Clostridium sp. as Bio-Catalyst for Fuels and Chemicals Production in a Biorefinery Context

 , , , and
, , , and
Abstract
:1. Introduction
2. Clostridium sp.
2.1. General Aspects
2.2. Workhorse Species
2.2.1. Clostridium carboxidivorans
2.2.2. Clostridium ragsdalei
2.2.3. Clostridium ljungdhali
2.2.4. Clostridium kluyveri
2.2.5. Clostridium beijerinckii
2.2.6. Clostridium saccharobutylicum
2.2.7. Clostridium saccharoperbutylacetonicum
2.2.8. Clostridium thermocellum
2.2.9. Clostridium phytofermentans
2.2.10. Clostridium acetobutylicum
2.2.11. Clostridium butyricum
2.2.12. Clostridium cellulolyticum
2.2.13. Clostridium cellulovorans
2.2.14. Clostridium pasteurianum
2.2.15. Clostridium perfringens
2.2.16. Clostridium tyrobutyricum
2.2.17. Other Species
2.3. Fermentation and Biochemical Pathways
2.3.1. Acetone-Butanol-Ethanol (ABE) Fermentation
2.3.2. Isopropanol-Butanol-Ethanol (IBE) Fermentation
2.3.3. Hexanol-Butanol-Ethanol (HBE) Fermentation
2.3.4. Embden-Meyerhof-Parnas (EMP) Pathway and Pentose Phosphate (PP) Pathway
2.3.5. Wood–Ljungdahl Pathway
3. Feedstock for Anaerobic Processes
3.1. Glycerol
3.2. Molasses
3.3. Corn Steep Liquor
3.4. Lignocellulosic Biomass
3.5. Syngas
4. Biomolecules Produced by Clostridium sp.
4.1. Ethanol
4.2. Butanol
4.3. Acetone
4.4. 1,3-Propanediol
4.5. Other Biomolecules
5. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Earth Overshoot Day Earth Overshoot Day 201 is August 1. Available online: https://www.overshootday.org/newsroom/press-release-july-2018-english/ (accessed on 7 January 2019).
- Intergovernmental Panel on Climate Change. Climate Change 2014: Synthesis Report; Chapter Observed Changes and Their Causes; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2014; ISBN 9789291691432. [Google Scholar]
- Cherubini, F. The biorefinery concept: Using biomass instead of oil for producing energy and chemicals. Energy Convers. Manag. 2010, 51, 1412–1421. [Google Scholar] [CrossRef]
- Sustainable Development Goals The Sustainable Development Agenda. Available online: https://www.un.org/sustainabledevelopment/development-agenda/ (accessed on 15 January 2019).
- Drosg, B.; Braun, R.; Bochmann, G.; Al Saedi, T. Analysis and characterisation of biogas feedstocks. In The Biogas Handbook: Science, Production and Applications; Murphy, J.D., Baxter, D., Eds.; Woodhead Publishing: Sawston/Cambridge, UK, 2013; pp. 52–84. ISBN 9780857097415. [Google Scholar]
- Al Seadi, T.; Rutz, D.; Janssen, R.; Drosg, B. Biomass resources for biogas production. In The Biogas Handbook: Science, Production and Applications; Murphy, J.D., Baster, D., Eds.; Woodhead Publishing: Sawston/Cambridge, UK, 2013; pp. 19–51. ISBN 9780857097415. [Google Scholar]
- Shen, Y.; Jarboe, L.; Brown, R.; Wen, Z. A thermochemical–biochemical hybrid processing of lignocellulosic biomass for producing fuels and chemicals. Biotechnol. Adv. 2015, 33, 1799–1813. [Google Scholar] [CrossRef] [PubMed]
- Latif, H.; Zeidan, A.A.; Nielsen, A.T.; Zengler, K. Trash to treasure: Production of biofuels and commodity chemicals via syngas fermenting microorganisms. Curr. Opin. Biotechnol. 2014, 27, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Murphy, J.D.; Korres, N.E.; Singh, A.; Smyth, B.; Nizami, A.-S.; Thamsiriroj, T. The Potential for Grass Biomethane as a Biofuel: Compressed Biomethane Generated from Grass, Utilised as a Transport Biofuel; Environmental Protection Authority, Ireland: Wexford, Ireland, 2011; ISBN 9781840954272. [Google Scholar]
- Hassan, S.S.; Willians, G.A.; Jaiswal, A.K. Moving towards the second generation of lignocellulosic biorefineries in the EU: Drivers, challenges, and opportunities. Renew. Sustain. Energy Rev. 2018, 101, 590–599. [Google Scholar] [CrossRef]
- International Energy Agency (IEA) Bioenergy. IEA Bioenergy Task 42 Biorefining; IEA: Paris, France, 2014. [Google Scholar]
- Parada, M.P.; Asveld, L.; Osseweijer, P.; Duque, J.A.P. An approach for incorporating sustainability in early stages of biorefinery design. In Proceedings of the European Biomass Conference and Exhibition, Stockholm, Sweden, 12–15 June 2017; pp. 105–123. [Google Scholar]
- Parajuli, R.; Dalgaard, T.; Jørgensen, U.; Adamsen, A.P.S.; Knudsen, M.T.; Birkved, M.; Gylling, M.; Schjørring, J.K. Biorefining in the prevailing energy and materials crisis: A review of sustainable pathways for biorefinery value chains and sustainability assessment methodologies. Renew. Sustain. Energy Rev. 2015, 43, 244–263. [Google Scholar] [CrossRef]
- Liou, J.S.C.; Balkwill, D.L.; Drake, G.R.; Tanner, R.S. Clostridium carboxidivorans sp. nov., a solvent-producing clostridium isolated from an agricultural settling lagoon, and reclassification of the acetogen Clostridium scatologenes strain SL1 as Clostridium drakei sp. nov. Int. J. Syst. Evol. Microbiol. 2005, 55, 2085–2091. [Google Scholar] [CrossRef]
- Daniell, J.; Köpke, M.; Simpson, S.D. Commercial biomass syngas fermentation. Energies 2012, 5, 5372–5417. [Google Scholar] [CrossRef]
- Schiel-bengelsdorf, B.; Dürre, P. Pathway engineering and synthetic biology using acetogens. FEBS Lett. 2012, 586, 2191–2198. [Google Scholar] [CrossRef]
- Kopke, M.; Held, C.; Hujer, S.; Liesegang, H.; Wiezer, A.; Wollherr, A.; Ehrenreich, A.; Liebl, W.; Gottschalk, G.; Durre, P. Clostridium ljungdahlii represents a microbial production platform based on syngas. Proc. Natl. Acad. Sci. USA 2010, 107, 13087–13092. [Google Scholar] [CrossRef]
- Lawson, P.A.; Rainey, F.A. Proposal to restrict the genus Clostridium prazmowski to Clostridium butyricum and related species. Int. J. Syst. Evol. Microbiol. 2016, 66, 1009–1016. [Google Scholar] [CrossRef]
- Vos, P.; Garrity, G.; Jones, D.; Krieg, N.R.; Ludwig, W.; Rainey, F.A.; Schleifer, K.; Whitman, W.B. (Eds.) Bergey’s Manual of Systematic Bacteriology: Volume 3: The Firmicutes; Springer Science & Business Media: Heidelberg, Germany; London, UK; New York, NY, USA, 2011; pp. 738–827. [Google Scholar]
- Gomes, M.J.P. Gênero Clostridium spp.; Favet-Ufrgs: Porto Alegre, Brazil, 2013; pp. 1–67. [Google Scholar]
- Dalgleish, T.; Williams, J.M.G.; Golden, A.-M.J.; Perkins, N.; Barrett, L.F.; Barnard, P.J.; Yeung, C.A.; Murphy, V.; Elward, R.; Tchanturia, K.; et al. Chapter 2 Prokaryotes and Their Habitats; Springer: Berlin/Heidelberg, Germany, 2007; Volume 136, ISBN 9783662131893. [Google Scholar]
- Fernández-Naveira, Á.; Abubackar, H.N.; Veiga, M.C.; Kennes, C. Efficient butanol-ethanol (BE) production from carbon monoxide fermentation by Clostridium carboxidivorans. Appl. Microbiol. Biotechnol. 2016, 100, 3361–3370. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Brown, R.; Wen, Z. Syngas fermentation of Clostridium carboxidivoran P7 in a hollow fiber membrane biofilm reactor: Evaluating the mass transfer coefficient and ethanol production performance. Biochem. Eng. J. 2014, 85, 21–29. [Google Scholar] [CrossRef]
- Sun, X.; Atiyeh, H.K.; Zhang, H.; Tanner, R.S.; Huhnke, R.L. Enhanced ethanol production from syngas by Clostridium ragsdalei in continuous stirred tank reactor using medium with poultry litter biochar. Appl. Energy 2019, 236, 1269–1279. [Google Scholar] [CrossRef]
- Atiyeh, H.K.; Lewis, R.S.; Phillips, J.R.; Huhnke, R.L. Method Improving Producer Gas Fermentation. U.S. Patent 10053711B2, 21 August 2018. [Google Scholar]
- Richter, H.; Martin, M.E.; Angenent, L.T. A two-stage continuous fermentation system for conversion of syngas into ethanol. Energies 2013, 6, 3987–4000. [Google Scholar] [CrossRef]
- Phillips, J.R.; Klasson, K.T.; Clausen, E.C.; Gaddy, J.L. Biological production of ethanol from coal synthesis gas—Medium development studies. Appl. Biochem. Biotechnol. 1993, 39–40, 559–571. [Google Scholar] [CrossRef]
- Yin, Y.; Zhang, Y.; Karakashev, D.B.; Wang, J.; Angelidaki, I. Biological caproate production by Clostridium kluyveri from ethanol and acetate as carbon sources. Bioresour. Technol. 2017, 241, 638–644. [Google Scholar] [CrossRef]
- Qureshi, N.; Cotta, M.A.; Saha, B.C. Bioconversion of barley straw and corn stover to butanol (a biofuel) in integrated fermentation and simultaneous product recovery bioreactors. Food Bioprod. Process. 2014, 92, 298–308. [Google Scholar] [CrossRef]
- Zhang, J.; Jia, B. Enhanced butanol production using Clostridium beijerinckii SE-2 from the waste of corn processing. Biomass Bioenergy 2018, 115, 260–266. [Google Scholar] [CrossRef]
- Khunchantuek, C.; Fiala, K. Optimization of key factors affecting butanol production from sugarcane juice by Clostridium beijerinckii TISTR 1461. Energy Procedia 2017, 138, 157–162. [Google Scholar] [CrossRef]
- Zhang, C.; Li, T.; He, J. Characterization and genome analysis of a butanol–isopropanol-producing Clostridium beijerinckii strain BGS1. Biotechnol. Biofuels 2018, 11, 280. [Google Scholar] [CrossRef]
- Dong, J.-J.; Ding, J.-C.; Zhang, Y.; Ma, L.; Xu, G.-C.; Han, R.-Z.; Ni, Y. Simultaneous saccharification and fermentation of dilute alkaline-pretreated corn stover for enhanced butanol production by Clostridium saccharobutylicum DSM 13864. FEMS Microbiol. Lett. 2016, 363, fnw003. [Google Scholar] [CrossRef] [PubMed]
- Darmayanti, R.F.; Tashiro, Y.; Noguchi, T.; Gao, M.; Sakai, K.; Sonomoto, K. Novel biobutanol fermentation at a large extractant volume ratio using immobilized Clostridium saccharoperbutylacetonicum N1-4. J. Biosci. Bioeng. 2018, 126, 750–757. [Google Scholar] [CrossRef] [PubMed]
- Balusu, R.; Paduru, R.R.; Kuravi, S.K.; Seenayya, G.; Reddy, G. Optimization of critical medium components using response surface methodology for ethanol production from cellulosic biomass by Clostridium thermocellum SS19. Process Biochem. 2005, 40, 3025–3030. [Google Scholar] [CrossRef]
- Jin, M.; Gunawan, C.; Balan, V.; Dale, B.E. Consolidated bioprocessing (CBP) of AFEX™-pretreated corn stover for ethanol production using Clostridium phytofermentans at a high solids loading. Biotechnol. Bioeng. 2012, 109, 1929–1936. [Google Scholar] [CrossRef]
- Lee, S.H.; Kim, S.; Kim, J.Y.; Cheong, N.Y.; Kim, K.H. Enhanced butanol fermentation using metabolically engineered Clostridium acetobutylicum with ex situ recovery of butanol. Bioresour. Technol. 2016, 218, 909–917. [Google Scholar] [CrossRef]
- Lehmann, D.; Lütke-Eversloh, T. Switching Clostridium acetobutylicum to an ethanol producer by disruption of the butyrate/butanol fermentative pathway. Metab. Eng. 2011, 13, 464–473. [Google Scholar] [CrossRef]
- Kong, X.; He, A.; Zhao, J.; Wu, H.; Jiang, M. Efficient acetone-butanol-ethanol production (ABE) by Clostridium acetobutylicum XY16 immobilized on chemically modified sugarcane bagasse. Bioprocess Biosyst. Eng. 2015, 38, 1365–1372. [Google Scholar] [CrossRef]
- Wilkens, E.; Ringel, A.K.; Hortig, D.; Willke, T.; Vorlop, K.D. High-level production of 1,3-propanediol from crude glycerol by Clostridium butyricum AKR102a. Appl. Microbiol. Biotechnol. 2012, 93, 1057–1063. [Google Scholar] [CrossRef]
- Li, Y.; Tschaplinski, T.J.; Engle, N.L.; Hamilton, C.Y.; Modriguez, R., Jr.; Liao, J.C.; Schadt, C.W.; Guss, A.M.; Yang, Y.; Graham, D.E. Combined inactivation of the Clostridium cellulolyticum lactate and malate dehydrogenase genes substantially increases ethanol yield from cellulose and switchgrass fermentations. Biotechnol. Biofuels 2012, 5, 2. [Google Scholar] [CrossRef]
- Ou, J.; Xu, N.; Ernst, P.; Ma, C.; Bush, M.; Goh, K.; Zhao, J.; Zhou, L.; Yang, S.T.; Liu, X.M. Process engineering of cellulosic n-butanol production from corn-based biomass using Clostridium cellulovorans. Process Biochem. 2017, 62, 144–150. [Google Scholar] [CrossRef]
- Malaviya, A.; Jang, Y.S.; Lee, S.Y. Continuous butanol production with reduced byproducts formation from glycerol by a hyper producing mutant of Clostridium pasteurianum. Appl. Microbiol. Biotechnol. 2012, 93, 1485–1494. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Dai, L.; Xin, B.; Tao, F.; Tang, H.; Shen, Y.; Xu, P. 1,3-Propanediol production by a newly isolated strain, Clostridium perfringens GYL. Bioresour. Technol. 2017, 233, 406–412. [Google Scholar] [CrossRef] [PubMed]
- Bengelsdorf, F.R.; Straub, M.; Dürre, P. Bacterial synthesis gas (syngas) fermentation. Environ. Technol. 2013, 34, 1639–1651. [Google Scholar] [CrossRef] [PubMed]
- Coelho, F.; Nele, M.; Ribeiro, R.; Ferreira, T.; Amaral, P. Clostridium carboxidivorans’ surface characterization using contact angle measurement (CAM). Chem. Eng. Trans. 2016, 50, 277–282. [Google Scholar]
- Bruant, G.; Lévesque, M.J.; Peter, C.; Guiot, S.R.; Masson, L. Genomic analysis of carbon monoxide utilization and butanol production by Clostridium carboxidivorans strain P7T. PLoS ONE 2010, 5, e13033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huhnke, R.; Lewis, R.; Tanner, R.S. Isolation and Characterization of Novel Clostridial Species. U.S. Patent 2008/0057554A1, 6 March 2008. [Google Scholar]
- Saxena, J.; Tanner, R.S. Effect of trace metals on ethanol production from synthesis gas by the ethanologenic acetogen, Clostridium ragsdalei. J. Ind. Microbiol. Biotechnol. 2011, 38, 513–521. [Google Scholar] [CrossRef]
- Ragsdale, S.W. Life with carbon monoxide. Crit. Rev. Biochem. Mol. Biol. 2004, 39, 165–195. [Google Scholar] [CrossRef]
- Saxena, J.; Tanner, R.S. Optimization of a corn steep medium for production of ethanol from synthesis gas fermentation by Clostridium ragsdalei. World J. Microbiol. Biotechnol. 2012, 28, 1553–1561. [Google Scholar] [CrossRef]
- Devarapalli, M.; Atiyeh, H.K.; Phillips, J.R.; Lewis, R.S.; Huhnke, R.L. Ethanol production during semi-continuous syngas fermentation in a trickle bed reactor using Clostridium ragsdalei. Bioresour. Technol. 2016, 209, 56–65. [Google Scholar] [CrossRef] [Green Version]
- Kundiyana, D.K.; Wilkins, M.R.; Maddipati, P.; Huhnke, R.L. Effect of temperature, pH and buffer presence on ethanol production from synthesis gas by “Clostridium ragsdalei”. Bioresour. Technol. 2011, 102, 5794–5799. [Google Scholar] [CrossRef]
- Tanner, R.S.; Miller, L.M.; Yang, D. Clostridium ljungdahlii sp. nov., an acetogenic species in clostridial rRNA homology group I. Int. J. Syst. Bacteriol. 1993, 43, 232–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barker, H.A.; Taha, S.M. Clostridium kluyverii, an organism concerned in the formation of caproic acid from ethyl alcohol. J. Bacteriol. 1941, 43, 347–363. [Google Scholar]
- Keis, S.; Shaheen, R.; Jones, D.T. Emended descriptions of Clostridium acetobutylicum and Clostridium beijerinckii, and descriptions of Clostridium saccharoperbutylacetonicum sp. nov. and Clostridium saccharobutylicum sp. nov. Int. J. Syst. Evol. Microbiol. 2001, 51, 2095–2103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trchounian, K.; Müller, N.; Schink, B.; Trchounian, A. Glycerol and mixture of carbon sources conversion to hydrogen by Clostridium beijerinckii DSM791 and effects of various heavy metals on hydrogenase activity. Int. J. Hydrog. Energy 2017, 42, 7875–7882. [Google Scholar] [CrossRef] [Green Version]
- Diallo, M.; Simons, A.D.; van der Wal, H.; Collas, F.; Houweling-Tan, B.; Kengen, S.W.M.; López-Contreras, A.M. L-Rhamnose metabolism in Clostridium beijerinckii strain DSM 6423. Appl. Environ. Microbiol. 2018, 85, e02656-18. [Google Scholar] [CrossRef] [Green Version]
- Ni, Y.; Xia, Z.; Wang, Y.; Sun, Z. Continuous butanol fermentation from inexpensive sugar-based feedstocks by Clostridium saccharobutylicum DSM 13864. Bioresour. Technol. 2013, 129, 680–685. [Google Scholar] [CrossRef]
- Gu, Y.; Feng, J.; Zhang, Z.T.; Wang, S.; Guo, L.; Wang, Y.; Wang, Y. Curing the endogenous megaplasmid in Clostridium saccharoperbutylacetonicum N1-4 (HMT) using CRISPR-Cas9 and preliminary investigation of the role of the plasmid for the strain metabolism. Fuel 2019, 236, 1559–1566. [Google Scholar] [CrossRef]
- Wang, S.; Dong, S.; Wang, P.; Tao, Y.; Wang, Y. Genome editing in Clostridium saccharoperbutylacetonicum N1-4 with the CRISPR-Cas9 system. Appl. Environ. Microbiol. 2017, 83, e00233-17. [Google Scholar] [CrossRef] [Green Version]
- Viljoen, J.A.; Fred, E.B.; Peterson, W.H. The fermentation of cellulose by thermophilic bacteria. J. Agric. Sci. 1926, 16, 1–17. [Google Scholar] [CrossRef]
- Lo, J.; Olson, D.G.; Murphy, S.J.; Tian, L.; Hon, S.; Lanahan, A.; Guss, A.M.; Lynd, L.R. Engineering electron metabolism to increase ethanol production in Clostridium thermocellum. Metab. Eng. 2017, 39, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Dash, S.; Olson, D.G.; Chan, S.H.J.; Amador-Noguez, D.; Lynd, L.R.; Maranas, C.D. Thermodynamic analysis of the pathway for ethanol production from cellobiose in Clostridium thermocellum. Metab. Eng. 2019, 55, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Methé, B.A.; Leschine, S.B.; Warnick, T.A. Clostridium phytofermentans sp. nov., a cellulolytic mesophile from forest soil. Int. J. Syst. Evol. Microbiol. 2002, 52, 1155–1160. [Google Scholar]
- Fathima, A.A.; Sanitha, M.; Kumar, T.; Iyappan, S.; Ramya, M. Direct utilization of waste water algal biomass for ethanol production by cellulolytic Clostridium phytofermentans DSM1183. Bioresour. Technol. 2016, 202, 253–256. [Google Scholar] [CrossRef] [PubMed]
- Algayyim, S.J.M.; Wandel, A.P.; Yusaf, T.; Hamawand, I. Production and application of ABE as a biofuel. Renew. Sustain. Energy Rev. 2018, 82, 1195–1214. [Google Scholar] [CrossRef]
- Tsvetanova, F.; Petrova, P.; Petrov, K. Microbial production of 1-butanol-recent advances and future prospects (review). J. Chem. Technol. Metall. 2018, 53, 683–696. [Google Scholar]
- Zhou, W.; Liu, J.; Fan, S.; Xiao, Z.; Qiu, B.; Wang, Y.; Li, J.; Liu, Y. Biofilm immobilization of Clostridium acetobutylicum on particulate carriers for acetone-butanol-ethanol (ABE) production. Bioresour. Technol. Rep. 2018, 3, 211–217. [Google Scholar] [CrossRef]
- Yang, Y.; Nie, X.; Jiang, Y.; Yang, C.; Gu, Y.; Jiang, W. Metabolic regulation in solventogenic clostridia: Regulators, mechanisms and engineering. Biotechnol. Adv. 2018, 36, 905–914. [Google Scholar] [CrossRef]
- Sun, C.; Zhang, S.; Xin, F.; Shanmugam, S.; Wu, Y.R. Genomic comparison of Clostridium species with the potential of utilizing red algal biomass for biobutanol production. Biotechnol. Biofuels 2018, 11, 42. [Google Scholar] [CrossRef]
- Li, C.; Wang, Y.; Xie, G.; Peng, B.; Zhang, B.; Chen, W.; Huang, X.; Wu, H.; Zhang, B. Complete genome sequence of Clostridium butyricum JKY6D1 isolated from the pit mud of a Chinese flavor liquor-making factory. J. Biotechnol. 2016, 220, 23–24. [Google Scholar] [CrossRef]
- Cardoso, C.J.T.; de Junior, J.S.O.; Kischel, H.; da Silva, W.A.L.; da Arruda, E.D.S.; Souza-Cáceres, M.B.; de Oliveira, F.A.M.; Nogueira, É.; de Nogueira, G.P.; de Melo-Sterza, F.A. Anti-Müllerian hormone (AMH) as a predictor of antral follicle population in heifers. Anim. Reprod. 2017, 15, 12–16. [Google Scholar] [CrossRef]
- Zhang, A.H.; Liu, H.L.; Huang, S.Y.; Fu, Y.S.; Fang, B.S. Metabolic profiles analysis of 1,3-propanediol production process by Clostridium butyricum through repeated batch fermentation coupled with activated carbon adsorption. Biotechnol. Bioeng. 2017, 115, 684–693. [Google Scholar] [CrossRef]
- Gaida, S.M.; Liedtke, A.; Jentges, A.H.W.; Engels, B.; Jennewein, S. Metabolic engineering of Clostridium cellulolyticum for the production of n-butanol from crystalline cellulose. Microb. Cell Fact. 2016, 15, 6. [Google Scholar] [CrossRef]
- Desvaux, M. Clostridium cellulolyticum: Model organism of mesophilic cellulolytic clostridia. FEMS Microbiol. Rev. 2005, 29, 741–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomita, H.; Okazaki, F.; Tamaru, Y. Direct IBE fermentation from mandarin orange wastes by combination of Clostridium cellulovorans and Clostridium beijerinckii. AMB Express 2019, 9, 1. [Google Scholar] [CrossRef] [PubMed]
- Xin, F.; Dong, W.; Zhang, W.; Ma, J.; Jiang, M. Biobutanol production from crystalline cellulose through consolidated bioprocessing. Trends Biotechnol. 2019, 37, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Groeger, C.; Wang, W.; Sabra, W.; Utesch, T.; Zeng, A.P. Metabolic and proteomic analyses of product selectivity and redox regulation in Clostridium pasteurianum grown on glycerol under varied iron availability. Microb. Cell Fact. 2017, 16, 64. [Google Scholar] [CrossRef]
- Ahn, J.-H.; Sang, B.-I.; Um, Y. Butanol production from thin stillage using Clostridium pasteurianum. Bioresour. Technol. 2011, 102, 4934–4937. [Google Scholar] [CrossRef]
- López-Enríquez, L.; Rodríguez-Lázaro, D.; Hernández, M. Quantitative detection of Clostridium tyrobutyricum in milk by real-time PCR. Appl. Environ. Microbiol. 2007, 73, 3747–3751. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Ai, H.; Zhang, S.; Li, S.; Liang, Z.; Wu, Z.-Q.; Yang, S.-T.; Wang, J.-F. Enhanced butanol production by coculture of Clostridium beijerinckii and Clostridium tyrobutyricum. Bioresour. Technol. 2013, 143, 397–404. [Google Scholar] [CrossRef]
- Abrini, J.; Naveau, H.; Nyns, E.-J. Clostridium autoethanogenumt, sp. nov., an anaerobic bacterium that produces ethanol from carbon monoxide. Arch. Microbiol. 1994, 161, 345–351. [Google Scholar] [CrossRef]
- Valgepea, K.; de Souza Pinto Lemgruber, R.; Abdalla, T.; Binos, S.; Takemori, N.; Takemori, A.; Tanaka, Y.; Tappel, R.; Köpke, M.; Simpson, S.D.; et al. H2 drives metabolic rearrangements in gas-fermenting Clostridium autoethanogenum. Biotechnol. Biofuels 2018, 11, 55. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Naveira, Á.; Abubackar, H.N.; Veiga, M.C.; Kennes, C. Production of chemicals from C1 gases (CO, CO2) by Clostridium carboxidivorans. World J. Microbiol. Biotechnol. 2017, 33, 43. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Naveira, Á.; Veiga, M.C.; Kennes, C. H-B-E (hexanol-butanol-ethanol) fermentation for the production of higher alcohols from syngas/waste gas. J. Chem. Technol. Biotechnol. 2017, 92, 712–731. [Google Scholar] [CrossRef]
- Jones, D.T.; Woods, D.R. Acetone-butanol fermentation revisited. Microbiol. Rev. 1986, 50, 484–524. [Google Scholar]
- Moon, H.G.; Jang, Y.-S.; Cho, C.; Lee, J.; Binkley, R.; Lee, S.Y. One hundred years of clostridial butanol fermentation. FEMS Microbiol. Lett. 2016, 363, fnw001. [Google Scholar] [CrossRef] [Green Version]
- Noguchi, T.; Tashiro, Y.; Yoshida, T.; Zheng, J.; Sakai, K.; Sonomoto, K. Efficient butanol production without carbon catabolite repression from mixed sugars with Clostridium saccharoperbutylacetonicum N1-4. J. Biosci. Bioeng. 2013, 116, 716–721. [Google Scholar] [CrossRef]
- Fast, A.G.; Schmidt, E.D.; Jones, S.W.; Tracy, B.P. Acetogenic mixotrophy: NOVEL options for yield improvement in biofuels and biochemicals production. Curr. Opin. Biotechnol. 2015, 33, 60–72. [Google Scholar] [CrossRef]
- Sun, X.; Atiyeh, H.K.; Adesanya, Y.A.; Zhang, H.; Okonkwo, C.; Ezeji, T. Enhanced acetone-butanol-ethanol production by Clostridium beijerinckii using biochar. In Proceedings of the 2019 ASABE Annual International Meeting, Boston, MA, USA, 7–10 July 2019; p. 1. [Google Scholar]
- dos Santos Vieira, C.F.; Filho, F.M.; Filho, R.M.; Mariano, A.P. Acetone-free biobutanol production: Past and recent advances in the Isopropanol-Butanol-Ethanol (IBE) fermentation. Bioresour. Technol. 2019, 287, 121425. [Google Scholar] [CrossRef]
- Li, G.; Lee, T.H.; Liu, Z.; Lee, C.F.; Zhang, C. Effects of injection strategies on combustion and emission characteristics of a common-rail diesel engine fueled with isopropanol-butanol-ethanol and diesel blends. Renew. Energy 2019, 130, 677–686. [Google Scholar] [CrossRef]
- Bengelsdorf, F.R.; Dürre, P. Gas fermentation for commodity chemicals and fuels. Microb. Biotechnol. 2017, 10, 1167–1170. [Google Scholar] [CrossRef]
- Raganati, F.; Procentese, A.; Olivieri, G.; Götz, P.; Salantino, P.; Marzocchella, A. Kinetic study of butanol production from various sugars by Clostridium acetobutylicum using a dynamic model. Biochem. Eng. 2015, 99, 156–166. [Google Scholar] [CrossRef]
- Shinto, H.; Tashiro, Y.; Yamashita, M.; Kobayashi, G.; Sekiguchi, T.; Hanai, T. Kinetic study of substrate dependency for higher butanol production in acetone–butanol–ethanol fermentation. Process Biochem. 2008, 43, 1452–1461. [Google Scholar] [CrossRef]
- Yu, Y.; Tangney, M.; Aass, H.C.; Mitchell, W.J. Analysis of the mechanism and regulation of lactose transport and metabolism in Clostridium acetobutylicum ATCC 824. Appl. Environ. Microbiol. 2007, 73, 1842–1850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abubackar, H.N.; Veiga, M.C.; Kennes, C. Carbon monoxide fermentation to ethanol by Clostridium autoethanogenum in a bioreactor with no accumulation of acetic acid. Bioresour. Technol. 2015, 186, 122–127. [Google Scholar] [CrossRef] [Green Version]
- Ciriminna, R.; Pina, C.D.; Rossi, M.; Pagliaro, M. Understanding the glycerol market. Eur. J. Lipid Sci. Technol. 2014, 116, 1432–1439. [Google Scholar] [CrossRef]
- Pagliaro, M.; Rossi, M. Chapter 1. Glycerol: Properties and production. In The Future of Glycerol, 2nd ed.; Royal Society of Chemistry (RSC) Publishing: Cambridge, UK, 2010; pp. 1–28. [Google Scholar]
- Ayoub, M.; Abdullah, A.Z. Critical review on the current scenario and significance of crude glycerol resulting from biodiesel industry towards more sustainable renewable energy industry. Renew. Sustain. Energy Rev. 2012, 16, 2671–2686. [Google Scholar] [CrossRef]
- Amaral, P.F.F.; Ferreira, T.F.; Fontes, G.C.; Coelho, M.A.Z. Glycerol valorization: New biotechnological routes. Food Bioprod. Process. 2009, 87, 179–186. [Google Scholar] [CrossRef]
- Pradima, J.; Kulkarni, M.R. Review on enzymatic synthesis of value added products of glycerol, a by-product derived from biodiesel production. Resour. Technol. 2016, 3, 394–405. [Google Scholar] [CrossRef]
- Sanguanchaipaiwong, V.; Leksawasdi, N. Using glycerol as a sole carbon source for Clostridium beijerinckii fermentation. Energy Procedia 2017, 138, 1105–1109. [Google Scholar] [CrossRef]
- da Silva, G.P.; Mack, M.; Contiero, J. Glycerol: A promising and abundant carbon source for industrial microbiology. Biotechnol. Adv. 2008, 27, 30–39. [Google Scholar] [CrossRef]
- Forsberg, C.W. Production of 1,3-propanediol from glycerol by Clostridium acetobutylicum and other Clostridium species. Appl. Environ. Microbiol. 1987, 53, 639–643. [Google Scholar] [PubMed]
- Tee, Z.K.; Jahim, J.M.; Tan, J.P.; Kim, B.H. Preeminent productivity of 1,3-propanediol by Clostridium butyricum JKT37 and the role of using calcium carbonate as pH neutraliser in glycerol fermentation. Bioresour. Technol. 2017, 233, 296–304. [Google Scholar] [CrossRef] [PubMed]
- García, C.A.; García-Treviño, E.S.; Aguilar-Rivera, N.; Armendáriz, C. Carbon footprint of sugar production in Mexico. J. Clean. Prod. 2016, 112, 2632–2641. [Google Scholar] [CrossRef]
- de Oliveira Lino, F.S.; Basso, T.O.; Sommer, M.O.A. A synthetic medium to simulate sugarcane molasses. Biotechnol. Biofuels 2018, 11, 221. [Google Scholar] [CrossRef] [PubMed]
- Li, H.G.; Luo, W.; Gu, Q.Y.; Wang, Q.; Hu, W.J.; Yu, X.B. Acetone, butanol, and ethanol production from cane molasses using Clostridium beijerinckii mutant obtained by combined low-energy ion beam implantation and N-methyl-N-nitro-N-nitrosoguanidine induction. Bioresour. Technol. 2013, 137, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Farooq, U.; Anjum, F.; Zahoor, T.; Rahman, S.; Randhawa, M.; Ahmed, A.; Akram, K. Optimization of lactic acid production from cheap raw material: Sugarcane molasses. Pak. J. Bot. 2012, 44, 333. [Google Scholar]
- Syed, Q.; Nadeem, M.; Nelofer, R. Enhanced butanol production by mutant strains of Clostridium acetobutylicum in molasses medium. Turk. J. Biochem. 2008, 33, 25–30. [Google Scholar]
- van der Merwe, A.B.; Cheng, H.; Görgens, J.F.; Knoetze, J.H. Comparison of energy efficiency and economics of process designs for biobutanol production from sugarcane molasses. Fuel 2013, 105, 451–458. [Google Scholar] [CrossRef]
- Jiang, L.; Wang, J.; Liang, S.; Wang, X.; Cen, P.; Xu, Z. Butyric acid fermentation in a fibrous bed bioreactor with immobilized Clostridium tyrobutyricum from cane molasses. Bioresour. Technol. 2009, 100, 3403–3409. [Google Scholar] [CrossRef]
- Valli, V.; Gómez-Caravaca, A.M.; di Nunzio, M.; Danesi, F.; Caboni, M.F.; Bordoni, A. Sugar cane and sugar beet molasses, antioxidant-rich alternatives to refined sugar. J. Agric. Food Chem. 2012, 60, 12508–12515. [Google Scholar] [CrossRef]
- Guan, Y.; Tang, Q.; Fu, X.; Yu, S.; Wu, S.; Chen, M. Preparation of antioxidants from sugarcane molasses. Food Chem. 2014, 152, 552–557. [Google Scholar] [CrossRef] [PubMed]
- de Azeredo, L.A.I.; de Lima, M.B.; Coelho, R.R.R.; Freire, D.M.G. A low-cost fermentation medium for thermophilic protease production by Streptomyces sp. 594 using feather meal and corn steep liquor. Curr. Microbiol. 2006, 53, 335. [Google Scholar] [CrossRef] [PubMed]
- Maddipati, P.; Atiyeh, H.K.; Bellmer, D.D.; Huhnke, R.L. Ethanol production from syngas by Clostridium strain P11 using corn steep liquor as a nutrient replacement to yeast extract. Bioresour. Technol. 2011, 102, 6494–6501. [Google Scholar] [CrossRef] [PubMed]
- Ramachandriya, K.D.; Kundiyana, D.K.; Wilkins, M.R.; Terrill, J.B.; Atiyeh, H.K.; Huhnke, R.L. Carbon dioxide conversion to fuels and chemicals using a hybrid green process. Appl. Energy 2013, 112, 289–299. [Google Scholar] [CrossRef]
- Saha, B.C. Hemicellulose bioconversion. J. Ind. Microbiol. Biotechnol. 2003, 30, 279–291. [Google Scholar] [CrossRef]
- Isikgor, F.H.; Becer, C.R. Lignocellulosic biomass: A sustainable platform for the production of bio-based chemicals and polymers. Polym. Chem. 2015, 6, 4497–4559. [Google Scholar] [CrossRef] [Green Version]
- Tilman, D.; Hill, J.; Lehman, C. Supportin online material for: Carbon-negative biofuels from low-input high-diversity grassland biomass. Science 2006, 314, 1598–1600. [Google Scholar] [CrossRef] [Green Version]
- Ahn, Y.; Lee, S.H.; Kim, H.J.; Yang, Y.H.; Hong, J.H.; Kim, Y.H.; Kim, H. Electrospinning of lignocellulosic biomass using ionic liquid. Carbohydr. Polym. 2012, 88, 395–398. [Google Scholar] [CrossRef]
- Menon, V.; Rao, M. Trends in bioconversion of lignocellulose: Biofuels, platform chemicals & biorefinery concept. Prog. Energy Combust. Sci. 2012, 38, 522–550. [Google Scholar]
- Nielsen, J.; Villadsen, J.; Lidén, G. Principles, Bioreaction Engineering; Springer: Berlin, Germany, 2014; ISBN 9781441996879. [Google Scholar]
- Fengel, D.; Wegener, G. Wood—Chemistry, Ultrastructure, Reactions; Walter de Gruyter: Berlin, Germany, 1989; ISBN 9783935638395. [Google Scholar]
- Hendriks, A.T.W.M.; Zeeman, G. Pretreatments to enhance the digestibility of lignocellulosic biomass. Bioresour. Technol. 2009, 100, 10–18. [Google Scholar] [CrossRef]
- Li, F.; Cheng, S.; Yu, H.; Yang, D. Waste from livestock and poultry breeding and its potential assessment of biogas energy in rural China. J. Clean. Prod. 2016, 126, 451–460. [Google Scholar] [CrossRef]
- Ruiz, H.A.; Rodríguez-Jasso, R.M.; Fernandes, B.D.; Vicente, A.A.; Teixeira, J.A. Hydrothermal processing, as an alternative for upgrading agriculture residues and marine biomass according to the biorefinery concept: A review. Renew. Sustain. Energy Rev. 2013, 21, 35–51. [Google Scholar] [CrossRef] [Green Version]
- Amiri, H.; Karimi, K. Pretreatment and hydrolysis of lignocellulosic wastes for butanol production: Challenges and perspectives. Bioresour. Technol. 2018, 270, 702–721. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, N.; Ezeji, T.C.; Ebener, J.; Dien, B.S.; Cotta, M.A.; Blaschek, H.P. Butanol production by Clostridium beijerinckii. Part I: Use of acid and enzyme hydrolyzed corn fiber. Bioresour. Technol. 2008, 99, 5915–5922. [Google Scholar] [CrossRef]
- Bellido, C.; Pinto, M.L.; Coca, M.; González-Benito, G.; García-Cubero, M.T. Acetone–butanol–ethanol (ABE) production by Clostridium beijerinckii from wheat straw hydrolysates: Efficient use of penta and hexa carbohydrates. Bioresour. Technol. 2014, 167, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Liu, S. Production of n-butanol from concentrated sugar maple hemicellulosic hydrolysate by Clostridia acetobutylicum ATCC824. Biomass Bioenergy 2012, 39, 39–47. [Google Scholar] [CrossRef]
- Ruiz-Dueñas, F.J.; Martínez, Á.T. Microbial degradation of lignin: How a bulky recalcitrant polymer is efficiently recycled in nature and how we can take advantage of this. Microb. Biotechnol. 2009, 2, 164–177. [Google Scholar] [CrossRef] [Green Version]
- Orgill, J.J.; Atiyeh, H.K.; Devarapalli, M.; Phillips, J.R.; Lewis, R.S.; Huhnke, R.L. A comparison of mass transfer coefficients between trickle-bed, Hollow fiber membrane and stirred tank reactors. Bioresour. Technol. 2013, 133, 340–346. [Google Scholar] [CrossRef]
- Mohammadi, M.; Najafpour, G.D.; Younesi, H.; Lahijani, P.; Uzir, M.H.; Mohamed, A.R. Bioconversion of synthesis gas to second generation biofuels: A review. Renew. Sustain. Energy Rev. 2011, 15, 4255–4273. [Google Scholar] [CrossRef]
- Henstra, A.M.; Sipma, J.; Rinzema, A.; Stams, A.J. Microbiology of synthesis gas fermentation for biofuel production. Curr. Opin. Biotechnol. 2007, 18, 200–206. [Google Scholar] [CrossRef]
- Huber, G.W.; Iborra, S.; Corma, A. Synthesis of transportation fuels from biomass: Chemistry, catalysts, and engineering. Chem. Rev. 2006, 106, 4044–4098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woolcock, P.J.; Brown, R.C. A review of cleaning technologies for biomass-derived syngas. Biomass Bioenergy 2013, 52, 54–84. [Google Scholar] [CrossRef]
- Dry, M.E. The fischer-tropsch process: 1950–2000. Catal. Today 2002, 71, 227–241. [Google Scholar] [CrossRef]
- Phillips, J.R.; Clausen, E.C.; Gaddy, J.L. Synthesis gas as substrate for the biological production of fuels and chemicals. Appl. Biochem. Biotechnol. 1994, 45–46, 145–157. [Google Scholar] [CrossRef]
- Vega, J.L.; Clausen, E.C.; Gaddy, J.L. Design of bioreactors for coal synthesis gas fermentations. Resour. Conserv. Recycl. 1990, 3, 149–160. [Google Scholar] [CrossRef]
- Worden, R.M.; Grethlein, A.J.; Jain, M.K.; Datta, R. Production of butanol and ethanol from synthesis gas via fermentation. Fuel 1991, 70, 615–619. [Google Scholar] [CrossRef]
- Brown, R.C. Hybrid thermochemical/biological processing: Putting the cart before the horse? Appl. Biochem. Biotechnol. 2007, 137–140, 947–956. [Google Scholar] [CrossRef]
- Yasin, M.; Jeong, Y.; Park, S.; Jeong, J.; Lee, E.Y.; Lovitt, R.W.; Kim, B.H.; Lee, J.; Chang, I.S. Microbial synthesis gas utilization and ways to resolve kinetic and mass-transfer limitations. Bioresour. Technol. 2015, 177, 361–374. [Google Scholar] [CrossRef]
- Munasinghe, P.C.; Khanal, S.K. Syngas fermentation to biofuel: Evaluation of carbon monoxide mass transfer and analytical modeling using a composite hollow fiber (CHF) membrane bioreactor. Bioresour. Technol. 2012, 122, 130–136. [Google Scholar] [CrossRef]
- Bredwell, M.D.; Worden, R.M. Mass-transfer properties of microbubbles. 1. Experimental studies. Biotechnol. Prog. 1998, 14, 31–38. [Google Scholar] [CrossRef]
- Bredwell, M.D.; Srivastava, P.; Worden, R.M. Reactor design issues for synthesis-gas fermentations. Biotechnol. Prog. 1999, 15, 834–844. [Google Scholar] [CrossRef] [PubMed]
- Worden, R.M.; Bredwell, M.D. Mass-transfer properties of microbubbles. 2. Analysis using a dynamic model. Biotechnol. Prog. 1998, 14, 39–46. [Google Scholar] [CrossRef]
- Klasson, K.T.; Ackerson, M.D.; Clausen, E.C.; Gaddy, J.L. Bioreactor design for synthesis gas fermentations. Fuel 1991, 70, 605–614. [Google Scholar] [CrossRef]
- Christi, M.Y. Airlift Bioreactors; Elsevier Science Publishing Co.: New York, NY, USA, 1989; ISBN 1851663207. [Google Scholar]
- Kadic, E.; Heindel, T.J. An Introduction to Bioreactor Hydrodynamics and Gas-Liquid Mass Transfer; Wiley: Hoboken, NJ, USA, 2017; ISBN 9781118104019. [Google Scholar]
- Kumaresan, T.; Nere, N.K.; Joshi, J.B. Effect of internals on the flow pattern and mixing in stirred tanks. Ind. Eng. Chem. Res. 2005, 44, 9951–9961. [Google Scholar] [CrossRef]
- Munasinghe, P.C.; Khanal, S.K. Evaluation of hydrogen and carbon monoxide mass transfer and a correlation between the myoglobin-protein bioassay and gas chromatography method for carbon monoxide determination. RSC Adv. 2014, 4, 37575–37581. [Google Scholar] [CrossRef]
- Ungerman, A.J.; Heindel, T.J. Carbon monoxide mass transfer for syngas fermentation in a stirred tank reactor with dual impeller configurations. Biotechnol. Prog. 2007, 23, 613–620. [Google Scholar] [CrossRef]
- Yasin, M.; Park, S.; Jeong, Y.; Lee, E.Y.; Lee, J.; Chang, I.S. Effect of internal pressure and gas/liquid interface area on the CO mass transfer coefficient using hollow fibre membranes as a high mass transfer gas diffusing system for microbial syngas fermentation. Bioresour. Technol. 2014, 169, 637–643. [Google Scholar] [CrossRef]
- Coelho, F.M.B.; Botelho, A.M.; Ivo, O.F.; Amaral, P.F.F.; Ferreira, T.F. Volumetric mass transfer coefficient for carbon monoxide in a dual impeller stirred tank reactor considering a perfluorocarbon–water mixture as liquid phase. Chem. Eng. Res. Des. 2019, 143, 160–169. [Google Scholar] [CrossRef]
- Orgill, J.J.; Abboud, M.C.; Atiyeh, H.K.; Devarapalli, M.; Sun, X.; Lewis, R.S. Measurement and prediction of mass transfer coefficients for syngas constituents in a hollow fiber reactor. Bioresour. Technol. 2019, 276, 1–7. [Google Scholar] [CrossRef]
- Molitor, B.; Richter, H.; Martin, M.E.; Jensen, R.O.; Juminaga, A.; Mihalcea, C.; Angenent, L.T. Carbon recovery by fermentation of CO-rich off gases—Turning steel mills into biorefineries. Bioresour. Technol. 2016, 215, 386–396. [Google Scholar] [CrossRef]
- Chang, I.S.; Kim, B.H.; Lovitt, R.W.; Bang, J.S. Effect of CO partial pressure on cell-recycled continuous CO fermentation by Eubacterium limosum KIST612. Process Biochem. 2001, 37, 411–421. [Google Scholar] [CrossRef]
- Klasson, K.T.; Gupta, A.; Clausen, E.C.; Gaddy, J.L. Evaluation of mass-transfer and kinetic parameters for Rhodospirillum rubrum in a continuous stirred tank reactor. Appl. Biochem. Biotechnol. 1993, 39–40, 549–557. [Google Scholar] [CrossRef]
- Cowger, J.P.; Klasson, K.T.; Ackerson, M.D.; Clausen, E.; Caddy, J.L. Mass-transfer and kinetic aspects in continuous bioreactors using Rhodospirillum rubrum. Appl. Biochem. Biotechnol. 1992, 34–35, 613–624. [Google Scholar] [CrossRef]
- Lee, P.H.; Ni, S.Q.; Chang, S.Y.; Sung, S.; Kim, S.H. Enhancement of carbon monoxide mass transfer using an innovative external hollow fiber membrane (HFM) diffuser for syngas fermentation: Experimental studies and model development. Chem. Eng. J. 2012, 184, 268–277. [Google Scholar] [CrossRef]
- Riggs, S.S.; Heindel, T.J. Measuring carbon monoxide gas-liquid mass transfer in a stirred tank reactor for syngas fermentation. Biotechnol. Prog. 2006, 22, 903–906. [Google Scholar] [CrossRef] [PubMed]
- Kapic, A.; Jones, S.T.; Heindel, T.J. Carbon monoxide mass transfer in a syngas mixture. Ind. Eng. Chem. Res. 2006, 45, 9150–9155. [Google Scholar] [CrossRef] [Green Version]
- International Energy Agency (IEA). World Energy Outlook; IEA: Paris, France, 2018. [Google Scholar]
- International Energy Agency (IEA). Biofuels for Transport: Tracking Clean Energy Progress; IEA: Paris, France, 2018. [Google Scholar]
- Sun, Y.; Cheng, J. Hydrolysis of lignocellulosic materials for ethanol production: A review. Bioresour. Technol. 2002, 83, 1–11. [Google Scholar] [CrossRef]
- Cheng, J.J.; Timilsina, G.R. Status and barriers of advanced biofuel technologies: A review. Renew. Energy 2011, 36, 3541–3549. [Google Scholar] [CrossRef]
- Gallo, J.M.R.; Bueno, J.M.C.; Schuchardt, U. Catalytic transformations of ethanol for biorefineries. J. Braz. Chem. Soc. 2014, 25, 2229–2243. [Google Scholar] [CrossRef]
- Roozbehani, B.; Mirdrikvand, M.; Imani, S.; Roshan, A.C. Synthetic ethanol production in the Middle East: A way to make environmentally friendly fuels. Chem. Technol. Fuels Oils 2013, 49, 115–124. [Google Scholar] [CrossRef]
- Kundiyana, D.K.; Huhnke, R.L.; Wilkins, M.R. Syngas fermentation in a 100-L pilot scale fermentor: Design and process considerations. J. Biosci. Bioeng. 2010, 109, 492–498. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.E.; Richter, H.; Saha, S.; Angenent, L.T. Traits of selected Clostridium strains for syngas fermentation to ethanol. Biotechnol. Bioeng. 2016, 113, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Gu, Y.; Chai, C.; Jiang, W.; Zhuang, Y.; Wang, Y. Enhanced alcohol titre and ratio in carbon monoxide-rich off-gas fermentation of Clostridium carboxidivorans through combination of trace metals optimization with variable-temperature cultivation. Bioresour. Technol. 2017, 239, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.K.; Lee, H. Use of magnetic nanoparticles to enhance bioethanol production in syngas fermentation. Bioresour. Technol. 2016, 204, 139–144. [Google Scholar] [CrossRef]
- Li, D.; Meng, C.; Wu, G.; Xie, B.; Han, Y.; Guo, Y.; Song, C.; Gao, Z.; Huang, Z. Effects of zinc on the production of alcohol by Clostridium carboxidivorans P7 using model syngas. J. Ind. Microbiol. Biotechnol. 2018, 45, 61–69. [Google Scholar] [CrossRef]
- Abubackar, H.N.; Fernández-naveira, Á.; Veiga, M.C.; Kennes, C. Impact of cyclic pH shifts on carbon monoxide fermentation to ethanol by Clostridium autoethanogenum. Fuel 2016, 178, 56–62. [Google Scholar] [CrossRef]
- Wang, S.; Dong, S.; Wang, Y. Enhancement of solvent production by overexpressing key genes of the acetone-butanol-ethanol fermentation pathway in Clostridium saccharoperbutylacetonicum N1-4. Bioresour. Technol. 2017, 245, 426–433. [Google Scholar] [CrossRef]
- Fernández-Naveira, Á.; Veiga, M.C.; Kennes, C. Glucose bioconversion profile in the syngas-metabolizing species Clostridium carboxidivorans. Bioresour. Technol. 2017, 244, 552–559. [Google Scholar] [CrossRef]
- Singh, N.; Mathur, A.S.; Gupta, R.P.; Barrow, C.J.; Tuli, D.; Puri, M. Enhanced cellulosic ethanol production via consolidated bioprocessing by Clostridium thermocellum ATCC 31924. Bioresour. Technol. 2018, 250, 860–867. [Google Scholar] [CrossRef]
- Yang, X.; Xu, M.; Yang, S.T. Metabolic and process engineering of Clostridium cellulovorans for biofuel production from cellulose. Metab. Eng. 2015, 32, 39–48. [Google Scholar] [CrossRef]
- Khanna, S.; Goyal, A.; Moholkar, V.S. Effect of fermentation parameters on bio-alcohols production from glycerol using immobilized Clostridium pasteurianum: An optimization study. Prep. Biochem. Biotechnol. 2013, 43, 828–847. [Google Scholar] [CrossRef] [PubMed]
- Vohra, M.; Manwar, J.; Manmode, R.; Padgilwar, S.; Patil, S. Bioethanol production: Feedstock and current technologies. J. Environ. Chem. Eng. 2014, 2, 573–584. [Google Scholar] [CrossRef]
- Devarapalli, M.; Atiyeh, H.K. A review of conversion processes for bioethanol production with a focus on syngas fermentation. Biofuel Res. J. 2015, 2, 268–280. [Google Scholar] [CrossRef]
- Lynd, L.R.; Weimer, P.J.; van Zyl, W.H.; Pretorius, I.S. Microbial cellulose utilization: Fundamentals and biotechnology. Microbiol. Mol. Biol. Rev. 2003, 66, 506–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margeot, A.; Hahn-Hagerdal, B.; Edlund, M.; Slade, R.; Monot, F. New improvements for lignocellulosic ethanol. Curr. Opin. Biotechnol. 2009, 20, 372–380. [Google Scholar] [CrossRef]
- Lynd, L.R.; van Zyl, W.H.; McBride, J.E.; Laser, M. Consolidated bioprocessing of cellulosic biomass: An update. Curr. Opin. Biotechnol. 2005, 16, 577–583. [Google Scholar] [CrossRef]
- Sasaki, C.; Kushiki, Y.; Asada, C.; Nakamura, Y. Acetone-butanol-ethanol production by separate hydrolysis and fermentation (SHF) and simultaneous saccharification and fermentation (SSF) methods using acorns and wood chips of Quercus acutissima as a carbon source. Ind. Crop. Prod. 2014, 62, 286–292. [Google Scholar] [CrossRef]
- Shen, Y.; Brown, R.C.; Wen, Z. Syngas fermentation by Clostridium carboxidivorans P7 in a horizontal rotating packed bed biofilm reactor with enhanced ethanol production. Appl. Energy 2017, 187, 585–594. [Google Scholar] [CrossRef]
- Qureshi, N.; Blaschek, H.P. ABE production from corn: A recent economic evaluation. J. Ind. Microbiol. Biotechnol. 2001, 27, 292–297. [Google Scholar] [CrossRef]
- Lee, S.Y.; Park, J.H.; Jang, S.H.; Nielsen, L.K.; Kim, J.; Jung, K.S. Fermentative butanol production by clostridia. Biotechnol. Bioeng. 2008, 101, 209–228. [Google Scholar] [CrossRef]
- Jin, C.; Yao, M.; Liu, H.; Lee, C.F.F.; Ji, J. Progress in the production and application of n-butanol as a biofuel. Renew. Sustain. Energy Rev. 2011, 15, 4080–4106. [Google Scholar] [CrossRef]
- Zheng, J.; Tashiro, Y.; Wang, Q.; Sonomoto, K. Recent advances to improve fermentative butanol production: Genetic engineering and fermentation technology. J. Biosci. Bioeng. 2015, 119, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Nicolaou, S.A.; Gaida, S.M.; Papoutsakis, E.T. A comparative view of metabolite and substrate stress and tolerance in microbial bioprocessing: From biofuels and chemicals, to biocatalysis and bioremediation. Metab. Eng. 2010, 12, 307–331. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, R.; Alves, M.; Rodrigues, L.R. Modulation of crude glycerol fermentation by Clostridium pasteurianum DSM 525 towards the production of butanol. Biomass Bioenergy 2014, 71, 134–143. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.; Zhao, J.; Yang, S.T.; Wei, D. Fed-batch fermentation for n-butanol production from cassava bagasse hydrolysate in a fibrous bed bioreactor with continuous gas stripping. Bioresour. Technol. 2012, 104, 380–387. [Google Scholar] [CrossRef]
- Phillips, J.R.; Atiyeh, H.K.; Tanner, R.S.; Torres, J.R.; Saxena, J.; Wilkins, M.R.; Huhnke, R.L. Butanol and hexanol production in Clostridium carboxidivorans syngas fermentation: Medium development and culture techniques. Bioresour. Technol. 2015, 190, 114–121. [Google Scholar] [CrossRef]
- Zhang, J.; Yu, L.; Lin, M.; Yan, Q.; Yang, S.-T. n-Butanol production from sucrose and sugarcane juice by engineered Clostridium tyrobutyricum overexpressing sucrose catabolism genes and adhE2. Bioresour. Technol. 2017, 233, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Maiti, S.; Gallastegui, G.; Sarma, S.J.; Brar, S.K.; le Bihan, Y.; Drogui, P.; Buelna, G.; Verma, M. A re-look at the biochemical strategies to enhance butanol production. Biomass Bioenergy 2016, 94, 187–200. [Google Scholar] [CrossRef]
- Ahlawat, S.; Kaushal, M.; Palabhanvi, B.; Muthuraj, M.; Goswami, G.; Das, D. Nutrient modulation based process engineering strategy for improved butanol production from Clostridium acetobutylicum. Biotechnol. Prog. 2019, 35, e2771. [Google Scholar] [CrossRef]
- Dong, Z.; Liu, G.; Liu, S.; Liu, Z.; Jin, W. High performance ceramic hollow fiber supported PDMS composite pervaporation membrane for bio-butanol recovery. J. Membr. Sci. 2014, 450, 38–47. [Google Scholar] [CrossRef]
- Raganati, F.; Procentese, A.; Olivieri, G.; Russo, M.E.; Salatino, P.; Marzocchella, A. Bio-butanol separation by adsorption on various materials: Assessment of isotherms and effects of other ABE-fermentation compounds. Sep. Purif. Technol. 2018, 191, 328–339. [Google Scholar] [CrossRef]
- Research&Market. Acetone: Worldwide Market Outlook to 2023—Expected to Reach ~7666 Thousand Tons by the End of 2023. 2018. Available online: https://www.prnewswire.com/news-releases/acetone-worldwide-market-outlook-to-2023---expected-to-reach-7-666-thousand-tons-by-the-end-of-2023--300757524.html (accessed on 6 June 2019).
- Sauer, M. Industrial production of acetone and butanol by fermentation—100 years later. FEMS Microbiol. Lett. 2016, 363, fnw134. [Google Scholar] [CrossRef]
- Sreekumar, S.; Baer, Z.C.; Pazhamalai, A.; Gunbas, G.; Grippo, A.; Blanch, H.W.; Clark, D.S.; Toste, F.D. Production of an acetone-butanol-ethanol mixture from Clostridium acetobutylicum and its conversion to high-value biofuels. Nat. Protoc. 2015, 10, 528–537. [Google Scholar] [CrossRef]
- HIS Markit. Acetone. Chemical Economics Handbook. Available online: https://ihsmarkit.com/products/acetone-chemical-economics-handbook.html (accessed on 9 June 2019).
- Luo, H.; Ge, L.; Zhang, J.; Ding, J.; Chen, R.; Shi, Z. Enhancing acetone biosynthesis and acetone-butanol-ethanol fermentation performance by co-culturing Clostridium acetobutylicum/Saccharomyces cerevisiae integrated with exogenous acetate addition. Bioresour. Technol. 2016, 200, 111–120. [Google Scholar] [CrossRef]
- Wen, Z.; Ledesma-Amaro, R.; Lin, J.; Jiang, Y.; Yang, S. Improved n-butanol production from Clostridium cellulovorans by integrated metabolic and evolutionary engineering. Appl. Environ. Microbiol. 2019, 85, e02560-18. [Google Scholar] [CrossRef] [Green Version]
- Millat, T.; Winzer, K. Mathematical modelling of clostridial acetone-butanol-ethanol fermentation. Appl. Microbiol. Biotechnol. 2017, 101, 2251–2271. [Google Scholar] [CrossRef]
- Li, X.; Shi, Z.; Li, Z. Increasing butanol/acetone ratio and solvent productivity in ABE fermentation by consecutively feeding butyrate to weaken metabolic strength of butyrate loop. Bioprocess Biosyst. Eng. 2014, 37, 1609–1616. [Google Scholar] [CrossRef]
- Gao, M.; Tashiro, Y.; Yoshida, T.; Zheng, J.; Wang, Q.; Sakai, K.; Sonomoto, K. Metabolic analysis of butanol production from acetate in Clostridium saccharoperbutylacetonicum N1-4 using 13C tracer experiments. RSC Adv. 2015, 5, 8486–8495. [Google Scholar] [CrossRef]
- Boonsombuti, A.; Komolpis, K.; Luengnaruemitchai, A.; Wongkasemjit, S. Enhancement of ABE fermentation through regulation of ammonium acetate and D-xylose uptake from acid-pretreated corncobs. Ann. Microbiol. 2014, 64, 431–439. [Google Scholar] [CrossRef]
- Survase, S.A.; van Heiningen, A.; Granström, T. Continuous bio-catalytic conversion of sugar mixture to acetone-butanol-ethanol by immobilized Clostridium acetobutylicum DSM 792. Appl. Microbiol. Biotechnol. 2012, 93, 2309–2316. [Google Scholar] [CrossRef]
- Tjahjasari, D.; Kaeding, T.; Zeng, A.P. 1,3-Propanediol and polytrimethyleneterephthalate. In Comprehensive Biotechnology. Volume 3: Ind Biotechnol Commodity Products; Elsevier: Pergamon, Turkey, 2011; pp. 229–242. [Google Scholar]
- Selembo, P.A.; Perez, J.M.; Lloyd, W.A.; Logan, B.E. Enhanced hydrogen and 1,3-propanediol production from glycerol by fermentation using mixed cultures. Biotechnol. Bioeng. 2009, 104, 1098–1106. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Ma, C.; Fu, H.; Mu, Y.; Xiu, Z. 1,3-Propanediol. Bioprocessing of Renewable Resources to Commodity Bioproducts, 1st ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2014; pp. 289–326. [Google Scholar]
- Zhuge, B.; Zhang, C.; Fang, H.; Zhuge, J.; Permaul, K. Expression of 1,3-propanediol oxidoreductase and its isoenzyme in Klebsiella pneumoniae for bioconversion of glycerol into 1,3-propanediol. Appl. Microbiol. Biotechnol. 2010, 87, 2177–2184. [Google Scholar] [CrossRef] [PubMed]
- Zeng, A.-P. Pathway and kinetic analysis of 1,3-propanediol production from glycerol fermentation by Clostridium butyricum. Bioprocess Eng. 1996, 14, 169–175. [Google Scholar] [CrossRef]
- Lee, C.S.; Aroua, M.K.; Dauda, W.M.A.W.; Cognet, P.; Pérès-Lucchese, Y.; Fabre, P.-L.; Reynes, O.; Latapie, L. A review: Conversion of bioglycerol into 1,3-propanediol via biological and chemical method. Renew. Sustain. Energy Rev. 2015, 42, 963–972. [Google Scholar] [CrossRef] [Green Version]
- Jensen, T.O.; Kvist, T.; Mikkelsen, M.J.; Westermann, P. Production of 1,3-PDO and butanol by a mutant strain of Clostridium pasteurianum with increased tolerance towards crude glycerol. AMB Express 2012, 2, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbirato, F.; Himmi, E.-H.; Conte, T.; Bories, A. 1,3-Propanediol production by fermentation: An interesting way to valorize glycerin from the ester and ethanol industries. Ind. Crop. Prod. 1998, 7, 281–289. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Fick, M.; Aggelis, G. The effect of raw glycerol concentration on the production of 1,3-propanediol by Clostridium butyricum. J. Chem. Technol. Biotechnol. 2004, 79, 1189–1196. [Google Scholar] [CrossRef]
- González-Pajuelo, M.; Andrade, J.C.; Vasconcelos, I. Production of 1,3-propanediol by Clostridium Butyricum VPI 3266 using a synthetic medium and raw glycerol. J. Ind. Microbiol. Biotechnol. 2004, 31, 442–446. [Google Scholar] [CrossRef]
- Szymanowska-Powałowska, D.; Białas, W. Scale-up of anaerobic 1,3-propanediol production by Clostridium Butyricum DSP1 from crude glycerol. BMC Microbiol. 2014, 14, 45. [Google Scholar] [CrossRef] [Green Version]
- Chatzifragkou, A.; Papanikolaou, S.; Kopsahelis, N.; Kachrimanidou, V.; Dorado, M.P.; Koutinas, A.A. Biorefinery development through utilization of biodiesel industry by-products as sole fermentation feedstock for 1,3-propanediol production. Bioresour. Technol. 2014, 159, 167–175. [Google Scholar] [CrossRef]
- Martins, F.F.; Saab, V.S.; Ribeiro, C.M.S.; Coelho, M.A.Z.; Ferreira, T.F. Production of 1,3-propanediol by Clostridium butyricum growing on biodiesel derived glycerol. Chem. Eng. Trans. 2016, 50, 289–294. [Google Scholar]
- Wang, J.; Lin, M.; Xu, M.; Yang, S.T. Anaerobic fermentation for production of carboxylic acids as bulk chemicals from renewable biomass. Adv. Biochem. Eng. Biotechnol. 2016, 156, 323–362. [Google Scholar] [PubMed]
- Agler, M.T.; Spirito, C.M.; Usack, J.G.; Werner, J.J.; Angenent, L.T. Development of a highly specific and productive process for n-caproic acid production: Applying lessons from methanogenic microbiomes. Water Sci. Technol. 2013, 69, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Yang, J.; Yang, Y.; Luo, L.; Wang, R.; Zhang, Y.; Yuan, H. Consolidated bioprocessing performance of bacterial consortium EMSD5 on hemicellulose for isopropanol production. Bioresour. Technol. 2019, 292, 121965. [Google Scholar] [CrossRef] [PubMed]
- Saini, M.; Wang, Z.W.; Chiang, C.J.; Chao, Y.P. Metabolic engineering of Escherichia coli for production of butyric acid. J. Agric. Food Chem. 2014, 62, 4342–4348. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Yang, R.; Zhao, Y.; Wang, Z.; Liu, Z.; Huang, M.; Zeng, Q. Recent advances and strategies in process and strain engineering for the production of butyric acid by microbial fermentation. Bioresour. Technol. 2018, 253, 343–354. [Google Scholar] [CrossRef]
- Yang, S.T.; Yu, M.; Chang, W.L.; Tang, I.C. Anaerobic fermentations for the production of acetic and butyric acids. In Bioprocessing Technologies in Biorefinery for Sustainable Production of Fuels, Chemicals, and Polymers; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013. [Google Scholar]
- Cavalcante, W.A.; Leitão, R.C.; Gehring, T.A.; Angenent, L.T.; Santaella, S.T. Anaerobic fermentation for n-caproic acid production: A review. Process Biochem. 2017, 54, 106–119. [Google Scholar] [CrossRef]
- Zhu, X.; Zhou, Y.; Wang, Y.; Wu, T.; Li, X.; Li, D.; Tao, Y. Production of high-concentration n-caproic acid from lactate through fermentation using a newly isolated Ruminococcaceae bacterium CPB6. Biotechnol. Biofuels 2017, 10, 102. [Google Scholar] [CrossRef]
- Ge, S.; Usack, J.G.; Spirito, C.M.; Angenent, L.T. Long-term n-caproic acid production from yeast-fermentation beer in an anaerobic bioreactor with continuous product extraction. Environ. Sci. Technol. 2015, 49, 8012–8021. [Google Scholar] [CrossRef]
- Yang, Y.; Hoogewind, A.; Moon, Y.H.; Day, D. Production of butanol and isopropanol with an immobilized Clostridium. Bioprocess Biosyst. Eng. 2016, 39, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Moon, Y.H.; Han, K.J.; Kim, D.; Day, D.F. Enhanced production of butanol and isopropanol from sugarcane molasses using Clostridium beijerinckii optinoii. Biotechnol. Bioprocess Eng. 2015, 20, 871–877. [Google Scholar] [CrossRef]
- Mariano, A.P.; Dias, M.O.; Junqueira, T.L.; Cunha, M.P.; Bonomi, A.; Filho, R.M. Butanol production in a first-generation Brazilian sugarcane biorefinery: Technical aspects and economics of greenfield projects. Bioresour. Technol. 2013, 135, 316–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moncada, J.; Tamayo, J.A.; Cardona, C.A. Integrating first, second, and third generation biorefineries: Incorporating microalgae into the sugarcane biorefinery. Chem. Eng. Sci. 2014, 118, 126–140. [Google Scholar] [CrossRef]
- Rabelo, S.C.; Carrere, H.; Filho, R.M.; Costa, A.C. Production of bioethanol, methane and heat from sugarcane bagasse in a biorefinery concept. Bioresour. Technol. 2011, 102, 7887–7895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalett, O.; Junqueira, T.L.; Dias, M.O.; Jesus, C.D.; Mantelatto, P.E.; Cunha, M.P.; Franco, H.C.J.; Cardoso, T.F.; Filho, R.M.; Bonomi, A.; et al. Environmental and economic assessment of sugarcane first generation biorefineries in Brazil. Clean Technol. Environ. Policy 2012, 14, 399–410. [Google Scholar] [CrossRef]


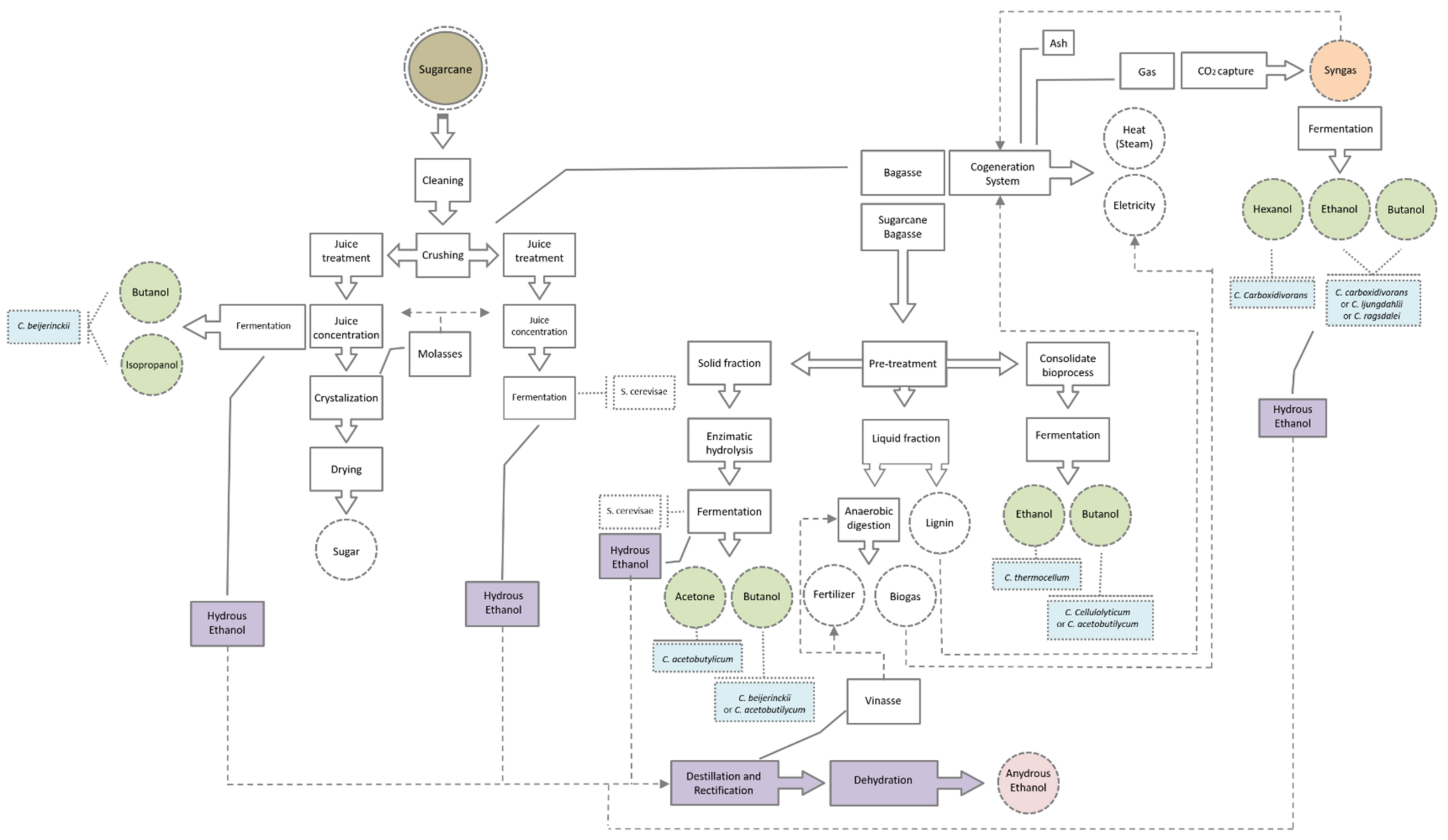{kind=link}
| Species | Main Products | PMax 1 (g/L) | QMax 1 (g/L.h) | Substrate or Feedstock | V 1 (L) | Time (h) 1 | Ref. 1 |
|---|---|---|---|---|---|---|---|
| C. carboxidivorans | Ethanol | 5.55 | NA 2 | 100% CO | 1.2 | 500 | [22] |
| Butyric acid | 1.43 | ||||||
| Butanol | 2.66 | ||||||
| Ethanol | 23.93 | 0.143 | 20% CO, 5% H2, 15% CO2, 60% N2 | 8 | 360 | [23] | |
| C. ragsdalei | Ethanol | 13.2 3 | NA | 40% CO, 30% H2, 30% CO2 | 2.4 | 290 | [24] |
| 19 | NA | 38% CO, 28.5% H2, 28.5% CO2, 5% N2 | 2.4 | 470 | [25] | ||
| C. ljungdahlii | Ethanol | 20.7 4 | 0.374 | 60% CO, 35% H2, 5% CO2 | 4 | 2014 | [26] |
| 48 | NA | 55% CO, 20% H2, 10% CO2, 15% N2 | 1 | 560 | [27] | ||
| C. kluyveri | n-caproic acid | 8.42 | 0.07 | Ethanol and acetate (10:1) | 0.1 | 120 | [28] |
| C. beijerinckii | Butanol | 34.77 4 | 0.48 | Corn stover hydrolysate | 1.25 | 72 | [29] |
| 11.92 | 0.17 | Corn cob hydrolysate | 50 | 72 | [30] | ||
| 11.77 | 0.10 | Sugarcane juice | 1.5 | 100 | [31] | ||
| Isopropanol | 3.41 | NA | Glucose | 0.06 | 48 | [32] | |
| C. saccharobutylicum | Butanol | 12.8 | 0.257 | Corn stover hydrolysate | 5 | 48 | [33] |
| ABE 5 | 19.9 | - | |||||
| C. saccharo- perbutylacetonicum | Butanol | 64.6 6 | 0.673 | Glucose | 0.5 | 96 | [34] |
| Acetone | 30.6 | 0.319 | |||||
| ABE | 97.3 | 1.01 | |||||
| C. thermocellum | Ethanol | 13.66 | NA | Cellulose | 0.02 | 120 | [35] |
| C. phytofermentans | Ethanol | 7.0 | 0.03 | AFEX™ -treated corn stover 7 | 0.3 | 250 | [36] |
| C. acetobutylicum | Butanol | 44.6 8 | 2.13 | Glucose | 4 | 88 | [37] |
| Ethanol | 33 9 | 0.47 | Glucose | 0.2 | 185 | [38] | |
| Acetone | 7.59 10 | 0.13 | Glucose | 5 | 54 | [39] | |
| C. butyricum | 1,3-propanediol | 93.7 | 3.30 | Pure glycerol | 1 | 32 | [40] |
| C. cellulolyticum | Ethanol | 2.5 10 | 0.007 | Crystalline AVICEL 11 Cellulose | 0.05 | 336 | [41] |
| C. cellulovorans | Butanol | 3.37 | 0.046 | Corn cob | 2 | 72 | [42] |
| C. pasteurianum | Butanol | 17.8 | 7.8 12 | Pure glycerol | 0.09 | 48 | [43] |
| 1,3-propanediol | 9.5 | 0.2 | 0.09 | 48 | |||
| C. perfringens | 1,3-propanediol | 40 | 2.0 | Pure glycerol | 3 | 48 | [44] |
| Conversion Route | Process | End Products |
|---|---|---|
| Thermo-chemical | Combustion | Heat and power |
| Gasification | Hydrogen, alcohol, olefins, gasoline, diesel | |
| Liquefaction | Hydrogen, methane, oils | |
| Pyrolysis | Hydrogen, olefins, oils, specialty chemicals | |
| Biochemical | Fermentation | Second-generation fuels (ethanol, butanol), chemicals and biochemicals |
| Anaerobic digestion | Methane, fertilizer (digestate) | |
| Hybrid [7] | Pyrolysis + fermentation | Ethanol, organic acids (butyric acid, acetic acid), 2,3 butanediol |
| Reactor | Agitation (rpm) | Specific Gas Flow Rate (vvm) | Microorganism | Gas | kLa (h−1) | Ref |
|---|---|---|---|---|---|---|
| BCR | n/a | 0.4 | n/a | CO | 72.0 | [160] |
| CHF | n/a | n/a | n/a | CO | 85.7–946.6 | [146] |
| CSTR | 300 | n/a | C. ljungdahlii | CO | 14.9 | [161] |
| CSTR | 400 | n/a | C. ljungdahlii | CO | 21.5 | [161] |
| CSTR | 400 | 0–0.32 | n/a | Syngas | 38.0 | [162] |
| CSTR | 500 | n/a | C. ljungdahlii | CO | 22.8 | [161] |
| CSTR | 600 | n/a | C. ljungdahlii | CO | 23.8 | [161] |
| CSTR | 700 | n/a | C. ljungdahlii | CO | 35.5 | [161] |
| GLR | n/a | 1.67 | n/a | CO | 129.6 | [154] |
| HFMBR | n/a | 0.625 | n/a | CO | 1096.2 | [23] |
| HFMBR | n/a | 0.029 | n/a | CO | 385.0 | [163] |
| HFR | n/a | 0.037 | n/a | CO | 420 | [158] |
| MBR | n/a | 0.00625–0.0625 | n/a | Syngas | 450.0 | [154] |
| PBC with microbubble sparger | n/a | 0–0.021 | R. rubrum | Syngas | 2.1 for CO | [148] |
| STR | 300 | 0–0.032 | C. ljungdahlii | Syngas | 35.0 for CO | [148] |
| STR | 300 | 0–0.032 | SBR mixed culture | Syngas | 31.0 for CO | [148] |
| STR | 300 | 2.7 | n/a | CO | 166.09 ± 13.15 | [157] |
| STR | 400 | 0.14–0.86 | n/a | CO | 10.8–155.0 | [164] |
| STR | 400 | 0.70–2.14 | n/a | CO | 72.0–153.0 | [155] |
| STR | 400 | 0.36–1.07 | n/a | CO | 72.0–122.4 | [165] |
| STR | 400 | 0–0.032 | R. rubrum | Syngas | 101.0 for CO | [148] |
| STR | 450 | 0–0.032 | R. rubrum | Syngas | 101.0 for CO | [148] |
| STR | 500 | 0.36–1.07 | n/a | CO | 129.6–144.0 | [165] |
| STR | 500 | 2.0–2.7 | n/a | CO | 11.48 ± 57.79–399.06 ± 26.80 | [157] |
| STR | 600 | 0.36–1.07 | n/a | CO | 147.6–208.8 | [165] |
| STR | 650 | 0.36–1.07 | n/a | CO | 172.8–252.0 | [165] |
| STR | 700 | 0.36–1.07 | n/a | CO | 187.2–288.0 | [165] |
| STR with microbubble sparger | 300 | n/a | SBR mixed culture | Syngas | 104.0 for CO | [148] |
| TBR | n/a | 0–0.021 | n/a | Syngas | 22.0 | [162] |
| TBR | n/a | 0–0.021 | C. ljungdahlii | Syngas | 137.0 for CO | [148] |
| TBR | n/a | 0–0.021 | R. rubrum | Syngas | 55.5 for CO | [148] |
| TBR | n/a | 0–0.021 | SBR mixed culture | Syngas | 121 for CO | [148] |
| Feedstock | Microorganism | Reactor | PEt 1 (g/L) | QEt 2 (g/L.h) | Ref. 3 |
|---|---|---|---|---|---|
| Syngas | C. carboxidivorans P7 | HFM-BR | 23.93 | 0.14 | [23] |
| Syngas | C. rasgdalei P11 | STR | 25.26 | 0.001 | [172] |
| Syngas | C. ljunghdalli ERI-2 | CSTR/Bubble column | 19.73 | 0.37 | [26] |
| Syngas | C. ljunghdalli PETC | CSTR | 19 | 0.30 | [173] |
| Syngas | C. carboxidivorans P7 | h-RPB | 7 | 0.28 | [174] |
| Syngas | C. ljunghdalli | Batch | 0.49 | 0.03 | [175] |
| Syngas | C. carboxidivorans P7 | Batch | 2 | 0.02 | [176] |
| CO | C. autoethanogenum DSM 10061 | Continuous gas-fed | 7.14 | - | [177] |
| Glucose | C. saccharoperbutylacetonium pSH2 | Batch | 7.9 | 0.11 | [178] |
| Glucose | C. acetobutylicum hbd::int(69) | Fed-batch | 33 | 0.5 | [38] |
| Glucose | C. carboxidivorans P7 | Batch | 2.34 | 0.01 | [179] |
| Lignocellulose | C. thermocellum ATCC31924 | Batch | 2.45 | 0.02 | [180] |
| Lignocellulose | C. cellulolyticum H10 | Batch | 2.5 | - | [41] |
| Lignocellulose and starch | C. acetobutylicum NBRC13948 | SHF | 1.7 | 0.018 | [180] |
| Lignocellulose and starch | C. acetobutylicum NBRC13948 | SSF | 1.5 | 0.01 | [180] |
| Cellobiose | C. thermocellum LL1275 | Batch | 5 | 0.07 | [63] |
| Cellobiose | C. cellulovorans 83151-adhE2 | CBP | 2.03 | 0.03 | [181] |
| Cellobiose | C. phytofermentans ATCC700394 | CBP | 7 | 0.03 | [36] |
| Glycerol | C. pasteurianum MTCC6013 | Immobilized cells | 1.94 | 0.01 | [182] |
| Strain | FS | Strategy | Scale | C1,3-PDO 1 | Y1,3-PDO 2 | Q1,3-PDO 3 | Ref. 4 |
|---|---|---|---|---|---|---|---|
| C. butyricum CNCM1211 | PG 5 | B 7 | 1-L | 37.1 | 0.53 | 1.32 | [221] |
| C. butyricum CNCM1211 | CG 6 | B | 1-L | 63.4 | 0.57 | 0.63 | [221] |
| C. butyricum F2b | CG | B | 2-L | 47.1 | 0.52 | 1.34 | [222] |
| C. butyricum F2b | CG | C 8 | 2-L | 44.0 | 0.51 | 1.76 | [222] |
| C. butyricum VPI 3266 | PG | C | 2-L | 29.7 | 0.62 | 2.98 | [220] |
| C. butyricum VPI 3266 | CG | C | 2-L | 31.5 | 0.50 | 3.15 | [223] |
| C. butyricum AKR102a | PG | FB 9 | 1-L | 93.7 | 0.52 | 3.30 | [40] |
| C. butyricum AKR102a | CG | FB | 1-L | 76.2 | 0.51 | 2.30 | [40] |
| C. butyricum AKR102a | CG | FB | 200-L | 61.5 | 0.53 | 2.10 | [40] |
| C. butyricum DSP1 | CG | B | 6.6-L | 37.6 | 0.53 | 1.12 | [224] |
| C. butyricum DSP1 | CG | FB | 6.6-L | 71.0 | 0.54 | 0.68 | [224] |
| C. butyricum DSP1 | CG | B | 42-L | 36.4 | 0.52 | 1.13 | [224] |
| C. butyricum DSP1 | CG | B | 150-L | 37.2 | 0.53 | 1.33 | [224] |
| C. butyricum VPI1718 | CG | RB 10 | 1-L | 65.5 | 0.52 | 1.15 | [225] |
| C. butyricum NCIMB 8082 | CG | B | 1-L | 32.2 | 0.52 | 2.38 | [226] |
| C. butyricum NCIMB 8082 | CG | FB | 1-L | 29.8 | 0.48 | 2.55 | [226] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liberato, V.; Benevenuti, C.; Coelho, F.; Botelho, A.; Amaral, P.; Pereira, N., Jr.; Ferreira, T. Clostridium sp. as Bio-Catalyst for Fuels and Chemicals Production in a Biorefinery Context. Catalysts 2019, 9, 962. https://doi.org/10.3390/catal9110962
Liberato V, Benevenuti C, Coelho F, Botelho A, Amaral P, Pereira N Jr., Ferreira T. Clostridium sp. as Bio-Catalyst for Fuels and Chemicals Production in a Biorefinery Context. Catalysts. 2019; 9(11):962. https://doi.org/10.3390/catal9110962
Chicago/Turabian StyleLiberato, Vanessa, Carolina Benevenuti, Fabiana Coelho, Alanna Botelho, Priscilla Amaral, Nei Pereira, Jr., and Tatiana Ferreira. 2019. "Clostridium sp. as Bio-Catalyst for Fuels and Chemicals Production in a Biorefinery Context" Catalysts 9, no. 11: 962. https://doi.org/10.3390/catal9110962