The Potential Applications of Bacillus sp. and Pseudomonas sp. Strains with Antimicrobial Activity against Phytopathogens, in Waste Oils and the Bioremediation of Hydrocarbons


 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
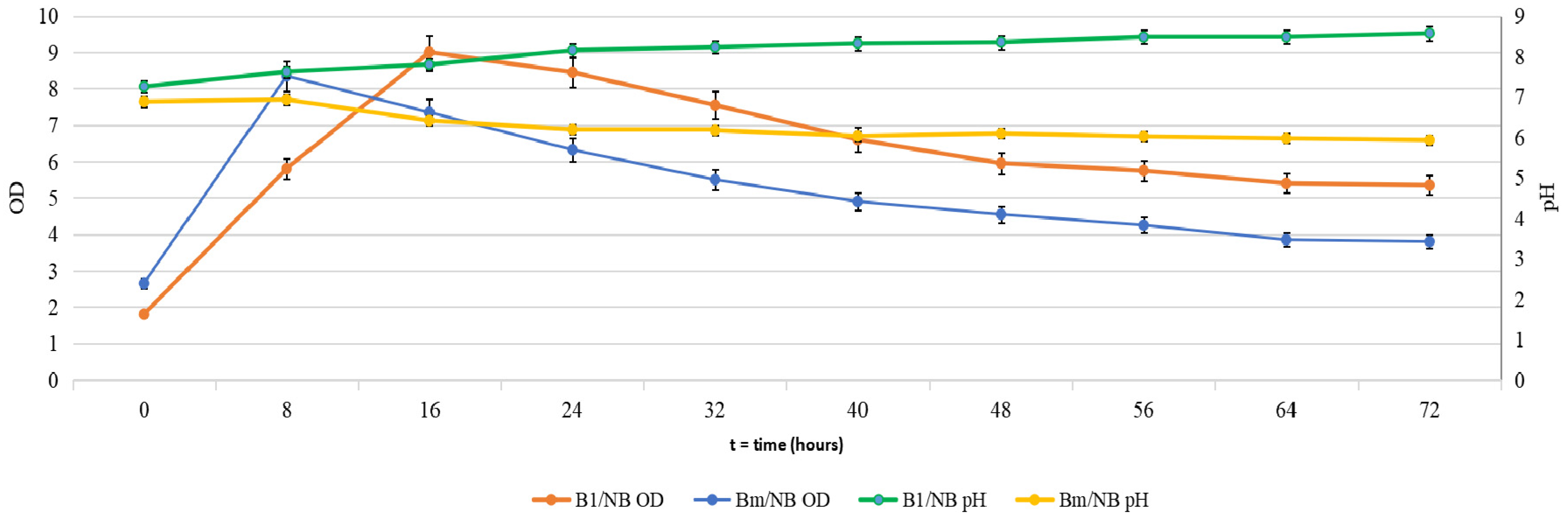
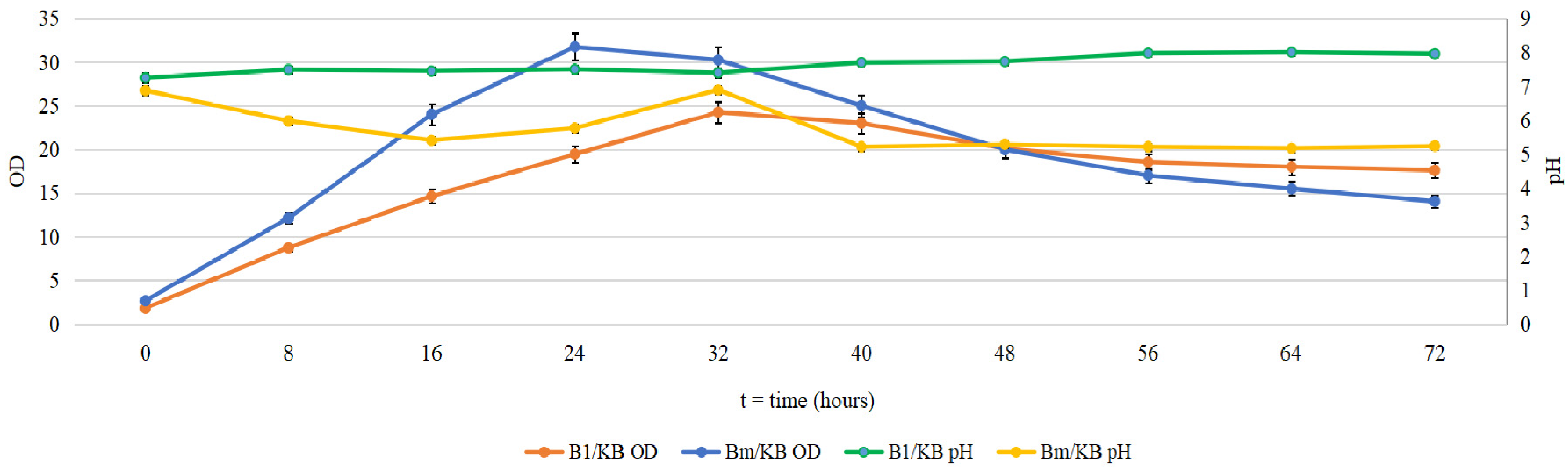
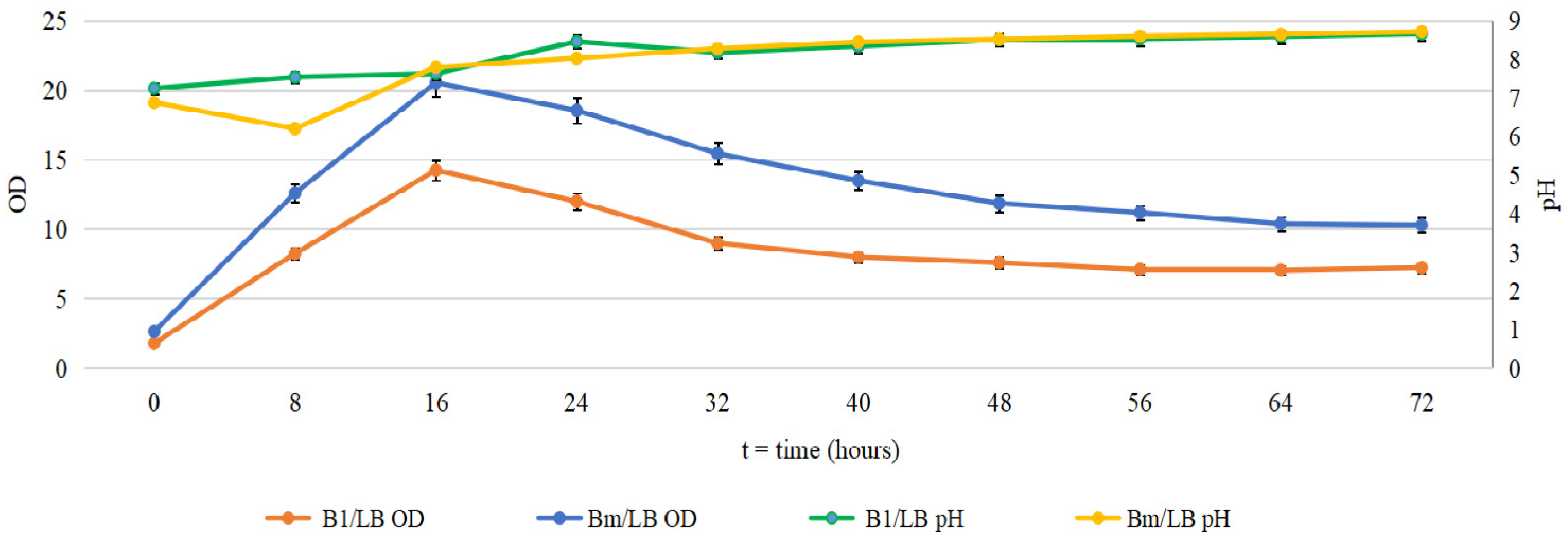
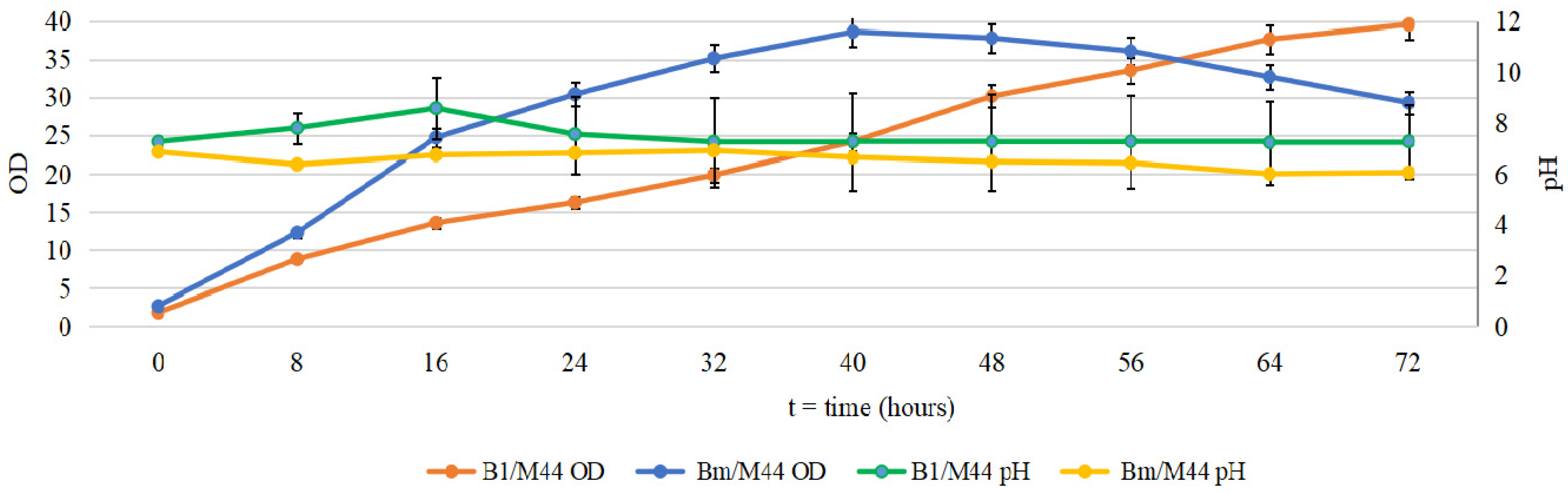
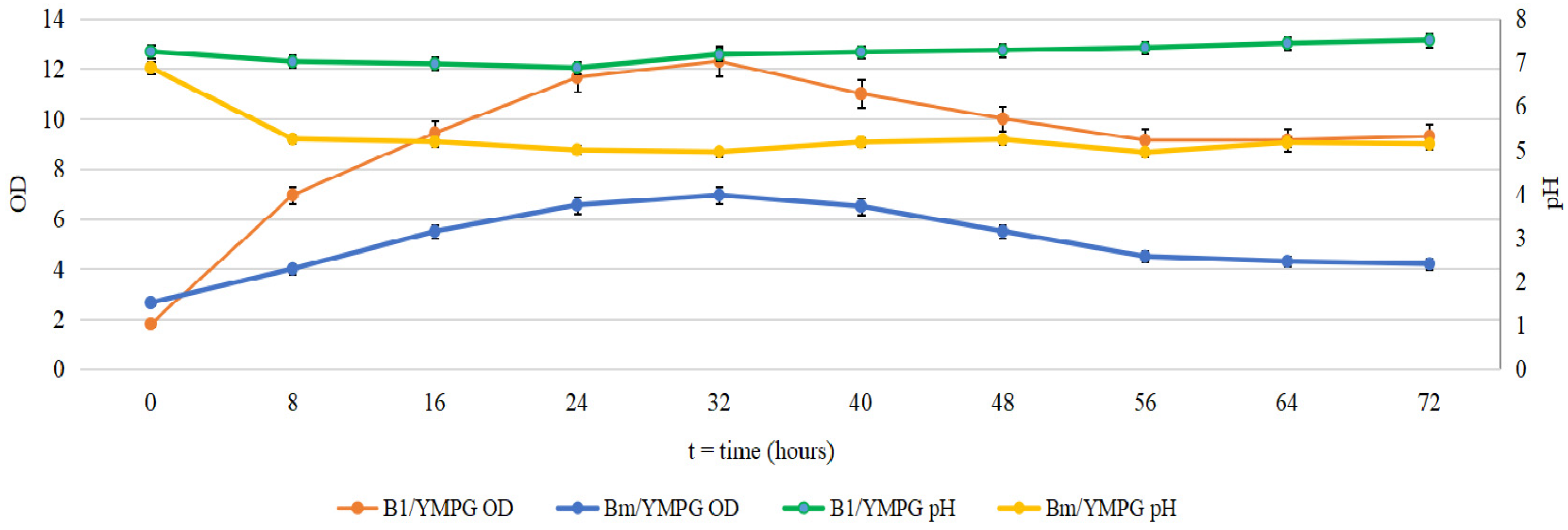
2.1. Cultivation of B. mycoides and P. putida in Several Media
2.2. Antimicrobial Activity
2.2.1. Antimicrobial Activity of P. putida and B. mycoides against E. carotovora and X. campestris
2.2.2. Antimicrobial Activity of the Biosurfactants from P. putida and B. mycoides Supernatants against E. carotovora and X. campestris
2.3. Enzyme Production
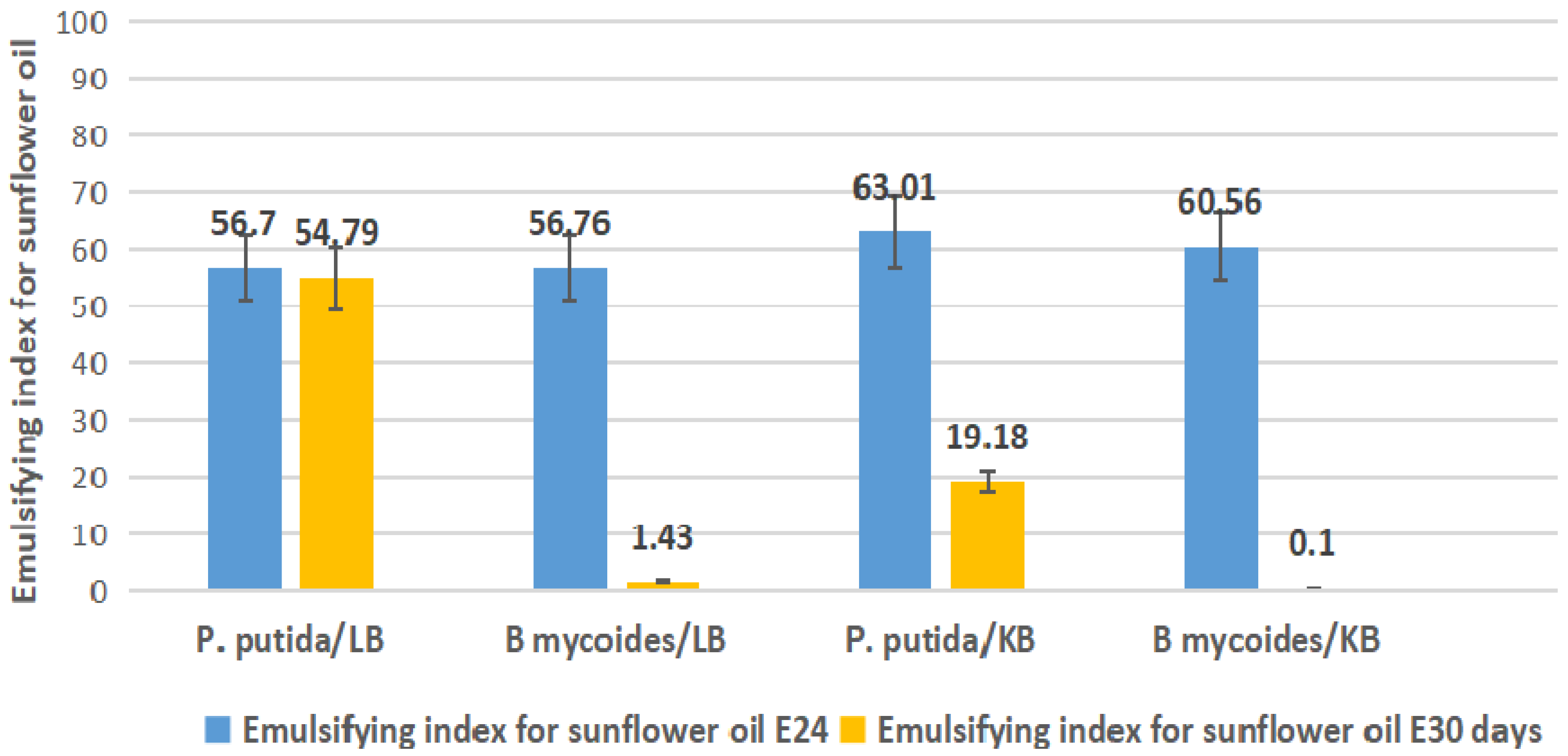
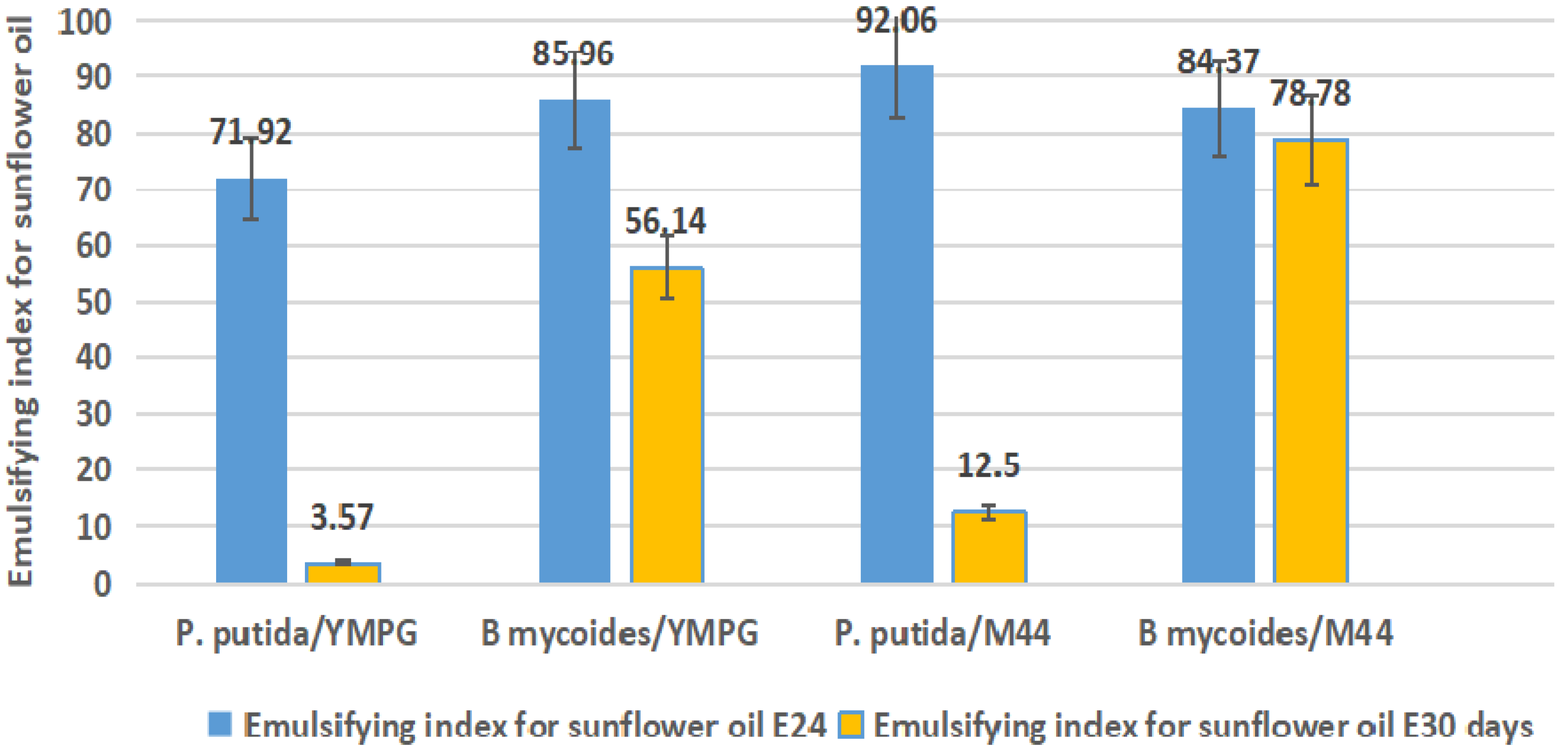
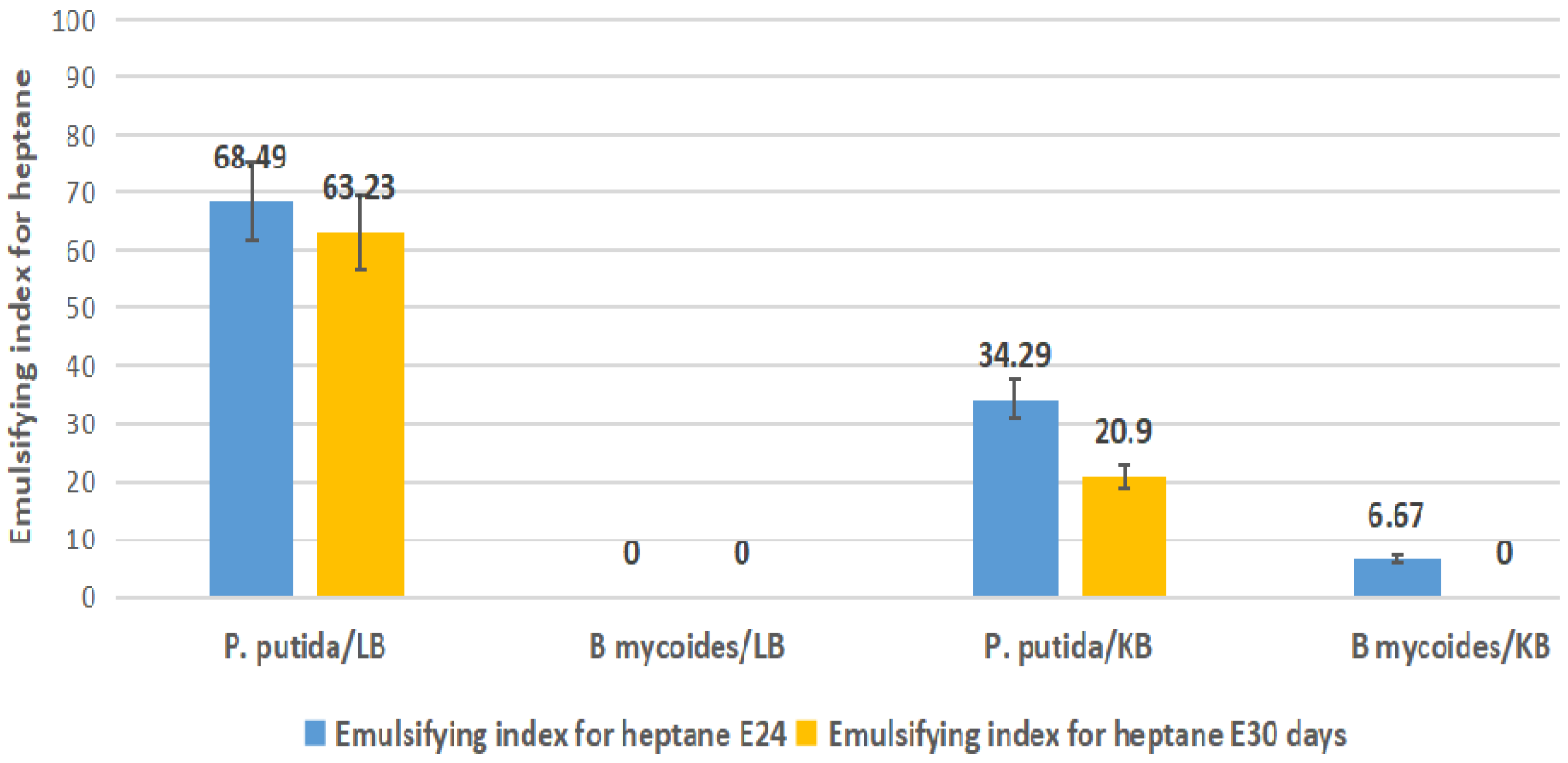
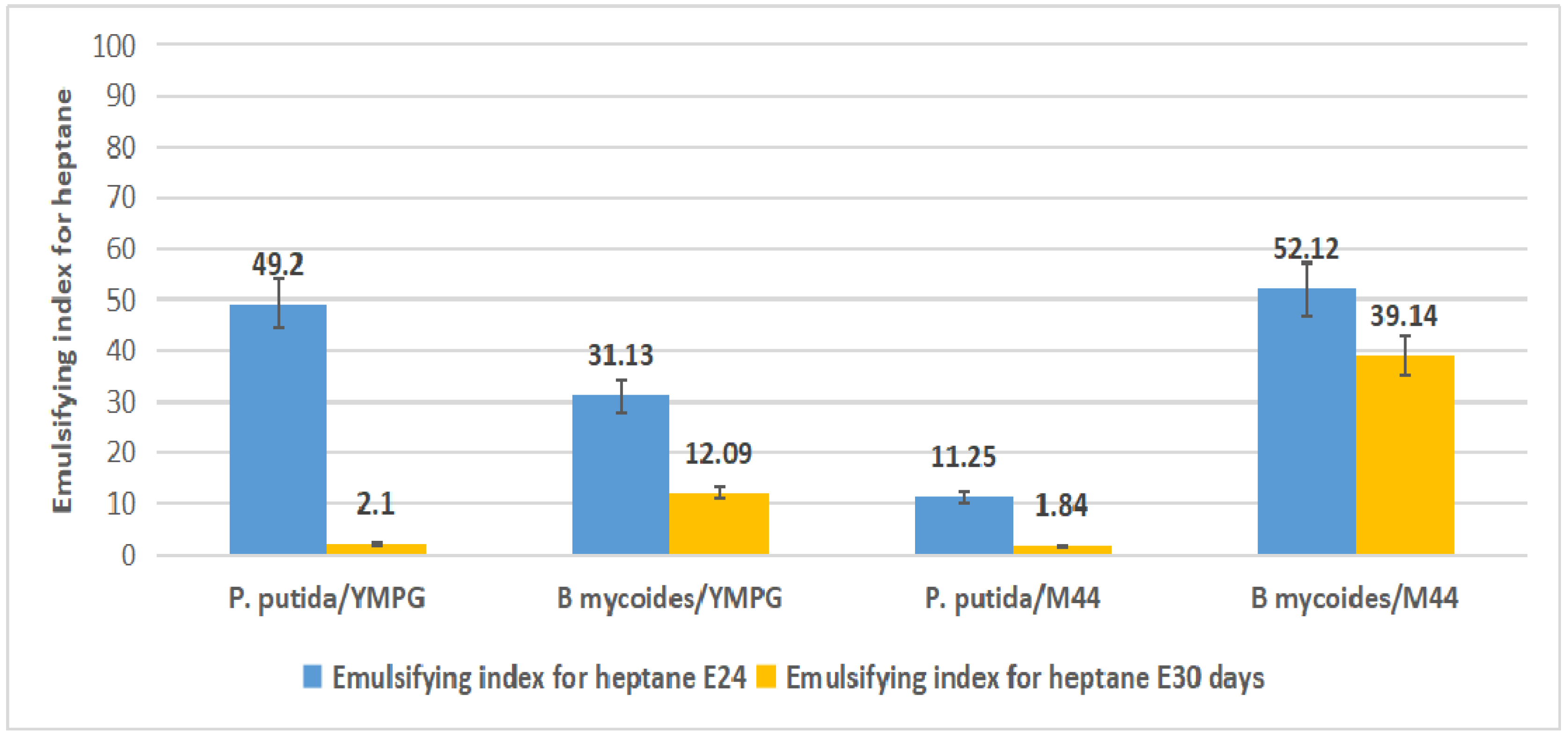
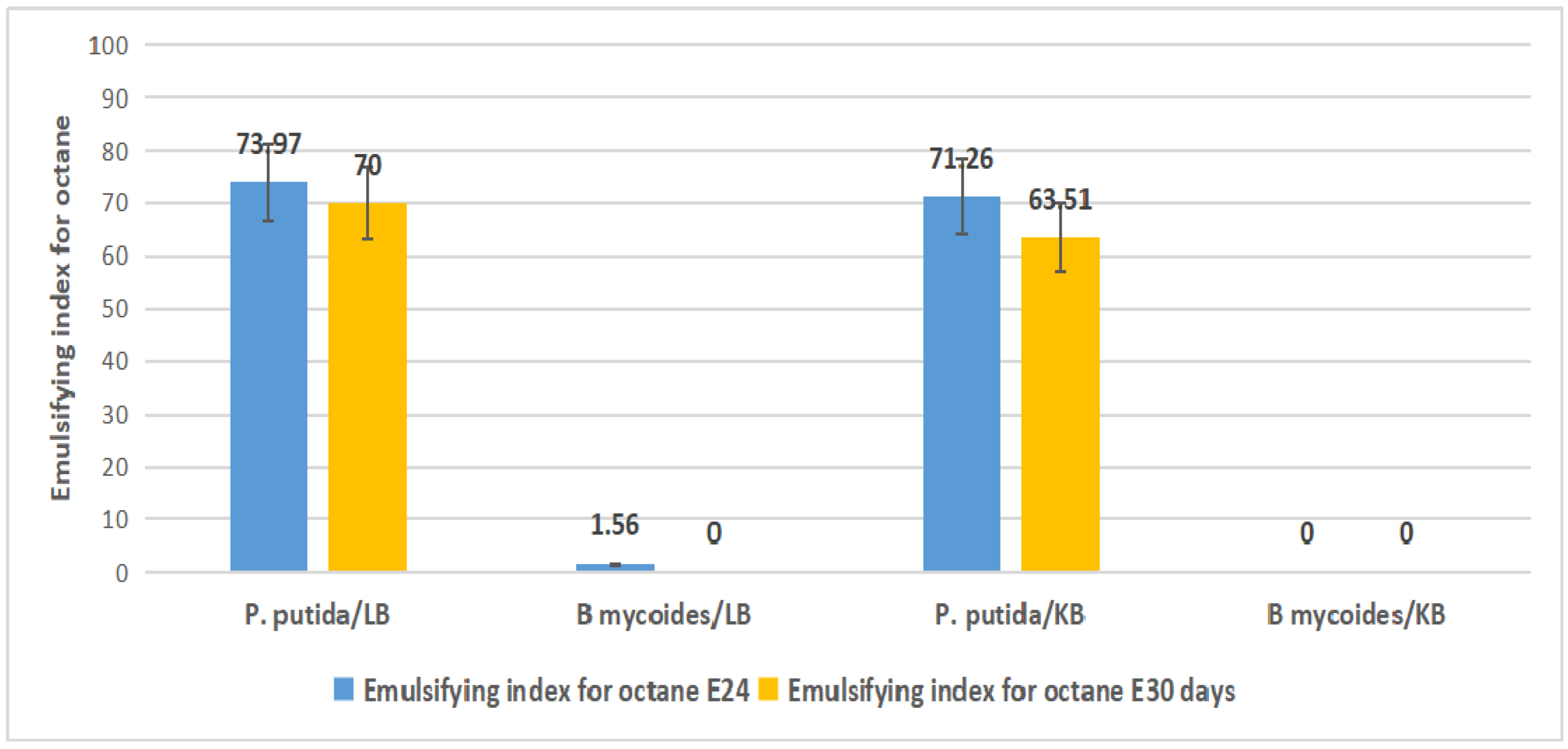
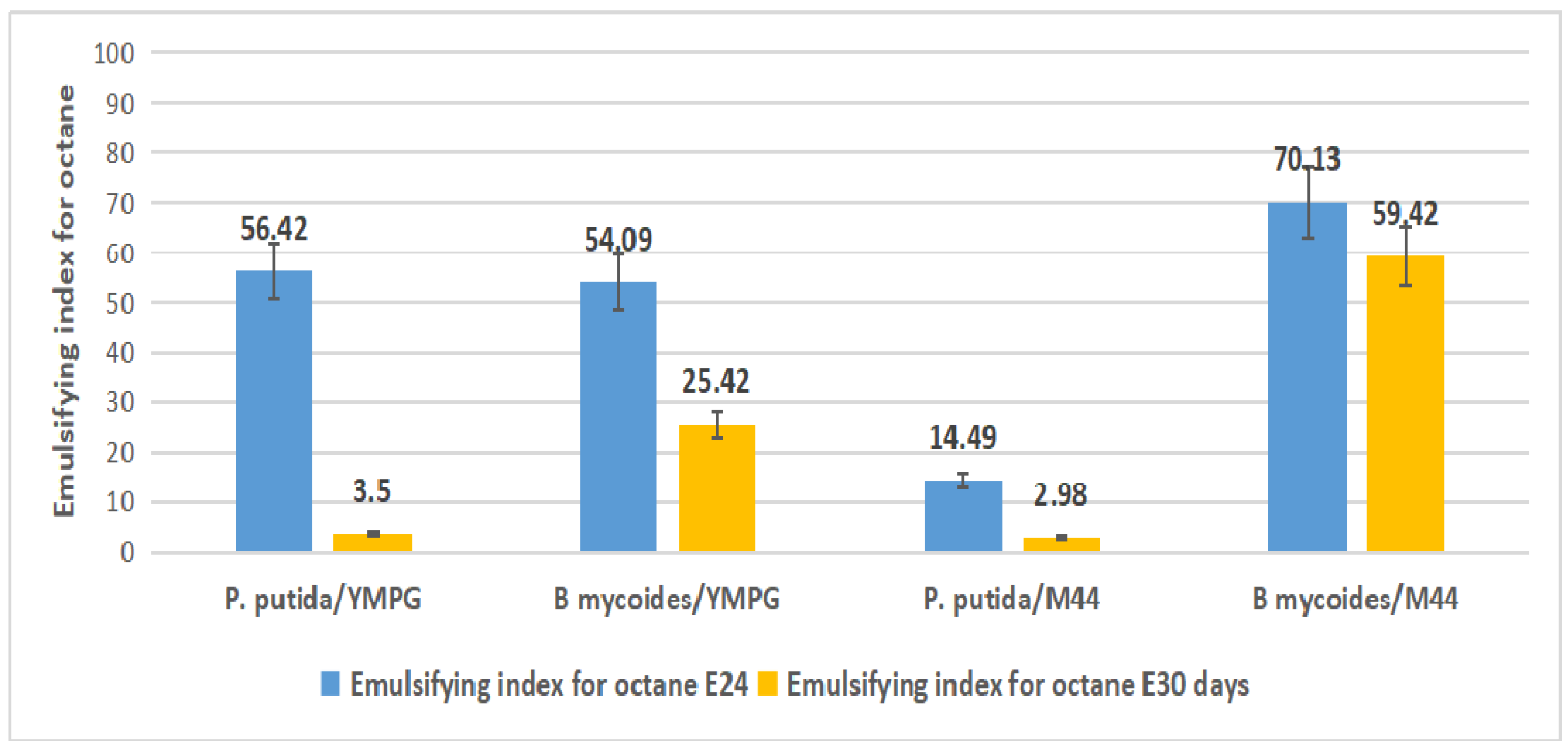
2.4. Emulsification Index
3. Materials and Methods
3.1. Biologic Material
3.2. Culture Media
3.3. Preservation of Strains and Cultivation Conditions
3.4. Antimicrobial Activity Assay
3.5. Enzyme Production


3.6. Emulsification Index Test
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bubici, G.; Kaushal, M.; Prigigallo, M.I.; Cabanás, G.-L.C.; Mercado-Blanco, J. Biological Control Agents Against Fusarium Wilt of Banana. Front. Microbiol. 2019, 10, 616. [Google Scholar] [CrossRef]
- Caulier, S.; Gillis, A.; Colau, G.; Licciardi, F.; Liépin, M.; Desoignies, N.; Modrie, P.; Legrève, A.; Mahillon, J.; Bragard, C. Versatile Antagonistic Activities of Soil-Borne Bacillus spp. and Pseudomonas spp. against Phytophthora infestans and Other Potato Pathogens. Front. Microbiol. 2018, 9, 143. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; De Bruijn, I.; Nybroe, O.; Ongena, M. Natural functions of lipopeptides from Bacillus and Pseudomonas: More than surfactants and antibiotics. FEMS Microbiol. Rev. 2010, 34, 1037–1062. [Google Scholar] [CrossRef]
- Fira, D.; Dimkic, I.; Beric, T.; Lozo, J.; Stankovic, S. Biological control of plant pathogens by Bacillus species. J. Biotechnol. 2018, 285, 44–55. [Google Scholar] [CrossRef]
- Gross, H.; Loper, J.E. Genomics of secondary metabolite production by Pseudomonas spp. Nat. Prod. Rep. 2009, 26, 1408–1446. [Google Scholar] [CrossRef]
- Santos, D.K.; Rufino, R.D.; Luna, J.M.; Santos, V.A.; Sarubbo, L.A. Biosurfactants: Multifunctional Biomolecules of the 21st Century. Int. J. Mol. Sci. 2016, 17, 401. [Google Scholar] [CrossRef]
- Singh, P.; Patil, Y.; Rale, V. Biosurfactant production: Emerging trends and promising strategies. J. Appl. Microbiol. 2018, 126, 2–13. [Google Scholar] [CrossRef]
- Gomaa, E.Z. Antimicrobial activity of a biosurfactant produced by Bacillus licheniformis strain M104 grown on whey. Braz. Arch. Biol. Technol. 2013, 56, 259–268. [Google Scholar] [CrossRef]
- Ndlovu, T.; Rautenbach, M.; Vosloo, J.A.; Khan, S.; Khan, W. Characterisation and antimicrobial activity of biosurfactant extracts produced by Bacillus amyloliquefaciens and Pseudomonas aeruginosa isolated from a wastewater treatment plant. AMB Express 2017, 7, 108. [Google Scholar] [CrossRef]
- Katz, E.; Demain, A.L. The peptide antibiotics of Bacillus: Chemistry, biogenesis and possible functions. Bacteriol. Rev. 1977, 41, 449. [Google Scholar]
- Grossman, A.D. Genetic networks controlling the initiation of sporulation and the development of genetic competence in Bacillus subtilis. Annu. Rev. Genet. 1995, 29, 477–508. [Google Scholar] [CrossRef] [PubMed]
- Caulier, S.; Nannan, C.; Gillis, A.; Licciardi, F.; Bragard, C.; Mahillon, J. Overview of the antimicrobial compounds produced by members of the Bacillus subtilis group. Front. Microbiol. 2019, 10, 302. [Google Scholar] [CrossRef] [PubMed]
- Nihorimbere, V.; Ongena, M.; Smargiassi, M.; Thonart, P. Beneficial effect of the rhizosphere microbial community for plant growth and health. Biotechnol. Agron. Soc. Environ. 2011, 15, 327–337. [Google Scholar]
- Sachdev, D.P.; Cameotra, S.S. Biosurfactants in agriculture. Appl. Microbiol. Biotechnol. 2013, 97, 1005–1016. [Google Scholar] [CrossRef] [PubMed]
- Rendueles, O.; Ghigo, J.M. Multi-species biofilms: How to avoid unfriendly neighbors. FEMS Microbiol. Rev. 2012, 36, 972–989. [Google Scholar] [CrossRef]
- Zhang, F.; Gu, W.; Xu, P.; Tang, S.; Xie, K.; Huang, X.; Huang, Q. Effects of alkyl polyglycoside (APG) on composting of agricultural wastes. Waste Manag. 2011, 31, 1333–1338. [Google Scholar] [CrossRef]
- Rosenberg, E.; Ron, E.Z. High- and low-molecular-mass microbial surfactants. Appl. Microbiol. Biotechnol. 1999, 52, 154–162. [Google Scholar] [CrossRef]
- Silva, E.J.; Correa, P.F.; Almeida, D.G.; Luna, J.M.; Rufino, R.D.; Sarubbo, L.A. Recovery of contaminated marine environments by biosurfactant-enhanced bior-emediation. Colloids Surf. B Biointerfaces 2018, 172, 127–135. [Google Scholar] [CrossRef]
- Chandankere, R.; Yao, J.; Cai, M.; Masakorala, K.; Jain, A.K.; Choi, M.M. Properties and characterization of biosurfactant in crude oil biodegradation by bacterium Bacillus methylotrophicus USTBa. Fuel 2014, 122, 140–148. [Google Scholar] [CrossRef]
- Desai, J.D.; Banat, I.M. Microbial production of surfactants and their commercial potential. Microbiol. Mol. Biol. Rev. 1997, 61, 47–64. [Google Scholar]
- Banat, I.M.; Makkar, R.S.; Cameotra, S.S. Potential commercial applications of microbial surfactants. Appl. Microbiol. Biotechnol. 2000, 53, 495–508. [Google Scholar] [CrossRef] [PubMed]
- Banat, I.M. Biosurfactants production and possible uses in microbial enhanced oil recovery and oil pollution remediation: A review. Bioresour. Technol. 1995, 51, 1–12. [Google Scholar] [CrossRef]
- Barkay, T.; Navon-Venezia, S.; Ron, E.Z.; Rosenberg, E. Enhancement of solubilization and biodegradation of polyaromatic hydrocarbons by the bioemulsifier alasan. Appl. Environ. Microbiol. 1991, 65, 2697–2702. [Google Scholar]
- Christofi, N.; Ivshina, I.B. Microbial surfactants and their use in field studies of soil remediation. J. Appl. Microbiol. 2002, 93, 915–929. [Google Scholar] [CrossRef] [PubMed]
- Kosaric, N. Biosurfactants and their application for soil bioremediation. Food Technol. Biotechnol. 2001, 39, 295–304. [Google Scholar]
- Calvo, C.; Manzanera, M.; Silva-Castro, G.A.; Uad, I.; González-López, J. Application of bioemulsifiers in soil oil bioremediation processes. Future prospects. Sci. Total Environ. 2009, 407, 3634–3640. [Google Scholar] [CrossRef]
- Mouafo, T.H.; Mbawala, A.; Ndjouenkeu, R. Effect of Different Carbon Sources on Biosurfactants’ Production by Three Strains of Lactobacillus spp. Biomed Res. Int. 2018, 2018, 5034783. [Google Scholar] [CrossRef]
- Nielsen, T.H.; Sørensen, D.; Tobiasen, C.; Andersen, J.B.; Christophersen, C.; Givskov, M.; Sørensen, J. Antibiotic and Biosurfactant Properties of Cyclic Lipopeptides Produced by Fluorescent Pseudomonas spp. from the Sugar Beet Rhizosphere. Appl. Environ. Microbiol. 2002, 68, 3416–3423. [Google Scholar] [CrossRef]
- Petatán-Sagahón, I.; Anducho-Reyes, M.A.; Silva-Rojas, H.V.; Arana-Cuenca, A.; Tellez-Jurado, A.; Cárdenas-Álvarez, I.O.; Mercado-Flores, Y. Isolation of Bacteria with Antifungal Activity against the Phytopathogenic Fungi Stenocarpella maydis and S macrospora. Int. J. Mol. Sci. 2011, 12, 5522–5537. [Google Scholar] [CrossRef]
- Available online: https://www.sigmaaldrich.com/technical-documents/protocols/biology/enzymatic-assay-of-a-amylase.html (accessed on 5 November 2019).
- Available online: https://www.sigmaaldrich.com/content/dam/sigma-aldrich/docs/Sigma/General_Information/lipase_-_titrimetric_ph_72.pdf (accessed on 4 November 2019).
- Available online: https://www.sigmaaldrich.com/technical-documents/protocols/biology/enzymatic-assay-of-catalase.html (accessed on 5 November 2019).
- Das, P.; Yang, X.P.; Ma, L.Z. Analysis of biosurfactants from industrially viable Pseudomonas strain isolated from crude oil suggests how rhamnolipids congeners affect emulsification property and antimicrobial activity. Front. Microbiol. 2014, 5, 696. [Google Scholar] [CrossRef]
- El-Sheshtawy, H.S.; Doheim, M.M. Selection of Pseudomonas aeruginosa for biosurfactant production and studies of its antimicrobial activity. Egypt. J. Pet. 2014, 23, 1–6. [Google Scholar] [CrossRef]
- Chong, H.; Li, Q. Microbial production of rhamnolipids: Opportunities, challenges and strategies. Microb. Cell Fact. 2017, 16, 137. [Google Scholar] [CrossRef] [PubMed]
- Gnanamani, A.; Kavitha, V.; Radhakrishnan, N.; Mandal, A.B. Bioremdiation of crude oil contamination using microbial surface-active agents isolation production and characterization. J. Bioremed. Biodegrad. 2010, 1, 107. [Google Scholar] [CrossRef]
- Lima, T.M.S.; Fonseco, A.F.; Leao, B.A.; Houndeer, A.H.; Totala, M.R.; Borges, A.C. Oil recovery from oil fuel oil storage tank sludges using biosurfactants. J. Bioremed. Biodegrad. 2011, 2, 125–137. [Google Scholar]
- Anyanwu, C.U.; Obi, S.K.C.; Okolo, B.M. Lipopeptide biosurfactant produced by Serratia marcescens NSK-1 strain isolated from petroleum contaminated soil. J. Appl. Sci. Res. 2011, 7, 79–87. [Google Scholar]
- Diab, A.; Sami, S.; Diab, A. Production, Characterization and Application of a New Biosurfactant Derived from Egyptian Sunflower Seeds. Int. J. Sci. Res. 2016, 5, 602–612. [Google Scholar]
- Pathak, K.V.; Keharia, H. Application of extracellular lipopeptide biosurfactant produced by endophytic Bacillus subtilis K1 isolated from aerial roots of banyan (Ficus benghalensis) in microbially enhanced oil recovery (MEOR). 3 Biotech 2014, 4, 41–48. [Google Scholar] [CrossRef]
- Sharma, R.; Singh, J.; Verma, N. Production, characterization and environmental applications of biosurfactants from Bacillus amyloliquefaciens and Bacillus subtilis. Biocatal. Agric. Biotechnol. 2018, 16, 132–139. [Google Scholar] [CrossRef]
- Available online: http://cfarm.ncpri.ro/files/cmii-iccf/CMII-ICCF.pdf (accessed on 27 September 2019).
- Soare, M.G.; Tomulescu, C.; Petrescu, M.M.; Lupescu, I.; Moscovici, M.; Popa, O.; Băbeanu, N. Antimicrobial activity of newly isolated Bacillus sp. and Pseudomonas sp. strains and their potential use as biocontrol agents. Sci. Bull. Ser. F Biotechnol. 2017, XXI, 81–86. [Google Scholar]
- Nitschke, M.; Pastore, G.M. Production and properties of a surfactant obtained from Bacillus subtilis grown on cassava wastewater. Bioresour. Technol. 2006, 97, 336–341. [Google Scholar] [CrossRef]
- Moro, G.V.; Almeida, R.T.R.; Napp, A.P.; Porto, C.; Pilau, E.J.; Lüdtke, D.S.; Moro, A.V.; Vainstein, M.H. Identification and ultra-high-performance liquid chromatography coupled with high-resolution mass spectrometry characterization of biosurfactants, including a new surfactin, isolated from oil-contaminated environments. Microb. Biotechnol. 2018, 11, 759–769. [Google Scholar] [CrossRef] [PubMed]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strain | Inhibition Zones (mm) against E. carotovora ICCF 138 | Inhibition Zones (mm) against X. campestris ICCF 274 | ||
|---|---|---|---|---|
| Experiment I | Experiment II | Experiment I | Experiment II | |
| P. putida (B1) | 35 ± 0.58 | 24 ± 0.29 | 36 ± 0.58 | 22 ± 0.29 |
| B. mycoides (Bm) | 60 ± 0.58 | 27 ± 0.58 | 43 ± 0.58 | 30 ± 0.29 |
| Control | - | - | - | - |
| Supernatants of the Strains | Inhibition Zone Diameters (in mm) Against E. c. ICCF138 | Inhibition Zone Diameters (in mm) Against X. c. ICCF274 | |
|---|---|---|---|
| P. putida (B1)/Medium * | 1 * | 7 ± 0.87 | 8 ± 0.29 |
| 2 * | 10 ± 0.58 | 10 ± 0.29 | |
| 3 * | 10 ± 0.58 | 9 ± 0.58 | |
| 4 * | 11 ± 0.58 | 12 ± 0.29 | |
| 5 * | 8 ± 0.58 | 9 ± 0.58 | |
| B. mycoides (Bm)/Medium * | 1 * | 8 ± 0.58 | 7 ± 0.58 |
| 2 * | 10 ± 0.29 | 9 ± 0.58 | |
| 3 * | 9 ± 0.58 | 10 ± 0.58 | |
| 4 * | 10 ± 0.58 | 11 ± 0.87 | |
| 5 * | 8 ± 0.58 | 7 ± 0.58 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soare, M.-G.; Lakatos, E.S.; Ene, N.; Malo, N.; Popa, O.; Babeanu, N. The Potential Applications of Bacillus sp. and Pseudomonas sp. Strains with Antimicrobial Activity against Phytopathogens, in Waste Oils and the Bioremediation of Hydrocarbons. Catalysts 2019, 9, 959. https://doi.org/10.3390/catal9110959
Soare M-G, Lakatos ES, Ene N, Malo N, Popa O, Babeanu N. The Potential Applications of Bacillus sp. and Pseudomonas sp. Strains with Antimicrobial Activity against Phytopathogens, in Waste Oils and the Bioremediation of Hydrocarbons. Catalysts. 2019; 9(11):959. https://doi.org/10.3390/catal9110959
Chicago/Turabian StyleSoare (Vladu), Mariana-Gratiela, Elena Simina Lakatos, Nicoleta Ene, Nereida Malo (Dalanaj), Ovidiu Popa, and Narcisa Babeanu. 2019. "The Potential Applications of Bacillus sp. and Pseudomonas sp. Strains with Antimicrobial Activity against Phytopathogens, in Waste Oils and the Bioremediation of Hydrocarbons" Catalysts 9, no. 11: 959. https://doi.org/10.3390/catal9110959