
Climatic Chamber Stability Tests of Lipase-Catalytic Octyl-Sepharose Systems
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Lipase CALB Immobilization Protocols
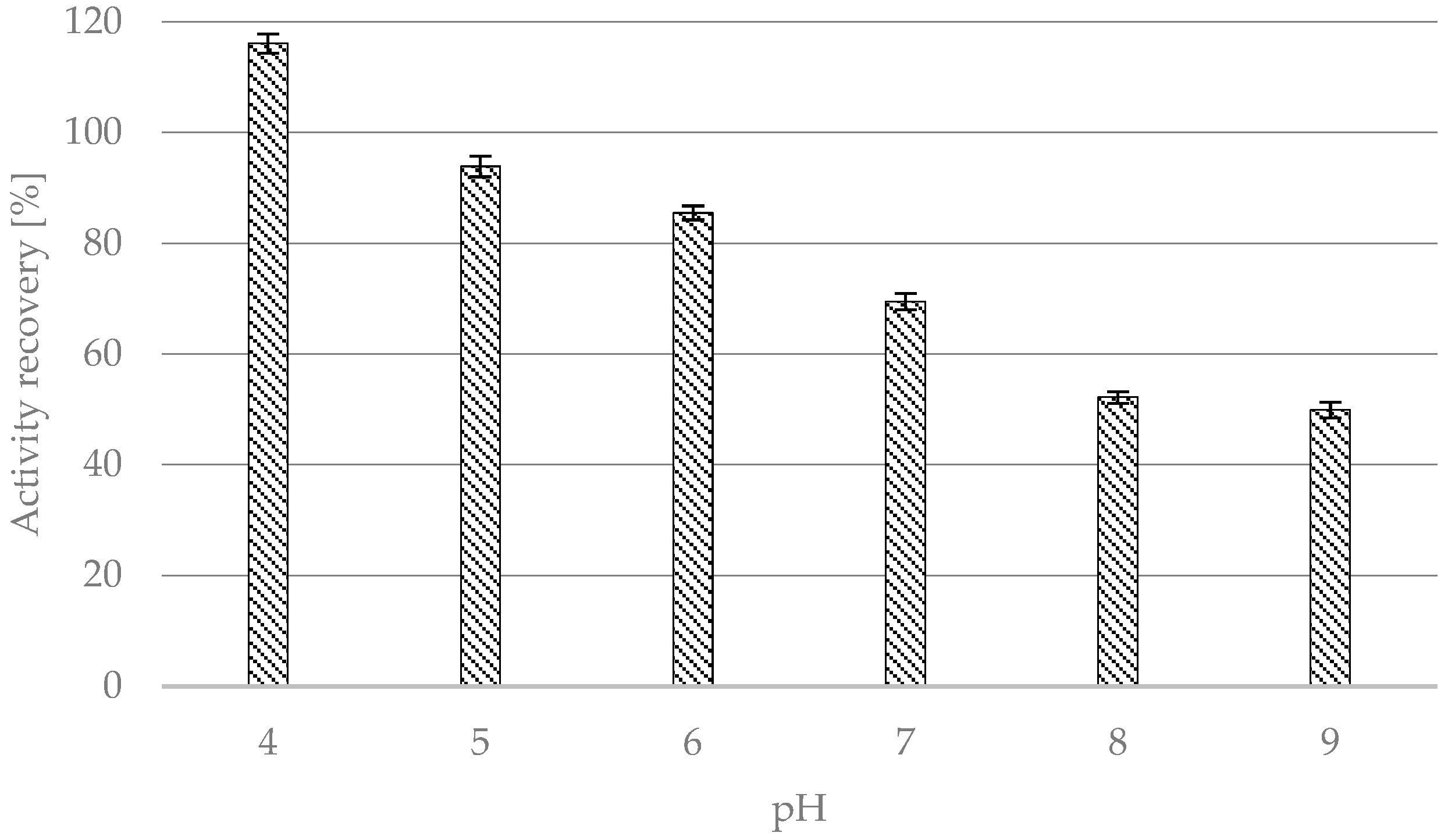
2.1.1. Effect of Buffer pH
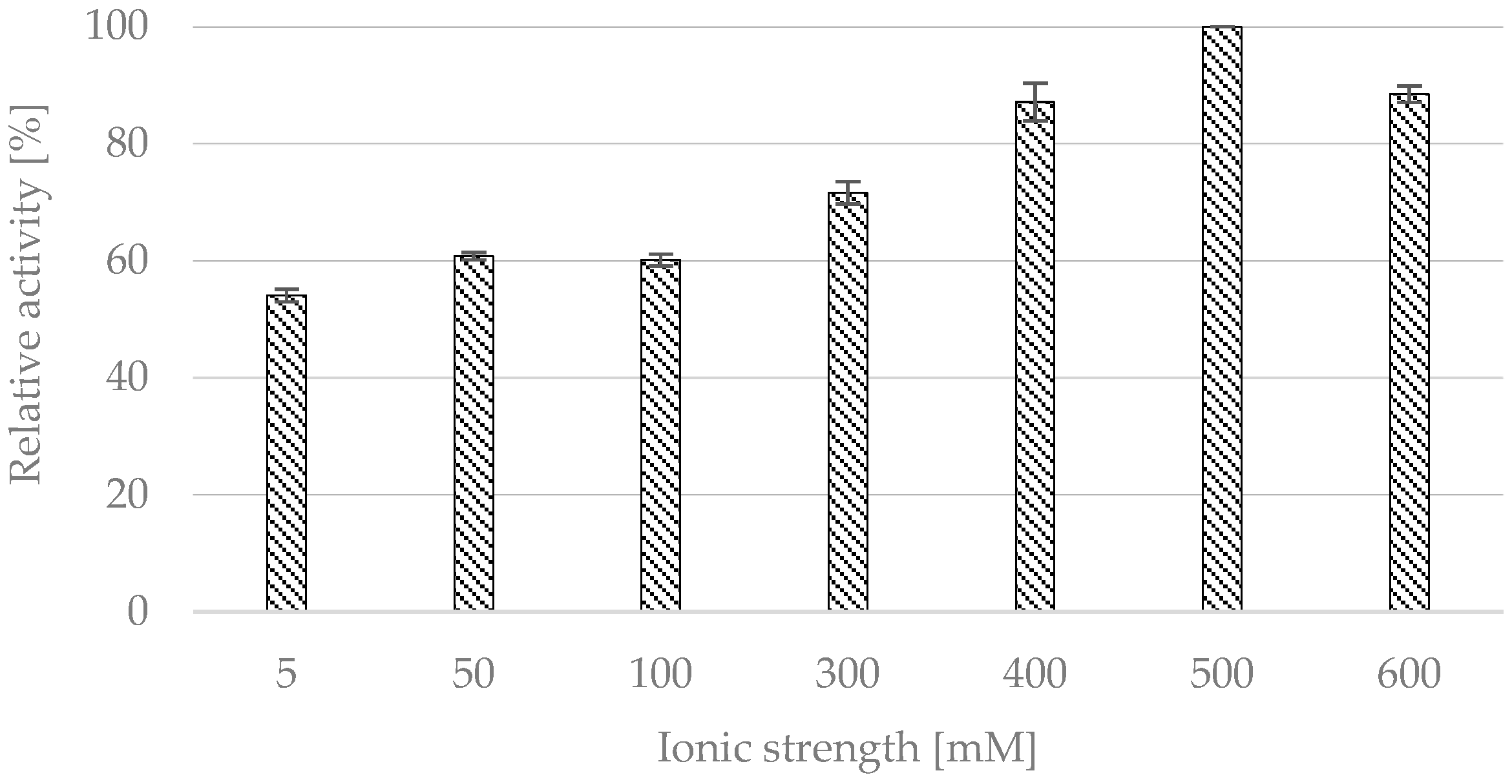
2.1.2. Effect of Ionic Strength
2.2. Lipase from Candida Rugosa (CRL OF) Immobilization Protocols
2.2.1. Effect of pH
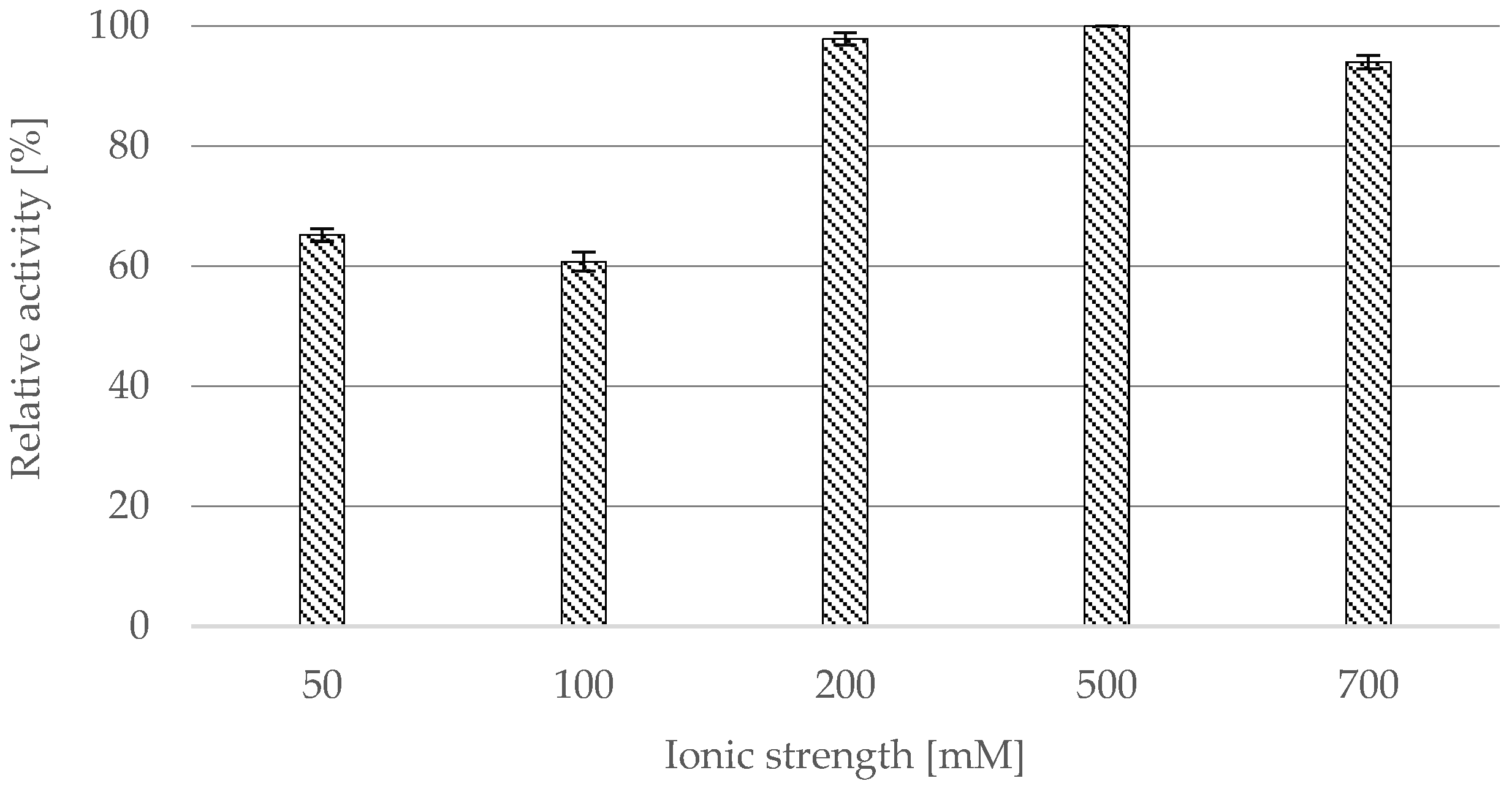
2.2.2. Effect of Ionic Strength
2.3. Climatic Chamber Storage Stability Tests of CALB and CRL-OF
3. Materials and Methods
3.1. Materials
3.2. Preparation of Octyl Sepharose CL-4B
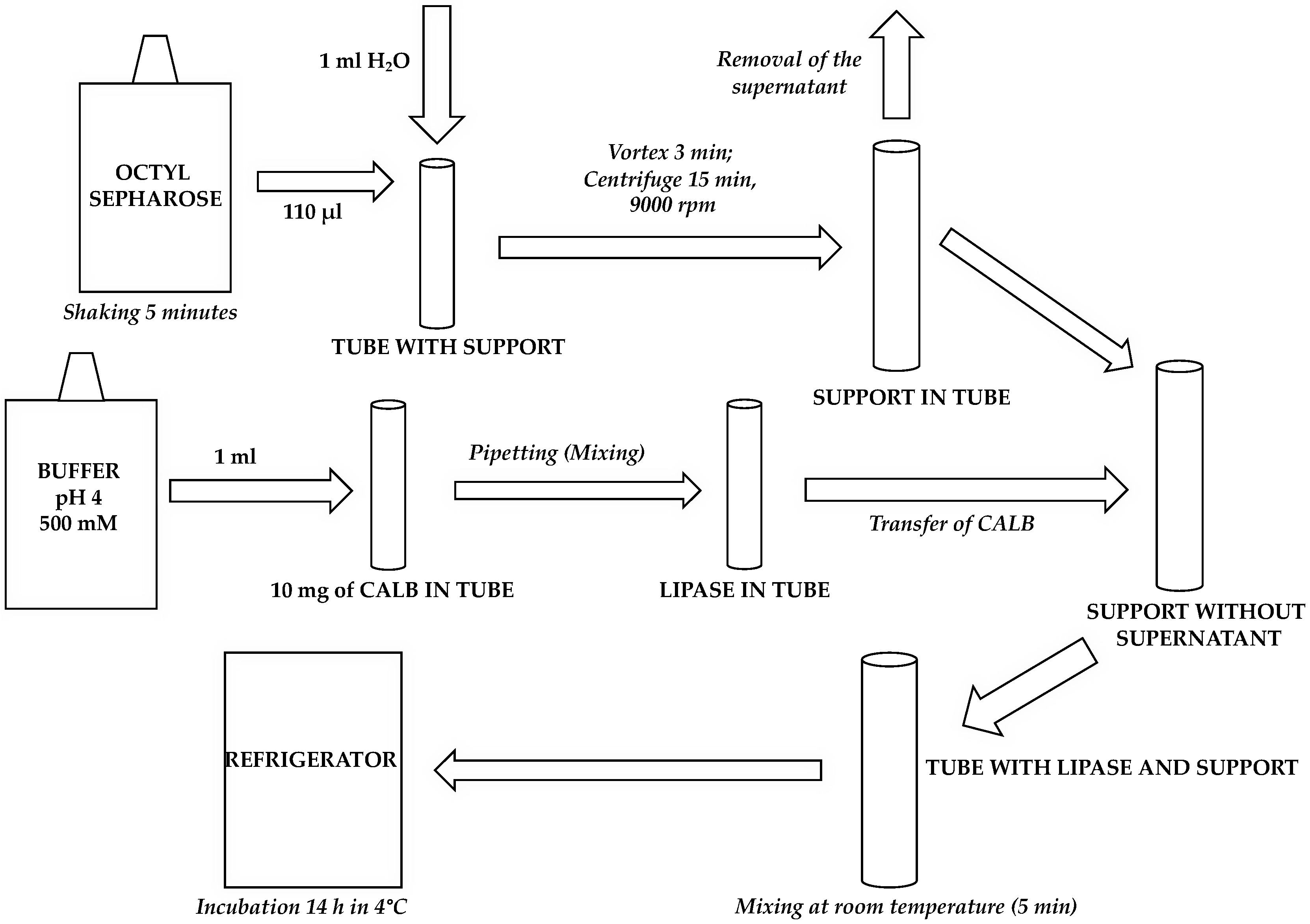
3.3. Immobilization of CALB onto Octyl-Sepharose CL-4B
- pH: CALB was suspended in 1.0 mL of an appropriate 100 mM buffer with pH 4, 5, and 6 (citrate buffer), 7, 8 (phosphate buffer), and 9 (Trizma Base buffer).
- Ionic strength: CALB was suspended in 1.0 mL of citrate buffer (pH 4) with ionic strength of: 5, 50, 100, 300, 400, 500, and 600 mM.
3.4. Immobilization of CRL-OF onto Octyl-Sepharose CL-4B
- pH: CRL-OF was suspended in 1.0 mL of an appropriate 100 mM buffer with pH 4 (citrate buffer), 7 (phosphate buffer), and 9 (Trizma Base buffer).
- Ionic strength: CRL-OF was suspended in 1.0 mL of citrate buffer (pH 4) with ionic strength of: 50, 100, 200, 500, and 700 mM.
3.5. Determination of the Amount of Immobilized CALB by Bradford Method
3.6. Lipolytic Activity of CALB and CRL-OF
3.7. Climatic Chamber Storage Stability Tests of CALB and CRL-OF in Dry Form
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alcantara, A.R.; de Maria, P.D.; Littlechild, J.A.; Schurmann, M.; Sheldon, R.A.; Wohlgemuth, R. Biocatalysis as Key to Sustainable Industrial Chemistry. Chemsuschem 2022, 15, e202102709. [Google Scholar] [PubMed]
- Sheldon, R.A.; Woodley, J.M. Role of Biocatalysis in Sustainable Chemistry. Chem. Rev. 2018, 118, 801–838. [Google Scholar] [CrossRef]
- Sheldon, R.A.; van Pelt, S. Enzyme immobilisation in biocatalysis: Why, what and how. Chem. Soc. Rev. 2013, 42, 6223–6235. [Google Scholar] [CrossRef] [Green Version]
- Anastas, P.; Eghbali, N. Green Chemistry: Principles and Practice. Chem. Soc. Rev. 2010, 39, 301–312. [Google Scholar] [CrossRef]
- Solano, D.M.; Hoyos, P.; Hernaiz, M.J.; Alcantara, A.R.; Sanchez-Montero, J.M. Industrial biotransformations in the synthesis of building blocks leading to enantiopure drugs. Bioresour. Technol. 2012, 115, 196–207. [Google Scholar] [CrossRef]
- Magnusson, C.D.; Haraldsson, G.G. Chemoenzymatic synthesis of symmetrically structured triacylglycerols possessing short-chain fatty acids. Tetrahedron 2010, 66, 2728–2731. [Google Scholar] [CrossRef]
- Magnusson, C.D.; Haraldsson, G.G. Activation of n-3 polyunsaturated fatty acids as oxime esters: A novel approach for their exclusive incorporation into the primary alcoholic positions of the glycerol moiety by lipase. Chem. Phys. Lipids 2012, 165, 712–720. [Google Scholar] [CrossRef] [PubMed]
- Gudmundsson, H.G.; Linderborg, K.M.; Kallio, H.; Yang, B.R.; Haraldsson, G.G. Synthesis of enantiopure ABC-type triacylglycerols. Tetrahedron 2020, 76, 130813–130825. [Google Scholar] [CrossRef]
- Manoel, E.A.; dos Santos, J.C.S.; Freire, D.M.G.; Rueda, N.; Fernandez-Lafuente, R. Immobilization of lipases on hydrophobic supports involves the open form of the enzyme. Enzym. Microb. Technol. 2015, 71, 53–57. [Google Scholar] [CrossRef]
- Khan, F.I.; Lan, D.; Durrani, R.; Huan, W.; Zhao, Z.; Wang, Y. The Lid Domain in Lipases: Structural and Functional Determinant of enzymatic Properties. Front. Bioeng. Biotechnol. 2017, 5, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, D.A.; Tonetto, G.M.; Ferreira, M.L. Burkholderia cepacia lipase: A versatile catalyst in synthesis reactions. Biotechnol. Bioeng. 2018, 115, 6–24. [Google Scholar] [CrossRef]
- Sun, T.; Dong, Z.; Wang, J.; Huang, F.H.; Zheng, M.M. Ultrasound-Assisted Interfacial Immobilization of Lipase on Hollow Mesoporous Silica Spheres in a Pickering Emulsion System: A Hyperactive and Sustainable Biocatalyst. ACS Sustain. Chem. Eng. 2020, 8, 17280–17290. [Google Scholar] [CrossRef]
- Wang, S.; Xu, Y.; Yu, X.W. A phenylalanine dynamic switch controls the interfacial activation of Rhizopus chinensis lipase. Int. J. Biol. Macromol. 2021, 173, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, C.; Ferreira, M.L.; Barbosa, O.; dos Santos, J.C.S.; Rodrigues, R.C.; Berenguer-Murcia, A.; Briand, L.E.; Fernandez-Lafuente, R. Novozym 435: The “perfect” lipase immobilized biocatalyst? Catal. Sci. Technol. 2019, 9, 2380–2420. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.H.; Wang, Z.X.; Zhuang, W.; Rabiee, H.; Zhu, C.J.; Deng, J.W.; Ge, L.; Ying, H.J. Amphiphilic Nanointerface: Inducing the Interfacial Activation for Lipase. ACS Appl. Mater. Interfaces 2022, 14, 39622–39636. [Google Scholar] [CrossRef]
- Zisis, T.; Freddolino, P.L.; Turunen, P.; van Teeseing, M.C.F.; Rowan, A.E.; Blank, K.G. Interfacial Activation of Candida antarctica Lipase B: Combined Evidence from Experiment and Simulation. Biochemistry 2015, 54, 5969–5979. [Google Scholar] [CrossRef] [Green Version]
- Siódmiak, T.; Haraldsson, G.G.; Dulęba, J.; Ziegler-Borowska, M.; Siódmiak, J.; Marszałł, M.P. Evaluation of Designed Immobilized Catalytic Systems: Activity Enhancement of Lipase B from Candida antarctica. Catalysts 2020, 10, 876. [Google Scholar] [CrossRef]
- Marszałł, M.P.; Siódmiak, T. Immobilization of Candida rugosa lipase onto magnetic beads for kinetic resolution of (R,S)-ibuprofen. Catal. Commun. 2012, 24, 80–84. [Google Scholar] [CrossRef]
- Silvestrini, L.; Cianci, M. Principles of lipid-enzyme interactions in the limbus region of the catalytic site of Candida antarctica Lipase B. Int. J. Biol. Macromol. 2020, 158, 358–363. [Google Scholar] [CrossRef]
- Rabbani, G.; Ahmad, E.; Khan, M.V.; Ashraf, M.T.; Bhat, R.; Khan, R.H. Impact of structural stability of cold adapted Candida antarctica lipase B (CaLB): In relation to pH, chemical and thermal denaturation. RSC Adv. 2015, 5, 20115–20131. [Google Scholar] [CrossRef]
- Miletic, N.; Abetz, V.; Ebert, K.; Loos, K. Immobilization of Candida antarctica lipase B on Polystyrene Nanoparticles. Macromol. Rapid. Commun. 2010, 31, 71–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahedi, M.; Habibi, Z.; Yousefi, M.; Brask, J.; Mohammadi, M. Improvement of biodiesel production from palm oil by co-immobilization of Thermomyces lanuginosa lipase and Candida antarctica lipase B: Optimization using response surface methodology. Int. J. Biol. Macromol. 2021, 170, 490–502. [Google Scholar] [CrossRef]
- Siódmiak, T.; Mangelings, D.; Heyden, Y.V.; Ziegler-Borowska, M.; Marszałł, M.P. High Enantioselective Novozym 435-Catalyzed Esterification of (R,S)- Flurbiprofen Monitored with a Chiral Stationary Phase. Appl. Biochem. Biotechnol. 2015, 175, 2769–2785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sikora, A.; Siódmiak, T.; Marszałł, M.P. Kinetic Resolution of Profens by Enantioselective Esterification Catalyzed by Candida antarctica and Candida rugosa Lipases. Chirality 2014, 26, 663–669. [Google Scholar] [CrossRef] [PubMed]
- Ghanem, A.; Aboul-Enein, H.Y. Application of lipases in kinetic resolution of racemates. Chirality 2005, 17, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Francolini, I.; Taresco, V.; Martinelli, A.; Piozzi, A. Enhanced performance of Candida rugosa lipase immobilized onto alkyl chain modified-magnetic nanocomposites. Enzym. Microb. Technol. 2020, 132, 109439–109447. [Google Scholar] [CrossRef]
- Fernandes, K.V.; Cavalcanti, E.D.C.; Cipolatti, E.P.; Aguieiras, E.C.G.; Pinto, M.C.C.; Tavares, F.A.; da Silva, P.R.; Fernandez-Lafuente, R.; Arana-Pena, S.; Pinto, J.C.; et al. Enzymatic synthesis of biolubricants from by-product of soybean oil processing catalyzed by different biocatalysts of Candida rugosa lipase. Catal. Today 2021, 362, 122–129. [Google Scholar] [CrossRef]
- Elias, N.; Wahab, R.A.; Chandren, S.; Lau, W.J. Performance of Candida rugosa lipase supported on nanocellulose-silica-reinforced polyethersulfone membrane for the synthesis of pentyl valerate: Kinetic, thermodynamic and regenerability studies. Mol. Catal. 2021, 514, 111852. [Google Scholar] [CrossRef]
- Menden, A.; Crynen, S.; Mathura, V.; Paris, D.; Crawford, F.; Mullan, M.; Ait-Ghezala, G. Novel, natural allosteric inhibitors and enhancers of Candida rugosa lipase activity. Bioorg. Chem. 2021, 109, 104732–104738. [Google Scholar] [CrossRef]
- Binhayeeding, N.; Yunu, T.; Pichid, N.; Klomklao, S.; Sangkharak, K. Immobilisation of Candida rugosa lipase on polyhydroxybutyrate via a combination of adsorption and cross-linking agents to enhance acylglycerol production. Process Biochem. 2020, 95, 174–185. [Google Scholar] [CrossRef]
- Vakhlu, J.; Kour, A. Yeast lipases: Enzyme purification, biochemical properties and gene cloning. Electron. J. Biotechnol. 2006, 9, 69–85. [Google Scholar] [CrossRef] [Green Version]
- Dulęba, J.; Siódmiak, T.; Marszałł, M.P. The influence of substrate systems on the enantioselective and lipolytic activity of immobilized Amano PS from Burkholderia cepacia lipase (APS-BCL). Process Biochem. 2022, 120, 126–137. [Google Scholar] [CrossRef]
- Mokhtar, N.F.R.; Rahman, A.B.D.; Noor, N.D.M.; Shariff, F.M.; Ali, M.S.M. The Immobilization of Lipases on Porous Support by Adsorption and Hydrophobic Interaction Method. Catalysts 2020, 10, 744. [Google Scholar] [CrossRef]
- Dulęba, J.; Siódmiak, T.; Marszałł, M.P. Amano Lipase PS from Burkholderia cepacia—Evaluation of the Effect of Substrates and Reaction Media on the Catalytic Activity. Curr. Org. Chem. 2020, 24, 798–807. [Google Scholar] [CrossRef]
- Thangaraj, B.; Solomon, P.R. Immobilization of Lipases—A Review. Part I: Enzyme Immobilization. Chembioeng. Rev. 2019, 6, 157–166. [Google Scholar] [CrossRef]
- Bolivar, J.M.; Woodley, J.M.; Fernandez-Lafuente, R. Is enzyme immobilization a mature discipline? Some critical considerations to capitalize on the benefits of immobilization. Chem. Soc. Rev. 2022, 51, 6251–6290. [Google Scholar] [CrossRef]
- Rodrigues, R.C.; Berenguer-Murcia, A.; Carballares, D.; Morellon-Sterling, R.; Fernandez-Lafuente, R. Stabilization of enzymes via immobilization: Multipoint covalent attachment and other stabilization strategies. Biotechnol. Adv. 2021, 52, 107821–107857. [Google Scholar] [CrossRef]
- Mohamad, N.R.; Marzuki, N.H.C.; Buang, N.A.; Huyop, F.; Wahab, R.A. An overview of technologies for immobilization of enzymes and surface analysis techniques for immobilized enzymes. Biotechnol. Biotechnol. Equip. 2015, 29, 205–220. [Google Scholar] [CrossRef]
- Fernandez-Lorente, G.; Palomo, J.M.; Cabrera, Z.; Guisan, J.M.; Fernandez-Lafuente, R. Specificity enhancement towards hydrophobic substrates by immobilization of lipases by interfacial activation on hydrophobic supports. Enzym. Microb. Technol. 2007, 41, 565–569. [Google Scholar] [CrossRef]
- Thangaraj, B.; Solomon, P.R. Immobilization of Lipases—A Review. Part II: Carrier Materials. Chembioeng. Rev. 2019, 6, 167–194. [Google Scholar] [CrossRef]
- Dos Santos, J.C.S.; Barbosa, O.; Ortiz, C.; Berenguer-Murcia, A.; Rodrigues, R.C.; Fernandez-Lafuente, R. Importance of the Support Properties for Immobilization or Purification of Enzymes. ChemCatChem 2015, 7, 2413–2432. [Google Scholar] [CrossRef] [Green Version]
- Sirisha, V.L.; Jain, A. Enzyme Immobilization: An Overview on Methods, Support Material, and Applications of Immobilized Enzymes. Adv. Food Nutr. Res. 2016, 79, 179–211. [Google Scholar] [PubMed]
- Ismail, A.R.; Baek, K.H. Lipase immobilization with support materials, preparation techniques, and applications: Present and future aspects. Int. J. Biol. Macromol. 2020, 163, 1624–1639. [Google Scholar] [CrossRef]
- Rafiee, F.; Rezaee, M. Different strategies for the lipase immobilization on the chitosan based supports and their applications. Int. J. Biol. Macromol. 2021, 179, 170–195. [Google Scholar] [CrossRef]
- Zucca, P.; Fernandez-Lafuente, R.; Sanjust, E. Agarose and Its Derivatives as Supports for Enzyme Immobilization. Molecules 2016, 21, 1577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alnoch, R.C.; dos Santos, L.A.; de Almeida, J.M.; Krieger, N.; Mateo, C. Recent Trends in Biomaterials for Immobilization of Lipases for Application in Non-Conventional Media. Catalysts 2020, 10, 697. [Google Scholar] [CrossRef]
- Rios, N.S.; Neto, D.M.A.; dos Santos, J.C.S.; Fechine, P.B.A.; Fernandez-Lafuente, R.; Goncalves, L.R.B. Comparison of the immobilization of lipase from Pseudomonas fluorescens on divinylsulfone or p-benzoquinone activated support. Int. J. Biol. Macromol. 2019, 134, 936–945. [Google Scholar] [CrossRef]
- Arana-Pena, S.; Lokha, Y.; Fernandez-Lafuente, R. Immobilization on octyl-agarose beads and some catalytic features of commercial preparations of lipase a from Candida antarctica (Novocor ADL): Comparison with immobilized lipase B from Candida antarctica. Biotechnol. Prog. 2019, 35, e2735. [Google Scholar] [CrossRef] [Green Version]
- Bramer, C.; Schreiber, S.; Scheper, T.; Beutel, S. Continuous purification of Candida antarctica lipase B using 3-membrane adsorber periodic counter-current chromatography. Eng. Life Sci. 2018, 18, 414–424. [Google Scholar] [CrossRef] [Green Version]
- Subinya, M.; Steudle, A.K.; Jurkowski, T.P. Stubenrauch, C. Conformation and activity of lipase B from Candida antarctica in bicontinuous microemulsions. Colloids Surf. B 2015, 131, 108–114. [Google Scholar] [CrossRef]
- Li, D.M.; Wang, W.F.; Liu, P.Z.; Xu, L.; Faiza, M.; Yang, B.; Wang, L.Y.; Lan, D.M.; Wang, Y.H. Immobilization of Candida antarctica Lipase B Onto ECR1030 Resin and its Application in the Synthesis of n-3 PUFA-Rich Triacylglycerols. Eur. J. Lipid Sci. Technol. 2017, 119, 1700266–1700275. [Google Scholar] [CrossRef]
- Brigida, A.I.S.; Pinheiro, A.D.T.; Ferreira, A.L.O.; Goncalves, L.R.B. Immobilization of Candida antarctica lipase B by adsorption to green coconut fiber. Appl. Biochem. Biotechnol. 2008, 146, 173–187. [Google Scholar] [CrossRef] [PubMed]
- Coskun, G.; Ciplak, Z.; Yildiz, N.; Mehmetoglu, U. Immobilization of Candida antarctica Lipase on Nanomaterials and Investigation of the Enzyme Activity and Enantioselectivity. Appl. Biochem. Biotechnol. 2021, 193, 430–445. [Google Scholar] [CrossRef] [PubMed]
- Tanaskovic, S.J.; Jokic, B.; Grbavcic, S.; Drvenica, I.; Prlainovic, N.; Lukovic, N.; Knezevic-Jugovic, Z. Immobilization of Candida antarctica lipase B on kaolin and its application in synthesis of lipophilic antioxidants. Appl. Clay Sci. 2017, 135, 103–111. [Google Scholar] [CrossRef]
- Arana-Pena, S.; Rios, N.S.; Carballares, D.; Goncalves, L.R.B.; Fernandez-Lafuente, R. Immobilization of lipases via interfacial activation on hydrophobic supports: Production of biocatalysts libraries by altering the immobilization conditions. Catal. Today 2021, 362, 130–140. [Google Scholar] [CrossRef]
- Cespugli, M.; Lotteria, S.; Navarini, L.; Lonzarich, V.; Del Terra, L.; Vita, F.; Zweyer, M.; Baldini, G.; Ferrario, V.; Ebert, C.; et al. Rice Husk as an Inexpensive Renewable Immobilization Carrier for Biocatalysts Employed in the Food, Cosmetic and Polymer Sectors. Catalysts 2018, 8, 471. [Google Scholar] [CrossRef] [Green Version]
- Mihailovic, M.; Stojanovic, M.; Banjanac, K.; Carevic, M.; Prlainovic, N.; Milosavic, N.; Bezbradica, D. Immobilization of lipase on epoxy-activated Purolite((R)) A109 and its post-immobilization stabilization. Process Biochem. 2014, 49, 637–646. [Google Scholar] [CrossRef]
- Cea, M.; Gonzalez, M.E.; Abarzua, M.; Navia, R. Enzymatic esterification of oleic acid by Candida rugosa lipase immobilized onto biochar. J. Environ. Manag. 2019, 242, 171–177. [Google Scholar] [CrossRef]
- De Melo, R.R.; Alnoch, R.C.; Vilela, A.F.L.; de Souza, E.M.; Krieger, N.; Ruller, R.; Sato, H.H.; Mateo, C. New Heterofunctional Supports Based on Glutaraldehyde-Activation: A Tool for Enzyme Immobilization at Neutral pH. Molecules 2017, 22, 1088. [Google Scholar] [CrossRef] [Green Version]
- Siódmiak, T.; Dulęba, J.; Kocot, N.; Watróbska-Świetlikowska, D.; Marszałł, M.P. The High ‘Lipolytic Jump’ of Immobilized Amano A Lipase from Aspergillus niger in Developed ‘ESS Catalytic Triangles’ Containing Natural Origin Substrates. Catalysts 2022, 12, 853. [Google Scholar] [CrossRef]
- Amjad, M.; Akhtar, S.S.; Yang, A.; Akhtar, J.; Jacobsen, S.E. Antioxidative Response of Quinoa Exposed to Iso-Osmotic, Ionic and Non-Ionic Salt Stress. J. Agron. Crop. Sci. 2015, 201, 452–460. [Google Scholar] [CrossRef]
- Podleśny, J.; Podleśna, A.; Gładyszewska, B.; Bojarszczuk, J. Effect of Pre-Sowing Magnetic Field Treatment on Enzymes and Phytohormones in Pea (Pisum sativum L.) Seeds and Seedlings. Agronomy 2021, 11, 494. [Google Scholar] [CrossRef]
- Poojari, Y.; Clarson, S.J. Thermal stability of Candida antarctica lipase Bimmobilized on macroporous acrylic resin particles in organic media. Biocatal. Agric. Biotechnol. 2013, 2, 7–11. [Google Scholar] [CrossRef]
- Zdarta, J.; Kłapiszewski, L.; Jędrzak, A.; Nowicki, M.; Moszyński, D.; Jesionowski, T. Lipase B from Candida antarctica Immobilized on a Silica-Lignin Matrix as a Stable and Reusable Biocatalytic System. Catalysts 2017, 7, 14. [Google Scholar] [CrossRef] [Green Version]
- Jafarian, F.; Bordbar, A.K.; Zare, A.; Khosropour, A. The performance of immobilized Candida rugosa lipase on various surface modified graphene oxide nanosheets. Int. J. Biol. Macromol. 2018, 111, 1166–1174. [Google Scholar] [CrossRef] [PubMed]
- Siódmiak, T.; Ziegler-Borowska, M.; Marszałł, M.P. Lipase-immobilized magnetic chitosan nanoparticles for kinetic resolution of (R,S)-ibuprofen. J. Mol. Catal. B Enzym. 2013, 94, 7–14. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Zdarta, J.; Kłapiszewski, L.; Wysokowski, M.; Norman, M.; Kołodziejczak-Radzimska, A.; Moszynski, D.; Ehrlich, H.; Maciejewski, H.; Stelling, A.L.; Jesionowski, T. Chitin-Lignin Material as a Novel Matrix for Enzyme Immobilization. Mar. Drugs 2015, 13, 2424–2446. [Google Scholar] [CrossRef] [Green Version]
- Zdarta, J.; Jesionowski, T. Luffa cylindrica Sponges as a Thermally and Chemically Stable Support for Aspergillus niger Lipase. Biotechnol. Prog. 2016, 32, 657–665. [Google Scholar] [CrossRef]
- Bie, J.; Sepodes, B.; Fernandes, P.C.B.; Ribeiro, M.H.L. Enzyme Immobilization and Co-Immobilization: Main Framework, Advances and Some Applications. Processes 2022, 10, 494. [Google Scholar] [CrossRef]
- Boudrant, J.; Woodley, J.M.; Fernandez-Lafuente, R. Parameters necessary to define an immobilized enzyme preparation. Process Biochem. 2020, 90, 66–80. [Google Scholar] [CrossRef]
- Sun, J.N.; Jiang, Y.J.; Zhou, L.Y.; Gao, J. Immobilization of Candida antarctica lipase B by adsorption in organic medium. N. Biotechnol. 2010, 27, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, E.A.L.; Lima, A.S.; Soares, C.M.F.; Santana, L. Lipase from Aspergillus niger obtained from mangaba residue fermentation: Biochemical characterization of free and immobilized enzymes on a sol-gel matrix. Acta Sci.-Technol. 2017, 39, 1–8. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | U (Immobilized CRL-OF) | Activity Recovery (%) |
|---|---|---|
| 4 | 34.37 ± 2.81 | 22.86 ± 0.02 |
| 7 | 37.65 ± 2.84 | 25.04 ± 0.02 |
| 9 | 10.78 ± 1.08 | 7.17 ± 0.01 |
| Storage Conditions | Test Tube | CALB | CRL-OF | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Temperature 65 °C | Humidity 75.0% | Vis 400–800 nm | Open/Closed/Dark Glass | CALB Residual Activity (%) | Activity (U) | Activity (U/g Support) | CRL-OF Residual Activity (%) | Activity (U) | Activity (U/g Support) |
| + | + | + | open | 171.3 ± 2.6 | 1.80 ± 0.13 | 36.04 ± 2.55 | 3.6 ± 0.6 | 1.30 ± 0.25 | 26.04 ± 5.09 |
| + | + | closed | 218.8 ± 4.6 | 2.30 ± 0.17 | 46.04 ± 3.47 | 5.4 ± 0.6 | 1.97 ± 0.25 | 39.38 ± 5.09 | |
| + | + | dark glass open | 218.8 ± 7.3 | 2.30 ± 0.25 | 46.04 ± 5.09 | 4.0 ± 0.8 | 1.47 ± 0.25 | 29.38 ± 5.09 | |
| + | dark glass closed | 203.0 ± 4.0 | 2.14 ± 0.21 | 42.71 ± 4.19 | 2.2 ± 0.2 | 0.80 ± 0.10 | 16.04 ± 1.92 | ||
| Refrigerator [4 °C] | closed | 203.0 ± 4.6 | 2.14 ± 0.17 | 42.71 ± 3.47 | 96.8 ± 5.2 | 35.47 ± 2.59 | 709.38 ± 51.75 | ||
| Dry Form of Lipases | ||||
|---|---|---|---|---|
| Type of Glass | Type of Tube | Incubation Time (days) | Temperature (°C)/Humidity (%) | Vis (400–800 nm) |
| Dark | Open 1 | 7 | 65/75 | - |
| Closed 1 | 65/- | |||
| Transparent | Open 1 | 65/75 | + | |
| Closed 1 | 65/- | |||
| Closed 2 | 4/- | - | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siódmiak, T.; Siódmiak, J.; Mastalerz, R.; Kocot, N.; Dulęba, J.; Haraldsson, G.G.; Wątróbska-Świetlikowska, D.; Marszałł, M.P. Climatic Chamber Stability Tests of Lipase-Catalytic Octyl-Sepharose Systems. Catalysts 2023, 13, 501. https://doi.org/10.3390/catal13030501
Siódmiak T, Siódmiak J, Mastalerz R, Kocot N, Dulęba J, Haraldsson GG, Wątróbska-Świetlikowska D, Marszałł MP. Climatic Chamber Stability Tests of Lipase-Catalytic Octyl-Sepharose Systems. Catalysts. 2023; 13(3):501. https://doi.org/10.3390/catal13030501
Chicago/Turabian StyleSiódmiak, Tomasz, Joanna Siódmiak, Rafał Mastalerz, Natalia Kocot, Jacek Dulęba, Gudmundur G. Haraldsson, Dorota Wątróbska-Świetlikowska, and Michał Piotr Marszałł. 2023. "Climatic Chamber Stability Tests of Lipase-Catalytic Octyl-Sepharose Systems" Catalysts 13, no. 3: 501. https://doi.org/10.3390/catal13030501