
The Studies of Sepharose-Immobilized Lipases: Combining Techniques for the Enhancement of Activity and Thermal Stability
 , , ,
, , ,
Abstract
:1. Introduction
2. Results and Discussion
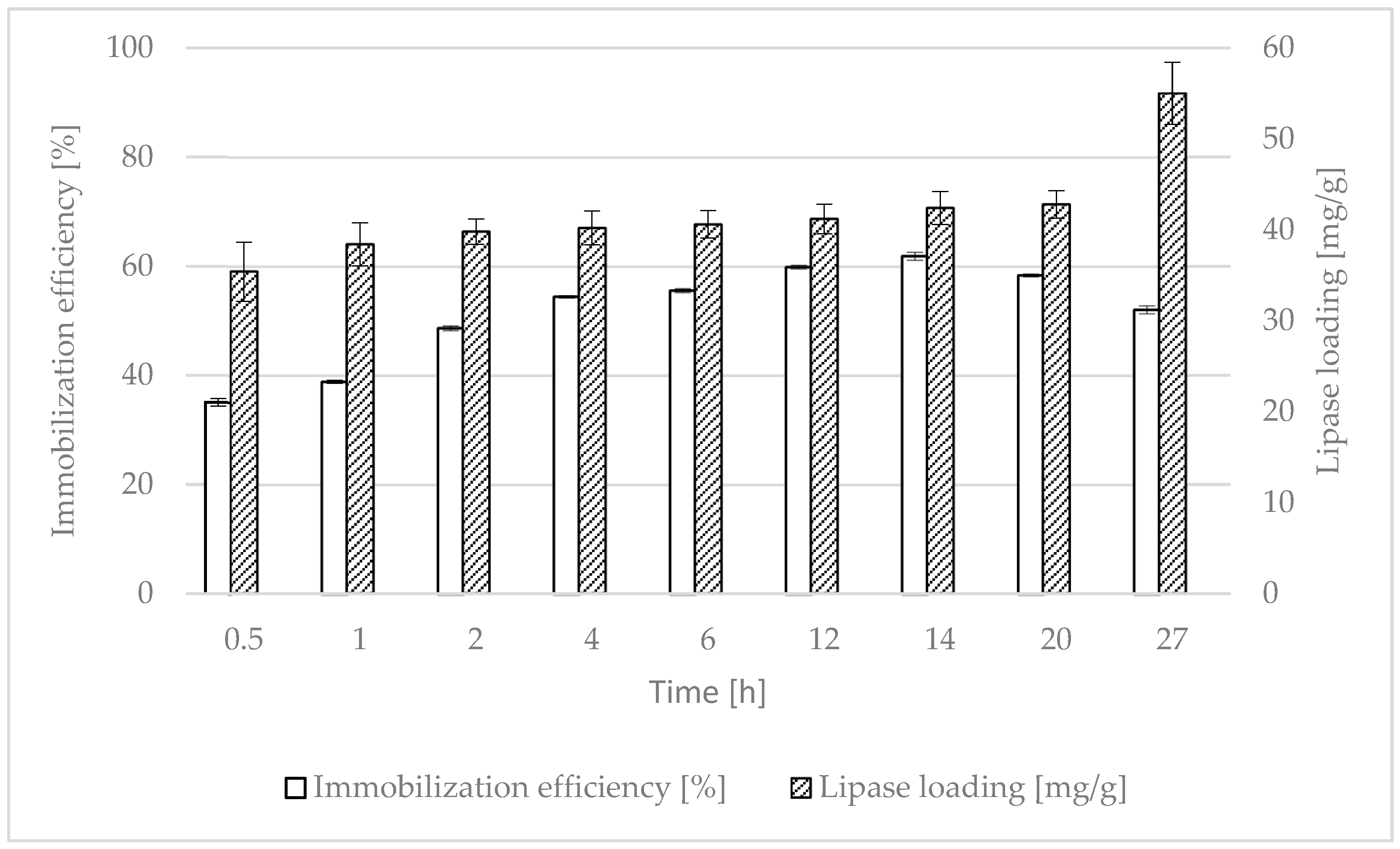
2.1. Effect of Immobilization Time on CALB Catalytic Activity and Lipase Loading
2.2. Effect of Immobilization Temperature on CALB Activity
2.3. Effect of Immobilization Conditions on CRL-OF Enzymatic Activity
2.4. Climatic Chamber Thermal Stability Tests of CALB and CRL-OF
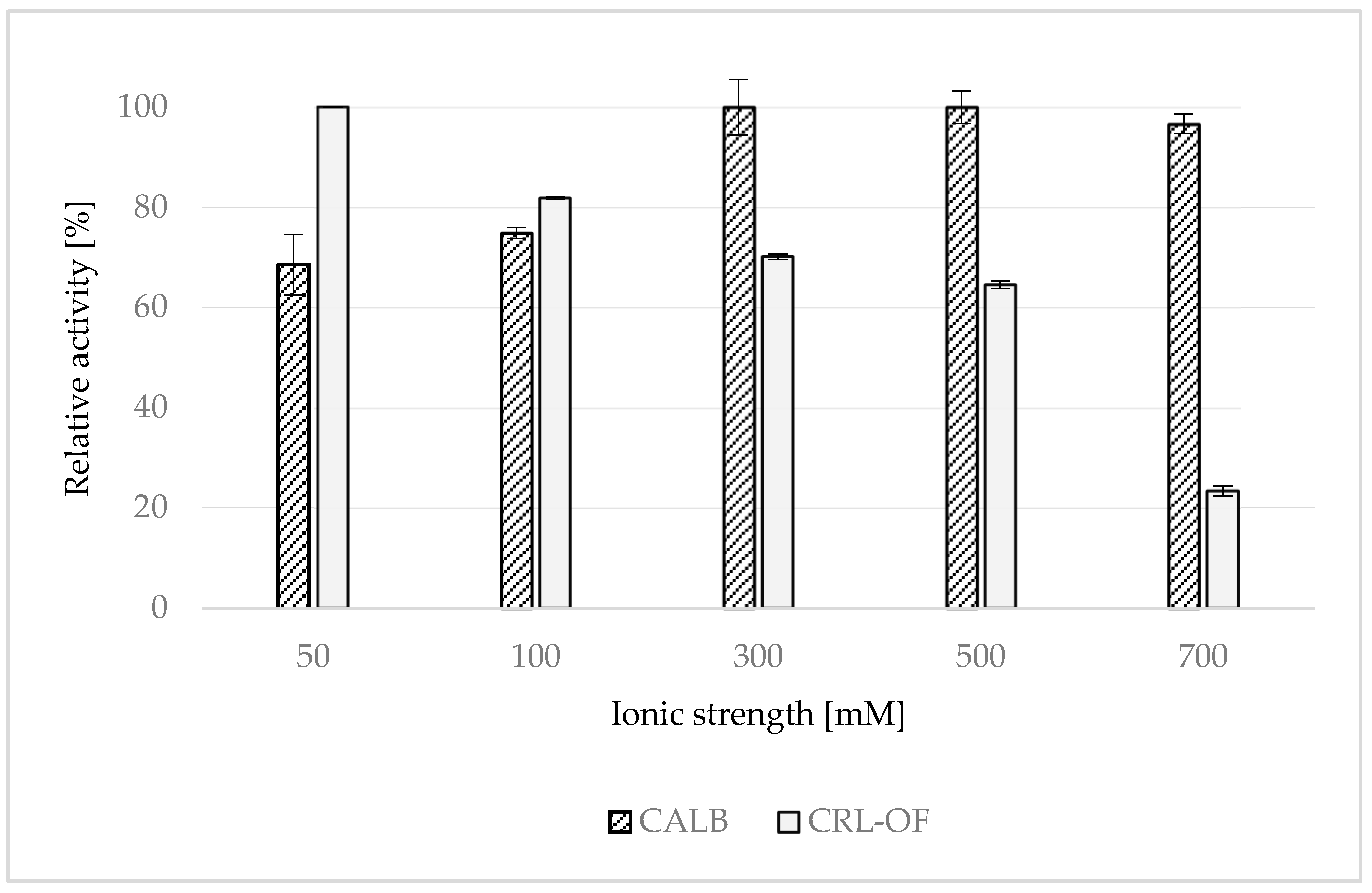
2.5. Effect of Lipase Storage in Buffers of Different Ionic Strength—24 h Stability Test
3. Materials and Methods
3.1. Materials
3.2. Octyl Sepharose CL-4B Preparation Protocol
3.3. Immobilization of CALB onto Octyl-Sepharose CL-4B
- (a)
- immobilization time—the samples were mixed for 5 min at room temperature and then stayed at a temperature of 4 °C for 0.5, 1, 2, 4, 6, 12, 14, 20, and 27 h;
- (b)
- temperature—the samples were mixed for 5 min at room temperature or in an ice bath, and then CALB was incubated for 14 h in the following temperature conditions: 4 °C (refrigerator); 22 °C (room temperature); and 37 °C; additionally selected samples were stirred during 14 h of incubation (at room temperature and 37 °C).
3.4. Immobilization of CRL-OF onto Octyl-Sepharose CL-4B
- (a)
- immobilization time—the samples were mixed for 5 min at room temperature and then stayed at a temperature of 4 °C for 0.5, 1, 2, 4, 6, 12, 14, 20, and 27 h.
3.5. Determination of the Amount of Immobilized CALB and CRL-OF by Bradford Method
3.6. Lipolytic Activity of CALB and CRL-OF
3.7. Climatic Chamber Thermal Stability Tests of CALB and CRL-OF in Aqueous Buffers
3.8. Effect of Lipase Storage in Buffers of Different Ionic Strength—24 h Stability Test
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Choi, J.M.; Han, S.S.; Kim, H.S. Industrial applications of enzyme biocatalysis: Current status and future aspects. Biotechnol. Adv. 2015, 33, 1443–1454. [Google Scholar] [CrossRef]
- Anastas, P.; Eghbali, N. Green Chemistry: Principles and Practice. Chem. Soc. Rev. 2010, 39, 301–312. [Google Scholar] [CrossRef]
- Dulęba, J.; Siódmiak, T.; Marszałł, M.P. Amano Lipase PS from Burkholderia cepacia—Evaluation of the Effect of Substrates and Reaction Media on the Catalytic Activity. Curr. Org. Chem. 2020, 24, 798–807. [Google Scholar] [CrossRef]
- Zisis, T.; Freddolino, P.L.; Turunen, P.; van Teeseing, M.C.F.; Rowan, A.E.; Blank, K.G. Interfacial Activation of Candida antarctica Lipase B: Combined Evidence from Experiment and Simulation. Biochemistry 2015, 54, 5969–5979. [Google Scholar] [CrossRef]
- Dulęba, J.; Siódmiak, T.; Marszałł, M.P. The influence of substrate systems on the enantioselective and lipolytic activity of immobilized Amano PS from Burkholderia cepacia lipase (APS-BCL). Process. Biochem. 2022, 120, 126–137. [Google Scholar] [CrossRef]
- Siódmiak, T.; Dulęba, J.; Kocot, N.; Watróbska-Świetlikowska, D.; Marszałł, M.P. The High ‘Lipolytic Jump’ of Immobilized Amano A Lipase from Aspergillus niger in Developed ‘ESS Catalytic Triangles’ Containing Natural Origin Substrates. Catalysts 2022, 12, 853. [Google Scholar] [CrossRef]
- de Maria, P.D.; Sanchez-Montero, J.M.; Sinisterra, J.V.; Alcantara, A.R. Understanding Candida rugosa lipases: An overview. Biotechnol. Adv. 2006, 24, 180–196. [Google Scholar] [CrossRef] [PubMed]
- Cunha, D.; Bartkevihi, L.; Robert, J.M.; Cipolatti, E.P.; Ferreira, A.T.S.; Oliveira, D.M.P.; Gomes-Neto, F.; Almeida, R.V.; Fernandez-Lafuente, R.; Freire, D.M.G.; et al. Structural differences of commercial and recombinant lipase B from Candida antarctica: An important implication on enzymes thermostability. Int. J. Biol. Macromol. 2019, 140, 761–770, Reprinted from Publication with permission from Elsevier (License Number-5518720480304). [Google Scholar] [CrossRef]
- Cygler, M.; Schrag, J.D. Structure and conformational flexibility of Candida rugosa lipase. Biochim. Biophys. Acta 1999, 1441, 205–214, Reprinted from Publication with permission from Elsevier (License Number-5518150100052). [Google Scholar] [CrossRef]
- Van Tassel, L.; Moilanen, A.; Ruddock, L.W. Efficient production of wild-type lipase B from Candida antarctica in the cytoplasm of Escherichia coli. Protein Expr. Purif. 2020, 165, 105498–105503. [Google Scholar] [CrossRef] [PubMed]
- Tejo, B.A.; Salleh, A.B.; Pleiss, J. Structure and dynamics of Candida rugosa lipase: The role of organic solvent. J. Mol. Model. 2004, 10, 358–366. [Google Scholar] [CrossRef]
- Siódmiak, T.; Haraldsson, G.G.; Dulęba, J.; Ziegler-Borowska, M.; Siódmiak, J.; Marszałł, M.P. Evaluation of Designed Immobilized Catalytic Systems: Activity Enhancement of Lipase B from Candida antarctica. Catalysts 2020, 10, 876. [Google Scholar] [CrossRef]
- Gabriele, F.; Spreti, N.; Del Giacco, T.; Germani, R.; Tiecco, M. Effect of Surfactant Structure on the Superactivity of Candida rugosa Lipase. Langmuir 2018, 34, 11510–11517. [Google Scholar] [CrossRef]
- Bramer, C.; Schreiber, S.; Scheper, T.; Beutel, S. Continuous purification of Candida antarctica lipase B using 3-membrane adsorber periodic counter-current chromatography. Eng. Life Sci. 2018, 18, 414–424. [Google Scholar] [CrossRef]
- Vanleeuw, E.; Winderickx, S.; Thevissen, K.; Lagrain, B.; Dusselier, M.; Cammue, B.P.A.; Sels, B.F. Substrate-Specificity of Candida rugosa Lipase and Its Industrial Application. ACS Sustain. Chem. Eng. 2019, 7, 15828–15844. [Google Scholar] [CrossRef]
- Kim, S.K.; Park, Y.C.; Lee, H.H.; Jeon, S.T.; Min, W.K.; Seo, J.H. Simple Amino Acid Tags Improve Both Expression and Secretion of Candida antarctica Lipase B in Recombinant Escherichia coli. Biotechnol. Bioeng. 2015, 112, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.C.; Lee, L.C.; Sava, V.; Shaw, J.F. Multiple mutagenesis of non-universal serine codons of the Candida rugosa LIP2 gene and biochemical characterization of purified recombinant LIP2 lipase overexpressed in Pichia pastoris. Biochem. J. 2002, 366, 603–611. [Google Scholar] [CrossRef]
- Hirata, D.B.; Albuquerque, T.L.; Rueda, N.; Virgen-Ortiz, J.J.; Tacias-Pascacio, V.G.; Fernandez-Lafuente, R. Evaluation of different immobilized lipases in transesterification reactions using tributyrin: Advantages of the heterofunctional octyl agarose beads. J. Mol. Catal. B Enzym. 2016, 133, 117–123. [Google Scholar] [CrossRef]
- Vakhlu, J.; Kour, A. Yeast lipases: Enzyme purification, biochemical properties and gene cloning. Electron. J. Biotechnol. 2006, 9, 69–85. [Google Scholar] [CrossRef]
- Siódmiak, T.; Mangelings, D.; Heyden, Y.V.; Ziegler-Borowska, M.; Marszałł, M.P. High Enantioselective Novozym 435-Catalyzed Esterification of (R,S)-Flurbiprofen Monitored with a Chiral Stationary Phase. Appl. Biochem. Biotechnol. 2015, 175, 2769–2785. [Google Scholar] [CrossRef] [PubMed]
- Marszałł, M.P.; Siódmiak, T. Immobilization of Candida rugosa lipase onto magnetic beads for kinetic resolution of (R,S)-ibuprofen. Catal. Commun. 2012, 24, 80–84. [Google Scholar] [CrossRef]
- Schmitt, J.; Brocca, S.; Schmid, R.D.; Pleiss, J. Blocking the tunnel: Engineering of Candida rugosa lipase mutants with short chain length specificity. Protein Eng. 2002, 15, 595–601. [Google Scholar] [CrossRef]
- Melais, N.; Aribi-Zouioueche, L.; Riant, O. The effect of the migrating group structure on enantioselectivity in lipase-catalyzed kinetic resolution of 1-phenylethanol. C. R. Chim. 2016, 19, 971–977. [Google Scholar] [CrossRef]
- Thangaraj, B.; Solomon, P.R. Immobilization of Lipases—A Review. Part II: Carrier Materials. Chembioeng. Rev. 2019, 6, 167–194. [Google Scholar] [CrossRef]
- Thangaraj, B.; Solomon, P.R. Immobilization of Lipases—A Review. Part I: Enzyme Immobilization. Chembioeng. Rev. 2019, 6, 157–166. [Google Scholar] [CrossRef]
- Mateo, C.; Palomo, J.M.; Fernandez-Lorente, G.; Guisan, J.M.; Fernandez-Lafuente, R. Improvement of enzyme activity, stability and selectivity via immobilization techniques. Enzyme Microb. Technol. 2007, 40, 1451–1463. [Google Scholar] [CrossRef]
- Boudrant, J.; Woodley, J.M.; Fernandez-Lafuente, R. Parameters necessary to define an immobilized enzyme preparation. Process. Biochem. 2020, 90, 66–80. [Google Scholar] [CrossRef]
- Arana-Pena, S.; Rios, N.S.; Carballares, D.; Goncalves, L.R.B.; Fernandez-Lafuente, R. Immobilization of lipases via interfacial activation on hydrophobic supports: Production of biocatalysts libraries by altering the immobilization conditions. Catal. Today 2021, 362, 130–140. [Google Scholar] [CrossRef]
- Peirce, S.; Tacias-Pascacio, V.G.; Russo, M.E.; Marzocchella, A.; Virgen-Ortiz, J.J.; Fernandez-Lafuente, R. Stabilization of Candida antarctica Lipase B (CALB) Immobilized on Octyl Agarose by Treatment with Polyethyleneimine (PEI). Molecules 2016, 21, 751. [Google Scholar] [CrossRef]
- Aryee, A.N.A.; Simpson, B.K. Immobilization of Lipase from Grey Mullet. Appl. Biochem. Biotechnol. 2012, 168, 2105–2122. [Google Scholar] [CrossRef] [PubMed]
- Arana-Pena, S.; Rios, N.S.; Carballares, D.; Mendez-Sanchez, C.; Lokha, Y.; Goncalves, L.R.B.; Fernandez-Lafuente, R. Effects of Enzyme Loading and Immobilization Conditions on the Catalytic Features of Lipase From Pseudomonas fluorescens Immobilized on Octyl-Agarose Beads. Front. Bioeng. Biotechnol. 2020, 8, 36. [Google Scholar] [CrossRef] [PubMed]
- Zucca, P.; Fernandez-Lafuente, R.; Sanjust, E. Agarose and Its Derivatives as Supports for Enzyme Immobilization. Molecules 2016, 21, 1577. [Google Scholar] [CrossRef] [PubMed]
- Arana-Pena, S.; Carballares, D.; Corberan, V.C.; Fernandez-Lafuente, R. Multi-Combilipases: Co-Immobilizing Lipases with Very Different Stabilities Combining Immobilization via Interfacial Activation and Ion Exchange. The Reuse of the Most Stable Co-Immobilized Enzymes after Inactivation of the Least Stable Ones. Catalysts 2020, 10, 1207. [Google Scholar] [CrossRef]
- Cejudo-Sanches, J.; Orrego, A.H.; Jaime-Mendoza, A.; Ghobadi, R.; Moreno-Perez, S.; Fernandez-Lorente, G.; Rocha-Martin, J.; Guisan, J.M. High stabilization of immobilized Rhizomucor miehei lipase by additional coating with hydrophilic crosslinked polymers: Poly-allylamine/Aldehyde-dextran. Process Biochem. 2020, 92, 156–163. [Google Scholar] [CrossRef]
- Manoel, E.A.; Pinto, M.; dos Santos, J.C.S.; Tacias-Pascacio, V.G.; Freire, D.M.G.; Pinto, J.C.; Fernandez-Lafuente, R. Design of a core-shell support to improve lipase features by immobilization. RSC Adv. 2016, 6, 62814–62824. [Google Scholar] [CrossRef]
- Manoel, E.A.; dos Santos, J.C.S.; Freire, D.M.G.; Rueda, N.; Fernandez-Lafuente, R. Immobilization of lipases on hydrophobic supports involves the open form of the enzyme. Enzyme Microb. Technol. 2015, 71, 53–57. [Google Scholar] [CrossRef]
- Alnoch, R.C.; dos Santos, L.A.; de Almeida, J.M.; Krieger, N.; Mateo, C. Recent Trends in Biomaterials for Immobilization of Lipases for Application in Non-Conventional Media. Catalysts 2020, 10, 697. [Google Scholar] [CrossRef]
- Hamdan, S.H.; Maiangwa, J.; Ali, M.S.M.; Normi, Y.M.; Sabri, S.; Leow, T.C. Thermostable lipases and their dynamics of improved enzymatic properties. Appl. Microbiol. Biotechnol. 2021, 105, 7069–7094. [Google Scholar] [CrossRef]
- Ishak, S.N.H.; Masomian, M.; Kamarudin, N.H.A.; Ali, M.S.M.; Leow, T.C.; Rahman, R.N.Z.R.A. Changes of Thermostability, Organic Solvent, and pH Stability in Geobacillus zalihae HT1 and Its Mutant by Calcium Ion. Int. J. Mol. Sci. 2019, 20, 2561. [Google Scholar] [CrossRef] [PubMed]
- Kawata, T.; Ogino, H. Amino acid residues involved in organic solvent-stability of the LST-03 lipase. Biochem. Biophys. Res. Commun. 2010, 400, 384–388. [Google Scholar] [CrossRef]
- Elgharbawy, A.A.; Hayyan, A.; Hayyan, M.; Rashid, S.N.; Nor, M.R.M.; Zulkifli, M.Y.; Alias, Y.; Mirghani, M.E.S. Shedding Light on Lipase Stability in Natural Deep Eutectic Solvents. Chem. Biochem. Eng. Q. 2018, 32, 359–370. [Google Scholar] [CrossRef]
- Kornecki, J.F.; Carballares, D.; Morellon-Sterling, R.; Siar, E.; Kashefi, S.; Chafiaa, M.; Arana-Pena, S.; Rios, N.S.; Goncalves, L.R.B.; Fernandez-Lafuente, R. Influence of phosphate anions on the stability of immobilized enzymes. Effect of enzyme nature, immobilization protocol and inactivation conditions. Process. Biochem. 2020, 95, 288–296. [Google Scholar] [CrossRef]
- Jiang, C.Y.; Cheng, C.C.; Hao, M.; Wang, H.B.; Wang, Z.Y.; Shen, C.; Cheong, L.Z. Enhanced catalytic stability of lipase immobilized on oxidized and disulfide-rich eggshell membrane for esters hydrolysis and transesterification. Int. J. Biol. Macromol. 2017, 105, 1328–1336. [Google Scholar] [CrossRef] [PubMed]
- Coskun, G.; Ciplak, Z.; Yildiz, N.; Mehmetoglu, U. Immobilization of Candida antarctica Lipase on Nanomaterials and Investigation of the Enzyme Activity and Enantioselectivity. Appl. Biochem. Biotechnol. 2021, 193, 430–445. [Google Scholar] [CrossRef] [PubMed]
- Badoei-Dalfard, A.; Shahba, A.; Zaare, F.; Sargazi, G.; Seyedalipour, B.; Karami, Z. Lipase immobilization on a novel class of Zr-MOF/electrospun nanofibrous polymers: Biochemical characterization and efficient biodiesel production. Int. J. Biol. Macromol. 2021, 192, 1292–1303. [Google Scholar] [CrossRef] [PubMed]
- Zusfahair, D.; Ningsih, R.; Nurhandayani, S.; Lestari, P.; Hermawan, D. Immobilization of Lipase From Azospirillum sp. PRD1 Using Chitosan Alginate as Supporting Agent. Malays. J. Sci. 2014, 10, 195–199. [Google Scholar] [CrossRef]
- Corovic, M.; Mihailovic, M.; Banjanac, K.; Carevic, M.; Milivojevic, A.; Milosavic, N.; Bezbradica, D. Immobilization of Candida antarctica lipase B onto Purolite® MN102 and its application in solvent-free and organic media esterification. Bioprocess Biosyst. Eng. 2017, 40, 23–34. [Google Scholar] [CrossRef]
- Li, D.M.; Wang, W.F.; Liu, P.Z.; Xu, L.; Faiza, M.; Yang, B.; Wang, L.Y.; Lan, D.M.; Wang, Y.H. Immobilization of Candida antarctica Lipase B Onto ECR1030 Resin and its Application in the Synthesis of n-3 PUFA-Rich Triacylglycerols. Eur. J. Lipid Sci. Technol. 2017, 119, 1700266–1700275. [Google Scholar] [CrossRef]
- Gokalp, N.; Ulker, C.; Guvenilir, Y. Synthesis of Polycaprolactone via Ring Opening Polymerization Catalyzed by Candida Antarctica Lipase B Immobilized onto an Amorphous Silica Support. J. Polym. Mater. 2016, 33, 87–100. [Google Scholar]
- Cunha, A.G.; Besteti, M.D.; Manoel, E.A.; da Silva, A.A.T.; Almeida, R.V.; Simas, A.B.C.; Fernandez-Lafuente, R.; Pinto, J.C.; Freire, D.M.G. Preparation of core-shell polymer supports to immobilize lipase B from Candida antarctica Effect of the support nature on catalytic properties. J. Mol. Catal. B Enzym. 2014, 100, 59–67. [Google Scholar] [CrossRef]
- Gan, Z.H.; Zhang, T.; Liu, Y.C.; Wu, D.C. Temperature-Triggered Enzyme Immobilization and Release Based on Cross-Linked Gelatin Nanoparticles. PLoS ONE 2012, 7, e47154. [Google Scholar] [CrossRef]
- da Rocha, T.N.; Carballares, D.; Guimaraes, J.R.; Rocha-Martin, J.; Tardioli, P.W.; Goncalves, L.R.B.; Fernandez-Lafuente, R. Determination of immobilized lipase stability depends on the substrate and activity determination condition: Stress inactivations and optimal temperature as biocatalysts stability indicators. Sustain. Chem. Pharm. 2022, 29, 100823–100836. [Google Scholar] [CrossRef]
- Aghabeigi, F.; Nikkhah, H.; Zilouei, H.; Bazarganipour, M. Immobilization of lipase on the graphene oxides magnetized with NiFe2O4 nanoparticles for biodiesel production from microalgae lipids. Process. Biochem. 2023, 126, 171–185. [Google Scholar] [CrossRef]
- Aghaei, H.; Yasinian, A.; Taghizadeh, A. Covalent immobilization of lipase from Candida rugosa on epoxy-activated cloisite 30B as a new heterofunctional carrier and its application in the synthesis of banana flavor and production of biodiesel. Int. J. Biol. Macromol. 2021, 178, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Abd Halin, N.I.; Al-Khatib, M.F.R.; Salleh, H.M.; Nasef, M.M. Immobilization of Candida rugosa Lipase on Aminated Polyvinyl Benzyl Chloride-Grafted Nylon-6 Microfibers. Bull. Chem. React. Eng. Catal. 2019, 14, 369–379. [Google Scholar] [CrossRef]
- Babaei, M.; Karimi, A.; Hejazi, M.A. Use of mesoporous MnO2 as a support for immobilization of lipase from Candida rugosa. Chem. Ind. Chem. Eng. Q. 2014, 20, 371–378. [Google Scholar] [CrossRef]
- Siódmiak, T.; Siódmiak, J.; Mastalerz, R.; Kocot, N.; Dulęba, J.; Haraldsson, G.G.; Wątróbska-Świetlikowska, D.; Marszałł, M.P. Climatic Chamber Stability Tests of Lipase-Catalytic Octyl-Sepharose Systems. Catalysts 2023, 13, 501. [Google Scholar] [CrossRef]
- Barbosa, J.M.P.; Souza, R.L.; de Melo, C.M.; Fricks, A.T.; Soares, C.M.F.; Lima, A.S. Biochemical characterisation of lipase from a new strain of Bacillus sp. ITP-001. Quim. Nova 2012, 35, 1173–1178. [Google Scholar] [CrossRef]
- Qin, S.; Zhao, Y.Y.; Wu, B.; He, B.F. A Calcium-Ion-Stabilized Lipase from Pseudomonas stutzeri ZS04 and its Application in Resolution of Chiral Aryl Alcohols. Appl. Biochem. Biotechnol. 2016, 180, 1456–1466. [Google Scholar] [CrossRef]
- de Almeida, A.F.; Terrasan, C.R.F.; Terrone, C.C.; Tauk-Tornisielo, S.M.; Carmona, E.C. Biochemical properties of free and immobilized Candida viswanathii lipase on octyl-agarose support: Hydrolysis of triacylglycerol and soy lecithin. Process. Biochem. 2018, 65, 71–80. [Google Scholar] [CrossRef]
- Chiaradia, V.; Soares, N.S.; Valerio, A.; de Oliveira, D.; Araujo, P.H.H.; Sayer, C. Immobilization of Candida antarctica Lipase B on Magnetic Poly(Urea-Urethane) Nanoparticles. Appl. Biochem. Biotechnol. 2016, 180, 558–575. [Google Scholar] [CrossRef]
- Vetrano, A.; Gabriele, F.; Germani, R.; Spreti, N. Characterization of lipase from Candida rugosa entrapped in alginate beads to enhance its thermal stability and recyclability. New J. Chem. 2022, 46, 10037–10047. [Google Scholar] [CrossRef]
- Yu, D.Y.; Li, Z.Y.; Zhou, X.A.; Wang, W.N.; Wang, L.Q.; Liu, T.Y.; Du, J. Study on the modification of magnetic graphene oxide and the effect of immobilized lipase. Int. J. Biol. Macromol. 2022, 216, 498–509. [Google Scholar] [CrossRef] [PubMed]
- Ameri, A.; Shakibaie, M.; Khoobi, M.; Faramarzi, M.A.; Forootanfar, H. Immobilization of thermoalkalophilic lipase from Bacillus atrophaeus FSHM2 on amine-modified graphene oxide nanostructures: Statistical optimization and its application for pentyl valerate synthesis. Appl. Biochem. Biotechnol. 2020, 191, 579–604. [Google Scholar] [CrossRef] [PubMed]
- Pazol, J.; Weiss, T.M.; Martínez, C.D.; Quesada, O.; Nicolau, E. The influence of calcium ions (Ca2+) on the enzymatic hydrolysis of lipopolysaccharide aggregates to liberate free fatty acids (FFA) in aqueous solution. JCIS Open 2022, 7, 100058–100067. [Google Scholar] [CrossRef]
- Yu, J.Y.; Wang, C.H.; Wang, A.M.; Li, N.N.; Chen, X.X.; Pei, X.L.; Zhang, P.F.; Wu, S.G. Dual-cycle immobilization to reuse both enzyme and support by reblossoming enzyme-inorganic hybrid nanoflowers. Rsc. Adv. 2018, 8, 16088–16094. [Google Scholar] [CrossRef] [PubMed]
- Arana-Pena, S.; Lokha, Y.; Fernandez-Lafuente, R. Immobilization of Eversa Lipase on Octyl Agarose Beads and Preliminary Characterization of Stability and Activity Features. Catalysts 2018, 8, 511. [Google Scholar] [CrossRef]
- Lokha, Y.; Arana-Pena, S.; Rios, N.S.; Mendez-Sanchez, C.; Goncalves, L.R.B.; Lopez-Gallego, F.; Fernandez-Lafuente, R. Modulating the properties of the lipase from Thermomyces lanuginosus immobilized on octyl agarose beads by altering the immobilization conditions. Enzym. Microb. Technol. 2020, 133, 109461. [Google Scholar] [CrossRef]
- Fernandez-Lopez, L.; Rueda, N.; Bartolome-Cabrero, R.; Rodriguez, M.D.; Albuquerque, T.L.; dos Santos, J.C.S.; Barbosa, O.; Fernandez-Lafuente, R. Improved immobilization and stabilization of lipase from Rhizomucor miehei on octyl-glyoxyl agarose beads by using CaCl2. Process. Biochem. 2016, 51, 48–52. [Google Scholar] [CrossRef]
- Vesoloski, J.F.; Todero, A.S.; Macieski, R.J.; Pereira, F.D.; Dallago, R.M.; Mignoni, M.L. Immobilization of Lipase from Candida antarctica B (CALB) by Sol-Gel Technique Using Rice Husk Ash as Silic Source and Ionic Liquid as Additive. Appl. Biochem. Biotechnol. 2022, 194, 6270–6286. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Lipase B from Candida antarctica | References | Lipase from Candida rugosa | References | |
|---|---|---|---|---|
| Molecular structure |  | [8] |  | [9] |
| Active site | Serine-105, Histidine-224, Aspartate-187 | [10] | Serine-209, Histidine-449, Glutamate-341 | [9,11] |
| The presence of a lid | Debated | [12] | Presence | [9,11,13] |
| Isoelectric point or points range | pH 6.0 | [14] | pH 4.0–5.5 | [15] |
| Optimum temperature | 10–60 °C | [16] | 30–50 °C | [17] |
| Optimum pH | 6.0–9.0 | [16] | 7.0 | [17] |
| Isoforms | 1 | [18] | 5 | [19] |
| Enantiopreference | R-enantiomers, S-enantiomers | [20] | S-enantiomers | [21] |
| Carbon chain length preference | Short-length | [6] | From short to long | [22] |
| First Step | Second Step | Results | |||||
|---|---|---|---|---|---|---|---|
| Stirring (5 min) in Ice Bath | Stirring (5 min) at Room Temperature | Incubation 14 h at 4 °C | Incubation 14 h at Room Temperature (22 °C) | Incubation 14 h at 37 °C | Stirring during 14 h of Incubation | Relative Activity (%) | Ie (%) |
| + | + | 93.20 ± 3.02 | 55.20 ± 1.00 | ||||
| + | + | 100.00 ± 2.25 | 61.90 ± 0.74 | ||||
| + | + | 90.70 ± 2.55 | 52.30 ± 0.84 | ||||
| + | + | + | 90.70 ± 1.22 | 51.50 ± 0.40 | |||
| + | + | 95.40 ± 2.41 | 57.70 ± 0.79 | ||||
| + | + | + | 93.20 ± 0.75 | 54.90 ± 0.25 | |||
| Alternative protocol: Incubation time: 30 min, 22 °C, constant stirring | 38.70 ± 2.53 | 21.15 ± 0.83 | |||||
| Alternative protocol: Incubation time: 1 h, 22 °C, constant stirring | 56.70 ± 2.11 | 37.41 ± 0.69 | |||||
| (a) | |||||||||
| Storage Conditions | Immobilized CALB | ||||||||
| Citrate Buffer (pH 4.0, 500 mM) | Water | Ca2+ | Vis (400–800 nm) | T [65 °C] | T [4 °C] | DCP | CALB Residual Activity [%] | Activity CALB [U] | Activity CALB [U/g Support] |
| + | + | 340.9 ± 30.2 | 3.30 ± 0.29 | 66.04 ± 5.85 | |||||
| + | + | + | 564.5 ± 21.6 | 5.47 ± 0.21 | 109.38 ± 4.19 | ||||
| + | + | + | 82.8 ± 17.9 | 0.80 ± 0.17 | 16.04 ± 3.47 | ||||
| + | + | 220.4 ± 26.3 | 2.14 ± 0.25 | 42.71 ± 5.09 | |||||
| + | + | + | 409.7 ± 13.1 | 3.97 ± 0.13 | 79.38 ± 2.55 | ||||
| + | + | + | 237.6 ± 30.2 | 2.30 ± 0.29 | 46.04 ± 5.85 | ||||
| (b) | |||||||||
| Storage Conditions | Immobilized CRL-OF | ||||||||
| Citrate Buffer (pH 4.0, 500 mM) | Water | Ca2+ | Vis (400–800 nm) | T [65 °C] | T [4 °C] | DCP | CRL-OF Residual Activity [%] | Activity CRL-OF [U] | Activity CRL-OF [U/g Support] |
| + | + | 4.2 ± 0.5 | 1.47 ± 0.19 | 29.38 ± 3.85 | |||||
| + | + | + | 3.8 ± 0.9 | 1.30 ± 0.33 | 26.04 ± 6.67 | ||||
| + | + | + | 0.4 ± 0.3 | 0.14 ± 0.10 | 2.71 ± 1.92 | ||||
| + | + | 89.9 ± 3.4 | 31.14 ± 1.83 | 622.71 ± 36.67 | |||||
| + | + | + | 2.3 ± 0.7 | 0.80 ± 0.25 | 16.04 ± 5.09 | ||||
| + | + | + | 2.3 ± 0.3 | 0.80 ± 0.10 | 16.04 ± 1.92 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siódmiak, T.; Dulęba, J.; Haraldsson, G.G.; Siódmiak, J.; Marszałł, M.P. The Studies of Sepharose-Immobilized Lipases: Combining Techniques for the Enhancement of Activity and Thermal Stability. Catalysts 2023, 13, 887. https://doi.org/10.3390/catal13050887
Siódmiak T, Dulęba J, Haraldsson GG, Siódmiak J, Marszałł MP. The Studies of Sepharose-Immobilized Lipases: Combining Techniques for the Enhancement of Activity and Thermal Stability. Catalysts. 2023; 13(5):887. https://doi.org/10.3390/catal13050887
Chicago/Turabian StyleSiódmiak, Tomasz, Jacek Dulęba, Gudmundur G. Haraldsson, Joanna Siódmiak, and Michał Piotr Marszałł. 2023. "The Studies of Sepharose-Immobilized Lipases: Combining Techniques for the Enhancement of Activity and Thermal Stability" Catalysts 13, no. 5: 887. https://doi.org/10.3390/catal13050887