 nM | 2.11 × 10−1 (33) | 8.85 × 10−2 (14) | 4.59 × 10−2 (7.1) | 6.50 × 10−3 (1) |
| [S]0 = 10 μM, [Et] = 26.0 nM | 1.43 (80) | 5.01 × 10−1 (28) | 2.54 × 10−1 (14) | 1.78 × 10−2 (1) |
| Classification according to relative rate of oxidation | Fast | Intermediate | Intermediate | Slow |
| Laccase inactivation (%) |
| [S]0 = 10 μM, [Et] = 26.0 nM | 2.2 | 3.6 | 1.5 | 0 |
| Classification according to relative tendency to inactivate | Moderate | High | Moderate | Non-Inactivating |
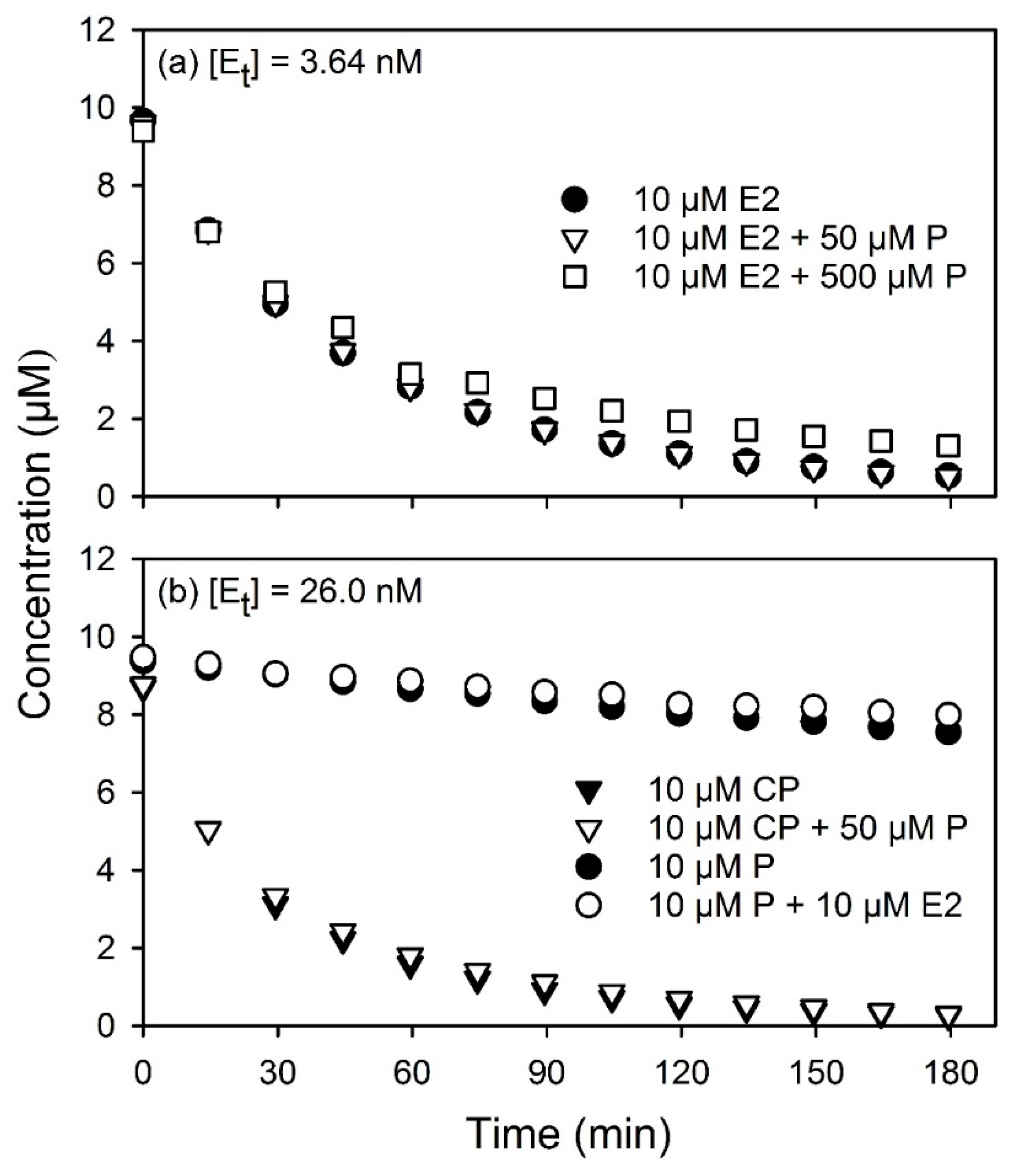
The calculated average reaction rates were tabulated for all four substrates in
Table A2. For ease of comparison, all rates in
Table A2 were also expressed relative to the rates of the slowest substrate, phenol. Over the range of conditions modeled, it can be seen that the rate of oxidation of estradiol was between 33 and 235 times faster than phenol under the same conditions. Cumylphenol was the next fastest substrate, with relative oxidation rates between 14 and 53 times that of phenol. Triclosan had relative rates between 7.1 and 17 times faster than phenol, and was between 2 and 3 times slower than cumylphenol. For these reasons, estradiol was deemed a “fast” substrate, cumylphenol and triclosan were deemed “intermediate” substrates, and phenol was deemed a “slow” substrate.
Similarly, for the purposes of this study, the substrates were also classified in terms of their tendency to inactivate laccase during the catalytic reaction. In order to classify them, the model was used to predict the quantity of inactivation that would occur during the first minute of reaction under a condition of high enzyme and substrate concentrations where the rate of inactivation would be greatest [
6]. As can be seen in the bottom rows of
Table A2, with
[S]0 = 10 μM and
[Et] = 26.0 nM, it was predicted that 3.6%, 2.2%, 1.5%, and 0% of the enzyme supplied to the reaction would be inactivated in the first minute when oxidizing cumylphenol, estradiol, triclosan, and phenol, respectively. As such, in terms of their relative tendency to inactivate laccase during the catalytic reaction, cumylphenol was deemed to be “high”, estradiol and triclosan were deemed to be “moderate”, and phenol was deemed to be “non-inactivating”. Note that phenol has been observed to inactivate laccase, but only at elevated concentrations in excess of 500 μM [
14]. As such, for the purpose of this study, where very low concentrations of phenol were used, this substrate is considered to be “non-inactivating”.
Appendix C. Investigation of Impacts of Secondary Substrates Using the Multi-Substrate Model
The multi-substrate kinetic model can be used to gain insights into the impacts of secondary substrates on the rate of conversion of a target substrate, especially under conditions that may be difficult to explore experimentally. For example, simulations were conducted here to investigate the impacts of hypothetical secondary substrates on the conversion of cumylphenol (intermediate rate, highly inactivating) and estradiol (fast substrate, moderately inactivating) as target substrates. In these simulations, the impacts of hypothetical substrates, denoted as X, that were kinetically similar to the target substrates, were investigated. This was done by setting the kinetic parameters of the hypothetical secondary substrate (i.e.,
,
and
) equal to those of the target substrates (see
Table A1). The effect of the concentration of this hypothetical secondary substrate was then investigated by varying its initial concentration,
[SX]0, in simulations. Furthermore, the effect of the tendency of the hypothetical secondary substrate to inactivate the enzyme was investigated by varying its inactivation rate constant,
.
First, in order to evaluate the impact of increasing the concentration of kinetically similar but non-inactivating substrates, simulations were performed in which the initial concentration of the secondary substrate, X, was varied, but where its inactivation rate constant,
, was set to 0. The results of simulations are shown in
Figure A2a and
Figure A3a for cumylphenol and estradiol, respectively, where the initial concentration of the target substrate was equal to 10 μM, and where the effect of the initial concentration of the hypothetical secondary substrate was simulated at 0, 10, and 20 μM.
Figure A2.
Predicted impacts of a hypothetical secondary substrate, X, on the kinetics of oxidation of the target substrate, cumylphenol (CP), and the corresponding fractions of laccase in the oxidized, E*, and inactivated, Ei, states over time: (a,b) effect of varying the initial concentration, [SX]0, of the secondary substrate, X, when it is non-inactivating (i.e., = 0); (c,d) effect of varying the inactivation rate constant, , of the secondary substrate, X, for a fixed initial secondary substrate concentration, [SX]0, of 10 μM. For all simulations, the initial concentration of the target substrate was 10 μM and the total enzyme supplied to the reaction, [Et], was 26 nM. The fractions of enzyme in the E* and Ei states over time are expressed as a percentage of the total enzyme, [Et]. The fraction of enzyme in the native state, E, is not shown.
Figure A2.
Predicted impacts of a hypothetical secondary substrate, X, on the kinetics of oxidation of the target substrate, cumylphenol (CP), and the corresponding fractions of laccase in the oxidized, E*, and inactivated, Ei, states over time: (a,b) effect of varying the initial concentration, [SX]0, of the secondary substrate, X, when it is non-inactivating (i.e., = 0); (c,d) effect of varying the inactivation rate constant, , of the secondary substrate, X, for a fixed initial secondary substrate concentration, [SX]0, of 10 μM. For all simulations, the initial concentration of the target substrate was 10 μM and the total enzyme supplied to the reaction, [Et], was 26 nM. The fractions of enzyme in the E* and Ei states over time are expressed as a percentage of the total enzyme, [Et]. The fraction of enzyme in the native state, E, is not shown.
Figure A3.
Predicted impacts of a hypothetical secondary substrate, X, on the kinetics of oxidation of the target substrate, estradiol (E2), and the corresponding fractions of laccase in the oxidized, E*, and inactivated, Ei, states over time: (a,b) effect of varying the initial concentration, [SX]0, of the secondary substrate, X, when it is non-inactivating (i.e., = 0); (c,d) effect of varying the inactivation rate constant, , of the secondary substrate, X, for a fixed initial secondary substrate concentration, [SX]0, of 10 μM. For all simulations, the initial concentration of the target substrate was 10 μM and the total enzyme supplied to the reaction, [Et], was 3.64 nM. The fractions of enzyme in the E*and Ei states over time are expressed as a percentage of the total enzyme, [Et]. The fraction of enzyme in the native state, E, is not shown.
Figure A3.
Predicted impacts of a hypothetical secondary substrate, X, on the kinetics of oxidation of the target substrate, estradiol (E2), and the corresponding fractions of laccase in the oxidized, E*, and inactivated, Ei, states over time: (a,b) effect of varying the initial concentration, [SX]0, of the secondary substrate, X, when it is non-inactivating (i.e., = 0); (c,d) effect of varying the inactivation rate constant, , of the secondary substrate, X, for a fixed initial secondary substrate concentration, [SX]0, of 10 μM. For all simulations, the initial concentration of the target substrate was 10 μM and the total enzyme supplied to the reaction, [Et], was 3.64 nM. The fractions of enzyme in the E*and Ei states over time are expressed as a percentage of the total enzyme, [Et]. The fraction of enzyme in the native state, E, is not shown.
The corresponding predicted distributions of enzyme over time for these simulations are shown in
Figure A2b and
Figure A3b. As can be seen in
Figure A2a, the impact of a kinetically similar but non-inactivating substrate on the predicted rate of oxidation of cumylphenol (intermediate rate, highly inactivating) is very little. In contrast, similar simulations using estradiol as the target substrate indicate that a non-inactivating but kinetically similar secondary substrate can have a significant impact on the rate of oxidation of the target substrate, and that this impact increases with concentration, as shown in
Figure A3a.
As can be seen in
Figure A2b and
Figure A3b, the presence of the non-inactivating secondary substrates is predicted to have only a minor impact on the rate of accumulation of inactive enzyme over time, even when present in significant concentration. However, these figures show that such secondary substrates significantly impact on the fraction of enzyme in the oxidized state, E*, especially in the early stages of the reaction. Furthermore, as shown in
Figure A3b, the impact on the enzyme distribution is especially significant when the target and secondary substrates are both kinetically very fast and particularly when the secondary substrate is at a relatively high concentration with respect to the target substrate.
In a second set of simulations, the impact of secondary substrates that are kinetically similar to the target substrate but with different tendencies to inactivate the enzyme were evaluated. The results are shown in
Figure A2c and
Figure A3c. Simulations are shown for reactions of the 10-μM target substrate alone and in the presence of 10 μM of a hypothetical secondary substrate that is non-inactivating (
= 0) or that have the same or twice the tendency to inactivate the enzyme as compared to the target substrate. The predicted distributions of enzyme over time for these simulations are shown in
Figure A2d and
Figure A3d. From these simulations, it is clear that the impact of an inactivating secondary substrate on the rate of oxidation of the target substrate is very significant. Furthermore, the magnitude of this impact is important for both intermediate and fast substrates. This is further demonstrated by the significant impact that the inactivating substrate has on the quantity of inactive enzyme, E
i, accumulated over time, and a corresponding reduction in the amount of enzyme in the E* state that is available to oxidize the substrates. The overall effect of this simultaneous accumulation of E
i and depletion of E* is a significant negative impact on the rate of oxidation of the target substrate. Similar simulations will show that this negative impact worsens with an increasing concentration of the secondary substrate (not shown).




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}








