
Comparison of Four Immobilization Methods for Different Transaminases
 , , , and
, , , and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Immobilization of the Transaminases
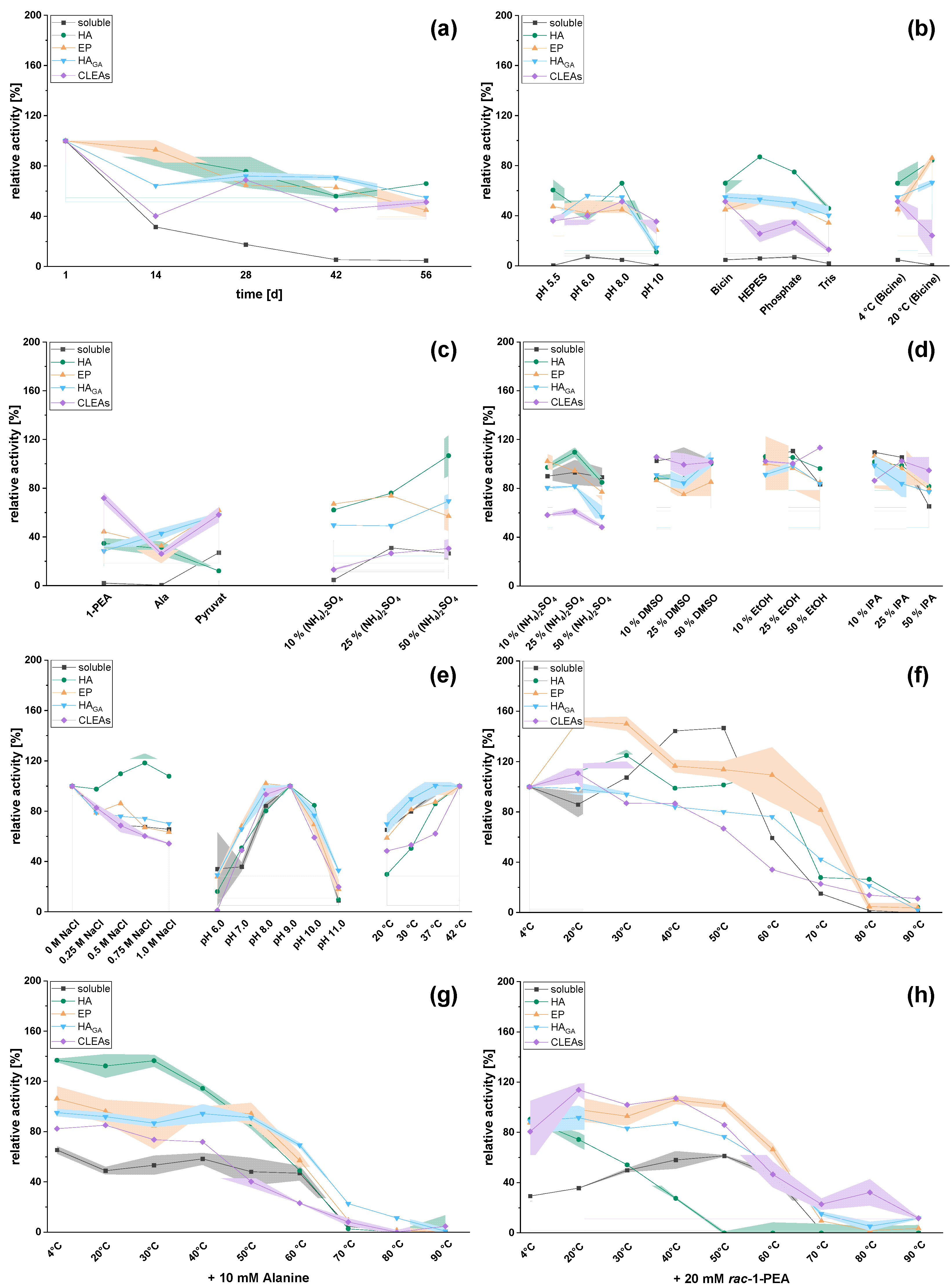
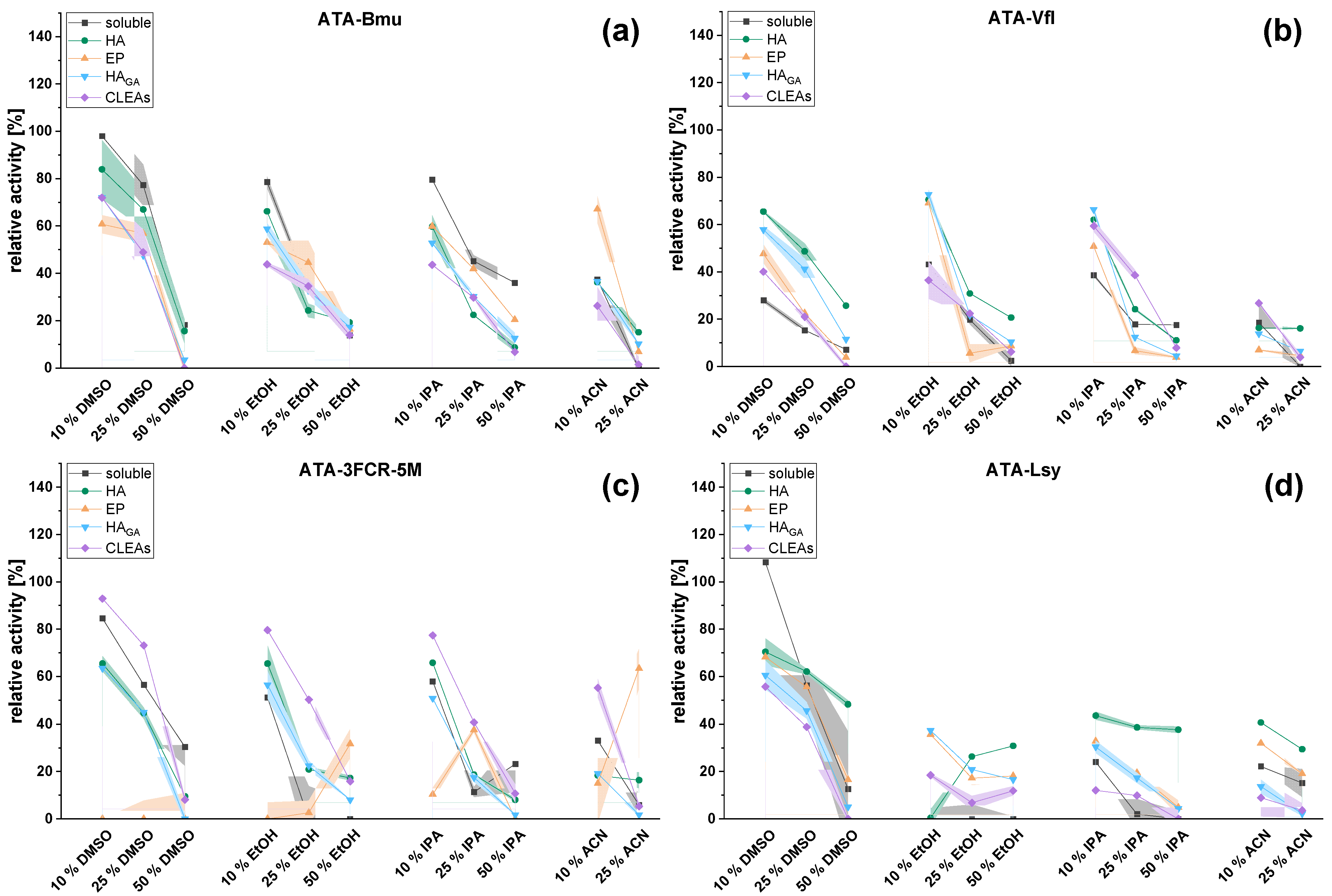
2.2. Effect of Immobilization: Reactivities and Stabilities of Soluble and Immobilized Transaminases
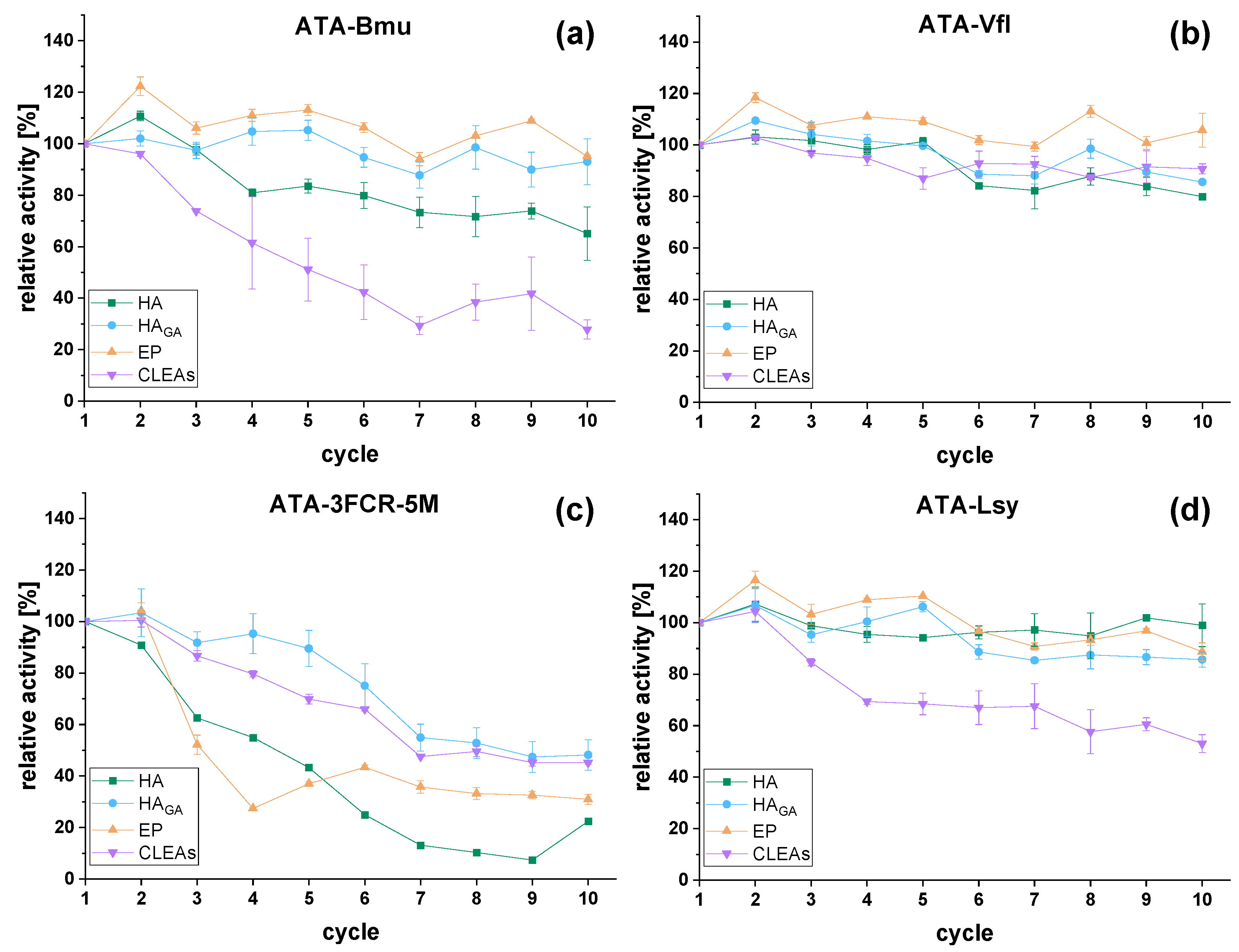
2.3. Reusability of Immobilized Transaminases
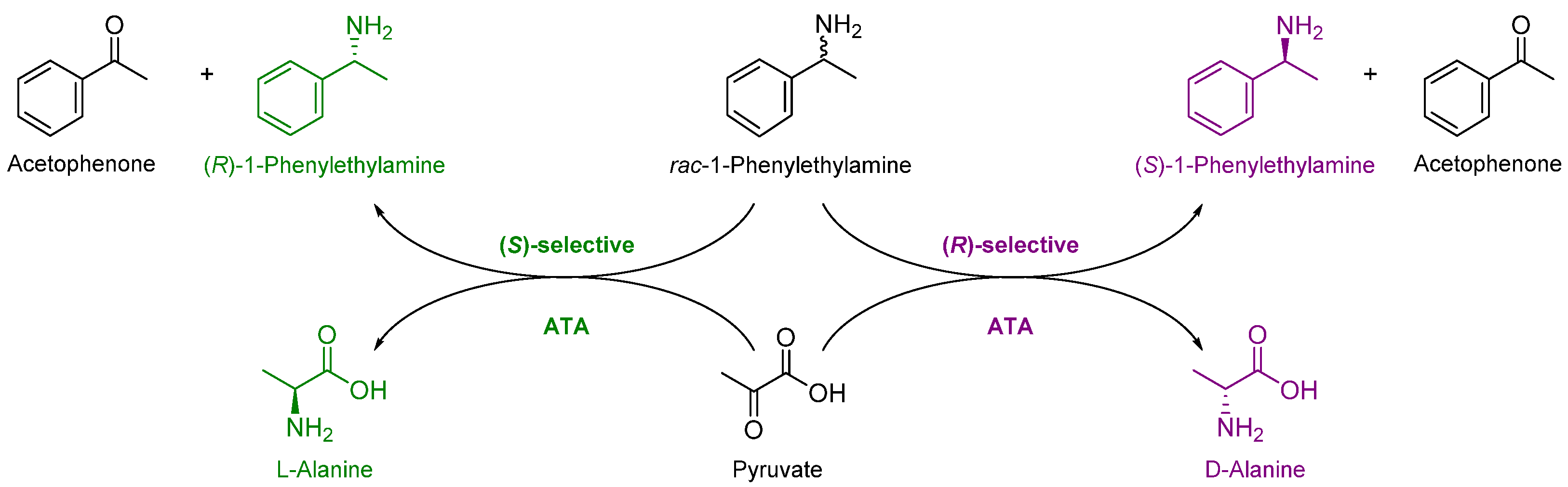
2.4. Upscaled Kinetic Resolution of rac-1-PEA Catalyzed by Immobilized Transaminases
3. Materials and Methods
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Savile, C.K.; Janey, J.M.; Mundorff, E.C.; Moore, J.C.; Tam, S.; Jarvis, W.R.; Colbeck, J.C.; Krebber, A.; Fleitz, F.J.; Brands, J.; et al. Biocatalytic asymmetric synthesis of chiral amines from ketones applied to sitagliptin manufacture. Science 2010, 329, 305–309. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, M.; Koszelewski, D.; Tauber, K.; Kroutil, W.; Faber, K. Chemoenzymatic asymmetric total synthesis of (S)-Rivastigmine using omega-transaminases. Chem. Commun. 2010, 46, 5500–5502. [Google Scholar] [CrossRef] [PubMed]
- Busto, E.; Simon, R.C.; Grischek, B.; Gotor-Fernández, V.; Kroutil, W. Cutting short the asymmetric synthesis of the ramatroban precursor by employing ω-transaminases. Adv. Synth. Catal. 2014, 356, 1937–1942. [Google Scholar] [CrossRef]
- Ghislieri, D.; Turner, N.J. Biocatalytic approaches to the synthesis of enantiomerically pure chiral amines. Top. Catal. 2014, 57, 284–300. [Google Scholar] [CrossRef]
- Tufvesson, P.; Lima-Ramos, J.; Jensen, J.S.; Al-Haque, N.; Neto, W.; Woodley, J.M. Process considerations for the asymmetric synthesis of chiral amines using transaminases. Biotechnol. Bioeng. 2011, 108, 1479–1493. [Google Scholar] [CrossRef]
- Singh, R.K.; Tiwari, M.K.; Singh, R.; Lee, J.-K. From protein engineering to immobilization: Promising strategies for the upgrade of industrial enzymes. Int. J. Mol. Sci. 2013, 14, 1232–1277. [Google Scholar] [CrossRef]
- Börner, T.; Rämisch, S.; Reddem, E.R.; Bartsch, S.; Vogel, A.; Thunnissen, A.-M.W.H.; Adlercreutz, P.; Grey, C. Explaining operational instability of amine transaminases: Substrate-induced inactivation mechanism and influence of quaternary structure on enzyme–cofactor intermediate stability. ACS Catal. 2017, 7, 1259–1269. [Google Scholar] [CrossRef]
- Chen, S.; Land, H.; Berglund, P.; Humble, M.S. Stabilization of an amine transaminase for biocatalysis. J. Mol. Catal. B Enzym. 2016, 124, 20–28. [Google Scholar] [CrossRef]
- Guo, F.; Berglund, P. Transaminase biocatalysis: Optimization and application. Green Chem. 2017, 19, 333–360. [Google Scholar] [CrossRef] [Green Version]
- Kelly, S.A.; Pohle, S.; Wharry, S.; Mix, S.; Allen, C.C.R.; Moody, T.S.; Gilmore, B.F. Application of ω-transaminases in the pharmaceutical industry. Chem. Rev. 2018, 118, 349–367. [Google Scholar] [CrossRef]
- Truppo, M.D.; Strotman, H.; Hughes, G. Development of an immobilized transaminase capable of operating in organic solvent. ChemCatChem 2012, 4, 1071–1074. [Google Scholar] [CrossRef]
- Rodrigues, R.C.; Ortiz, C.; Berenguer-Murcia, Á.; Torres, R.; Fernández-Lafuente, R. Modifying enzyme activity and selectivity by immobilization. Chem. Soc. Rev. 2013, 42, 6290–6307. [Google Scholar] [CrossRef]
- Kaličanin, N.; Kovačević, G.; Spasojević, M.; Prodanović, O.; Jovanović-Šanta, S.; Škorić, D.; Opsenica, D.; Prodanović, R. Immobilization of ArRMut11 omega-transaminase for increased operational stability and reusability in the synthesis of 3α-amino-5α-androstan-17β-ol. Process. Biochem. 2022, 121, 674–680. [Google Scholar] [CrossRef]
- Liese, A.; Hilterhaus, L. Evaluation of immobilized enzymes for industrial applications. Chem. Soc. Rev. 2013, 42, 6236–6249. [Google Scholar] [CrossRef]
- Bornscheuer, U.T. Immobilizing enzymes: How to create more suitable biocatalysts. Angew. Chem. Int. Ed. 2003, 42, 3336–3337. [Google Scholar] [CrossRef]
- Neto, W.; Schürmann, M.; Panella, L.; Vogel, A.; Woodley, J.M. Immobilisation of ω-transaminase for industrial application: Screening and characterisation of commercial ready to use enzyme carriers. J. Mol. Catal. B Enzym. 2015, 117, 54–61. [Google Scholar] [CrossRef]
- Velasco-Lozano, S.; Jackson, E.; Ripoll, M.; López-Gallego, F.; Betancor, L. Stabilization of ω-transaminase from Pseudomonas fluorescens by immobilization techniques. Int. J. Biol. Macromol. 2020, 164, 4318–4328. [Google Scholar] [CrossRef]
- Deepankumar, K.; Nadarajan, S.P.; Mathew, S.; Lee, S.-G.; Yoo, T.H.; Hong, E.Y.; Kim, B.-G.; Yun, H. Engineering transaminase for stability enhancement and site-specific immobilization through multiple noncanonical amino acids incorporation. ChemCatChem 2015, 7, 417–421. [Google Scholar] [CrossRef]
- Basso, A.; Serban, S. Industrial applications of immobilized enzymes—A review. Mol. Catal. 2019, 479, 110607. [Google Scholar] [CrossRef]
- Fernandez-Lafuente, R. Stabilization of multimeric enzymes: Strategies to prevent subunit dissociation. Enzym. Microb. Technol. 2009, 45, 405–418. [Google Scholar] [CrossRef]
- Carballares, D.; Morellon-Sterling, R.; Xu, X.; Hollmann, F.; Fernandez-Lafuente, R. Immobilization of the peroxygenase from Agrocybe aegerita. The effect of the immobilization pH on the features of an ionically exchanged dimeric peroxygenase. Catalysts 2021, 11, 560. [Google Scholar] [CrossRef]
- Deshmukh, S.S.; Dutta Choudhury, M.; Shankar, V. Preparation and properties of glucose isomerase immobilized on Indion 48-R. Appl. Biochem. Biotechnol. 1993, 42, 95–104. [Google Scholar] [CrossRef]
- Kappauf, K.; Majstorovic, N.; Agarwal, S.; Rother, D.; Claaßen, C. Modulation of transaminase activity by encapsulation in temperature-sensitive poly(N-acryloyl glycinamide) hydrogels. ChemBioChem 2021, 22, 3452–3461. [Google Scholar] [CrossRef]
- Talekar, S.; Joshi, A.; Joshi, G.; Kamat, P.; Haripurkar, R.; Kambale, S. Parameters in preparation and characterization of cross linked enzyme aggregates (CLEAs). RSC Adv. 2013, 3, 12485–12511. [Google Scholar] [CrossRef]
- Roy, I.; Mukherjee, J.; Gupta, M.N. Cross-linked enzyme aggregates for applications in aqueous and nonaqueous media. Methods Mol. Biol. 2017, 1504, 109–123. [Google Scholar] [CrossRef]
- Rojas, M.; Amaral-Fonseca, M.; Zanin, G.; Fernandez-Lafuente, R.; Giordano, R.; Tardioli, P. Preparation of crosslinked enzyme aggregates of a thermostable cyclodextrin glucosyltransferase from Thermoanaerobacter sp. critical effect of the crosslinking agent. Catalysts 2019, 9, 120. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, H.; Kiyota, Y.; Miyazaki, M. Techniques for preparation of cross-linked enzyme aggregates and their applications in bioconversions. Catalysts 2018, 8, 174. [Google Scholar] [CrossRef] [Green Version]
- Kahar, U.M.; Sani, M.H.; Chan, K.-G.; Goh, K.M. Immobilization of α-amylase from Anoxybacillus sp. SK3-4 on ReliZyme and immobead supports. Molecules 2016, 21, 1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Gallego, F.; Guisan, J.M.; Betancor, L. Immobilization of enzymes on supports activated with glutaraldehyde: A very simple immobilization protocol. Methods Mol. Biol. 2020, 2100, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Betancor, L.; López-Gallego, F.; Hidalgo, A.; Alonso-Morales, N.; Dellamora-Ortiz, G.M.; Fernández-Lafuente, R.; Guisán, J.M. Different mechanisms of protein immobilization on glutaraldehyde activated supports: Effect of support activation and immobilization conditions. Enzym. Microb. Technol. 2006, 39, 877–882. [Google Scholar] [CrossRef]
- Bezbradica, D.I.; Mateo, C.; Guisan, J.M. Novel support for enzyme immobilization prepared by chemical activation with cysteine and glutaraldehyde. J. Mol. Catal. B Enzym. 2014, 102, 218–224. [Google Scholar] [CrossRef]
- De Souza, S.P.; Junior, I.I.; Silva, G.M.A.; Miranda, L.S.M.; Santiago, M.F.; Leung-Yuk Lam, F.; Dawood, A.; Bornscheuer, U.T.; de Souza, R.O.M.A. Cellulose as an efficient matrix for lipase and transaminase immobilization. RSC Adv. 2016, 6, 6665–6671. [Google Scholar] [CrossRef]
- Mallin, H.; Höhne, M.; Bornscheuer, U.T. Immobilization of (R)- and (S)-amine transaminases on chitosan support and their application for amine synthesis using isopropylamine as donor. J. Biotechnol. 2014, 191, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Jia, D.-X.; Xu, H.-P.; Sun, C.-Y.; Peng, C.; Li, J.-L.; Jin, L.-Q.; Cheng, F.; Liu, Z.-Q.; Xue, Y.-P.; Zheng, Y.-G. Covalent immobilization of recombinant Citrobacter koseri transaminase onto epoxy resins for consecutive asymmetric synthesis of l-phosphinothricin. Bioprocess. Biosyst. Eng. 2020, 43, 1599–1607. [Google Scholar] [CrossRef]
- Abaházi, E.; Sátorhelyi, P.; Erdélyi, B.; Vértessy, B.G.; Land, H.; Paizs, C.; Berglund, P.; Poppe, L. Covalently immobilized Trp60Cys mutant of ω-transaminase from Chromobacterium violaceum for kinetic resolution of racemic amines in batch and continuous-flow modes. Biochem. Eng. J. 2018, 132, 270–278. [Google Scholar] [CrossRef]
- Bilgin, R.; Yalcin, M.S.; Yildirim, D. Optimization of covalent immobilization of Trichoderma reesei cellulase onto modified ReliZyme HA403 and Sepabeads EC-EP supports for cellulose hydrolysis, in buffer and ionic liquids/buffer media. Artif. Cells Nanomed. Biotechnol. 2016, 44, 1276–1284. [Google Scholar] [CrossRef] [PubMed]
- Bayraktar, H.; Serilmez, M.; Karkaş, T.; Celem, E.B.; Onal, S. Immobilization and stabilization of α-galactosidase on Sepabeads EC-EA and EC-HA. Int. J. Biol. Macromol. 2011, 49, 855–860. [Google Scholar] [CrossRef] [PubMed]
- Yepes, C.; Estévez, J.; Arroyo, M.; Ladero, M. Immobilization of an industrial β-glucosidase from Aspergillus fumigatus and its use for cellobiose hydrolysis. Processes 2022, 10, 1225. [Google Scholar] [CrossRef]
- Nagy, F.; Gyujto, I.; Tasnádi, G.; Barna, B.; Balogh-Weiser, D.; Faber, K.; Poppe, L.; Hall, M. Design and application of a bi-functional redox biocatalyst through covalent co-immobilization of ene-reductase and glucose dehydrogenase. J. Biotechnol. 2020, 323, 246–253. [Google Scholar] [CrossRef]
- Wang, X.; Xie, Y.; Wang, Z.; Zhang, K.; Wang, H.; Wei, D. Efficient synthesis of (S)-1-boc-3-aminopiperidine in a continuous flow system using ω-transaminase-immobilized amino-ethylenediamine-modified epoxide supports. Org. Process. Res. Dev. 2022, 26, 1351–1359. [Google Scholar] [CrossRef]
- Li, M.; Cheng, F.; Li, H.; Jin, W.; Chen, C.; He, W.; Cheng, G.; Wang, Q. Site-specific and covalent immobilization of His-tagged proteins via surface vinyl sulfone-imidazole coupling. Langmuir 2019, 35, 16466–16475. [Google Scholar] [CrossRef] [PubMed]
- Ley, C.; Holtmann, D.; Mangold, K.-M.; Schrader, J. Immobilization of histidine-tagged proteins on electrodes. Colloids Surf. B Biointerfaces 2011, 88, 539–551. [Google Scholar] [CrossRef]
- Zhong, M.; Fang, J.; Wei, Y. Site specific and reversible protein immobilization facilitated by a DNA binding fusion tag. Bioconjugate Chem. 2010, 21, 1177–1182. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-H.; Tang, M.-Z.; Yu, X.-T.; Xu, C.-M.; Yang, H.-M.; Tang, J.-B. Site-specific, covalent immobilization of an engineered enterokinase onto magnetic nanoparticles through transglutaminase-catalyzed bioconjugation. Colloids Surf. B Biointerfaces 2019, 177, 506–511. [Google Scholar] [CrossRef] [PubMed]
- Camarero, J.A. Recent developments in the site-specific immobilization of proteins onto solid supports. Biopolymers 2008, 90, 450–458. [Google Scholar] [CrossRef] [PubMed]
- Janson, N.; Heinks, T.; Beuel, T.; Alam, S.; Höhne, M.; Bornscheuer, U.T.; von Mollard, G.F.; Sewald, N. Efficient site-selective immobilization of aldehyde-tagged peptides and proteins by knoevenagel ligation. ChemCatChem 2022, 14, e202101485. [Google Scholar] [CrossRef]
- Raliski, B.K.; Howard, C.A.; Young, D.D. Site-specific protein immobilization using unnatural amino acids. Bioconjugate Chem. 2014, 25, 1916–1920. [Google Scholar] [CrossRef]
- Peng, J.; Alam, S.; Radhakrishnan, K.; Mariappan, M.; Rudolph, M.G.; May, C.; Dierks, T.; von Figura, K.; Schmidt, B. Eukaryotic formylglycine-generating enzyme catalyses a monooxygenase type of reaction. FEBS J. 2015, 282, 3262–3274. [Google Scholar] [CrossRef] [PubMed]
- Dierks, T.; Lecca, M.R.; Schlotterhose, P.; Schmidt, B.; von Figura, K. Sequence determinants directing conversion of cysteine to formylglycine in eukaryotic sulfatases. EMBO J. 1999, 18, 2084–2091. [Google Scholar] [CrossRef] [Green Version]
- Peng, Q.; Zang, B.; Zhao, W.; Li, D.; Ren, J.; Ji, F.; Jia, L. Efficient continuous-flow aldehyde tag conversion using immobilized formylglycine generating enzyme. Catal. Sci. Technol. 2020, 10, 484–492. [Google Scholar] [CrossRef]
- Ju, W.; Zhen, X.; Zheng, L. Soluble expression and biomimetic immobilization of a ω-transaminase from Bacillus subtilis: Development of an efficient and recyclable biocatalyst. Biochem. Eng. J. 2020, 160, 107635. [Google Scholar] [CrossRef]
- Böhmer, W.; Knaus, T.; Volkov, A.; Slot, T.K.; Shiju, N.R.; Cassimjee, K.E.; Mutti, F.G. Highly efficient production of chiral amines in batch and continuous flow by immobilized ω-transaminases on controlled porosity glass metal-ion affinity carrier. J. Biotechnol. 2019, 291, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Yi, S.-S.; Lee, C.; Kim, J.; Kyung, D.; Kim, B.-G.; Lee, Y.-S. Covalent immobilization of ω-transaminase from Vibrio fluvialis JS17 on chitosan beads. Process. Biochem. 2007, 42, 895–898. [Google Scholar] [CrossRef]
- Molnár, Z.; Farkas, E.; Lakó, Á.; Erdélyi, B.; Kroutil, W.; Vértessy, B.G.; Paizs, C.; Poppe, L. Immobilized whole-cell transaminase biocatalysts for continuous-flow kinetic resolution of amines. Catalysts 2019, 9, 438. [Google Scholar] [CrossRef] [Green Version]
- Cárdenas-Fernández, M.; Neto, W.; López, C.; Álvaro, G.; Tufvesson, P.; Woodley, J.M. Immobilization of Escherichia coli containing ω-transaminase activity in LentiKats®. Biotechnol. Prog. 2012, 28, 693–698. [Google Scholar] [CrossRef]
- Mallin, H.; Menyes, U.; Vorhaben, T.; Höhne, M.; Bornscheuer, U.T. Immobilization of two (R)-amine transaminases on an optimized chitosan support for the enzymatic synthesis of optically pure amines. ChemCatChem 2013, 5, 588–593. [Google Scholar] [CrossRef]
- Martin, A.R.; Shonnard, D.; Pannuri, S.; Kamat, S. Characterization of free and immobilized (S)-aminotransferase for acetophenone production. Appl. Microbiol. Biotechnol. 2007, 76, 843–851. [Google Scholar] [CrossRef]
- Ni, K.; Zhou, X.; Zhao, L.; Wang, H.; Ren, Y.; Wei, D. Magnetic catechol-chitosan with bioinspired adhesive surface: Preparation and immobilization of ω-transaminase. PLoS ONE 2012, 7, e41101. [Google Scholar] [CrossRef] [Green Version]
- Semproli, R.; Vaccaro, G.; Ferrandi, E.E.; Vanoni, M.; Bavaro, T.; Marrubini, G.; Annunziata, F.; Conti, P.; Speranza, G.; Monti, D.; et al. Use of immobilized amine transaminase from Vibrio fluvialis under flow conditions for the synthesis of (S)-1-(5-fluoropyrimidin-2-yl)-ethanamine. ChemCatChem 2020, 12, 1359–1367. [Google Scholar] [CrossRef]
- Genz, M.; Vickers, C.; van den Bergh, T.; Joosten, H.-J.; Dörr, M.; Höhne, M.; Bornscheuer, U.T. Alteration of the donor/acceptor spectrum of the (S)-amine transaminase from Vibrio fluvialis. Int. J. Mol. Sci. 2015, 16, 26953–26963. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.-C.; Yun, H.; Park, H.H. Structural dynamics of the transaminase active site revealed by the crystal structure of a co-factor free omega-transaminase from Vibrio fluvialis JS17. Sci. Rep. 2018, 8, 11454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genz, M.; Melse, O.; Schmidt, S.; Vickers, C.; Dörr, M.; van den Bergh, T.; Joosten, H.-J.; Bornscheuer, U.T. Engineering the amine transaminase from Vibrio fluvialis towards branched-chain substrates. ChemCatChem 2016, 8, 3199–3202. [Google Scholar] [CrossRef]
- Chen, S.; Campillo-Brocal, J.C.; Berglund, P.; Humble, M.S. Characterization of the stability of Vibrio fluvialis JS17 amine transaminase. J. Biotechnol. 2018, 282, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.-S.; Yun, H.; Jang, J.-W.; Park, I.; Kim, B.-G. Purification, characterization, and molecular cloning of a novel amine:pyruvate transaminase from Vibrio fluvialis JS17. Appl. Microbiol. Biotechnol. 2003, 61, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Kollipara, M.; Matzel, P.; Sowa, M.; Brott, S.; Bornscheuer, U.; Höhne, M. Characterization of proteins from the 3N5M family reveals an operationally stable amine transaminase. Appl. Microbiol. Biotechnol. 2022, 106, 5563–5574. [Google Scholar] [CrossRef]
- Steffen-Munsberg, F.; Vickers, C.; Thontowi, A.; Schätzle, S.; Meinhardt, T.; Svedendahl Humble, M.; Land, H.; Berglund, P.; Bornscheuer, U.T.; Höhne, M. Revealing the structural basis of promiscuous amine transaminase activity. ChemCatChem 2013, 5, 154–157. [Google Scholar] [CrossRef]
- Steffen-Munsberg, F.; Vickers, C.; Thontowi, A.; Schätzle, S.; Tumlirsch, T.; Svedendahl Humble, M.; Land, H.; Berglund, P.; Bornscheuer, U.T.; Höhne, M. Connecting unexplored protein crystal structures to enzymatic function. ChemCatChem 2013, 5, 150–153. [Google Scholar] [CrossRef]
- Pavlidis, I.V.; Weiß, M.S.; Genz, M.; Spurr, P.; Hanlon, S.P.; Wirz, B.; Iding, H.; Bornscheuer, U.T. Identification of (S)-selective transaminases for the asymmetric synthesis of bulky chiral amines. Nat. Chem. 2016, 8, 1076–1082. [Google Scholar] [CrossRef]
- Höhne, M.; Schätzle, S.; Jochens, H.; Robins, K.; Bornscheuer, U.T. Rational assignment of key motifs for function guides in silico enzyme identification. Nat. Chem. Biol. 2010, 6, 807–813. [Google Scholar] [CrossRef]
- Konia, E.; Chatzicharalampous, K.; Drakonaki, A.; Muenke, C.; Ermler, U.; Tsiotis, G.; Pavlidis, I.V. Rational engineering of Luminiphilus syltensis (R)-selective amine transaminase for the acceptance of bulky substrates. Chem. Commum. 2021, 57, 12948–12951. [Google Scholar] [CrossRef]
- Schätzle, S.; Höhne, M.; Redestad, E.; Robins, K.; Bornscheuer, U.T. Rapid and sensitive kinetic assay for characterization of omega-transaminases. Anal. Chem. 2009, 81, 8244–8248. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.S.; Honig, B. On the pH dependence of protein stability. J. Mol. Biol. 1993, 231, 459–474. [Google Scholar] [CrossRef] [PubMed]
- Mazzini, A.; Polverini, E.; Parisi, M.; Sorbi, R.T.; Favilla, R. Dissociation and unfolding of bovine odorant binding protein at acidic pH. J. Struct. Biol. 2007, 159, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Guranda, D.T.; Volovik, T.S.; Svedas, V.K. pH stability of penicillin acylase from Escherichia coli. Biochemistry Mosc. 2004, 69, 1386–1390. [Google Scholar] [CrossRef] [PubMed]
- López-Gallego, F.; Fernandez-Lorente, G.; Rocha-Martin, J.; Bolivar, J.M.; Mateo, C.; Guisan, J.M. Stabilization of enzymes by multipoint covalent immobilization on supports activated with glyoxyl groups. Methods Mol. Biol. 2013, 1051, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Guisan, J.M.; Fernandez-Lorente, G.; Rocha-Martin, J.; Moreno-Gamero, D. Enzyme immobilization strategies for the design of robust and efficient biocatalysts. Curr. Opin. Green Sustain. Chem. 2022, 35, 100593. [Google Scholar] [CrossRef]
- Wang, B.; Zhou, J.; Zhang, X.-Y.; Yang, Y.-S.; Liu, C.-H.; Zhu, H.-L.; Jiao, Q.-C. Covalently immobilize crude d-amino acid transaminase onto UiO-66-NH2 surface for d-Ala biosynthesis. Int. J. Biol. Macromol. 2021, 175, 451–458. [Google Scholar] [CrossRef]
- Zhang, X.-J.; Fan, H.-H.; Liu, N.; Wang, X.-X.; Cheng, F.; Liu, Z.-Q.; Zheng, Y.-G. A novel self-sufficient biocatalyst based on transaminase and pyridoxal 5’-phosphate covalent co-immobilization and its application in continuous biosynthesis of sitagliptin. Enzym. Microb. Technol. 2019, 130, 109362. [Google Scholar] [CrossRef]
- Shin, J.-S.; Kim, B.-G.; Shin, D.-H. Kinetic resolution of chiral amines using packed-bed reactor. Enzyme Microb. Technol. 2001, 29, 232–239. [Google Scholar] [CrossRef]
- Koszelewski, D.; Müller, N.; Schrittwieser, J.H.; Faber, K.; Kroutil, W. Immobilization of ω-transaminases by encapsulation in a sol–gel/celite matrix. J. Mol. Catal. B Enzym. 2010, 63, 39–44. [Google Scholar] [CrossRef]
- Wang, C.; Tang, K.; Dai, Y.; Jia, H.; Li, Y.; Gao, Z.; Wu, B. Identification, characterization, and site-specific mutagenesis of a thermostable ω-transaminase from Chloroflexi bacterium. ACS Omega 2021, 6, 17058–17070. [Google Scholar] [CrossRef] [PubMed]
- Ferrandi, E.E.; Previdi, A.; Bassanini, I.; Riva, S.; Peng, X.; Monti, D. Novel thermostable amine transferases from hot spring metagenomes. Appl. Microbiol. Biotechnol. 2017, 101, 4963–4979. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.; Yi, Y.; Gao, Z.; Jia, H.; Li, Y.; Cao, F.; Zhou, H.; Jiang, M.; Wei, P. Identification, heterologous expression and characterization of a transaminase from Rhizobium sp. Catal. Lett. 2020, 150, 2415–2426. [Google Scholar] [CrossRef]
- Tentori, F.; Brenna, E.; Crotti, M.; Pedrocchi-Fantoni, G.; Ghezzi, M.C.; Tessaro, D. Continuous-flow biocatalytic process for the synthesis of the best stereoisomers of the commercial fragrances leather cyclohexanol (4-isopropylcyclohexanol) and woody acetate (4-(tert-butyl)cyclohexyl acetate). Catalysts 2020, 10, 102. [Google Scholar] [CrossRef] [Green Version]
- Weltz, J.S.; Kienle, D.F.; Schwartz, D.K.; Kaar, J.L. Reduced enzyme dynamics upon multipoint covalent immobilization leads to dtability-sctivity trade-off. J. Am. Chem. Soc. 2020, 142, 3463–3471. [Google Scholar] [CrossRef]
- Rodrigues, R.C.; Berenguer-Murcia, Á.; Carballares, D.; Morellon-Sterling, R.; Fernandez-Lafuente, R. Stabilization of enzymes via immobilization: Multipoint covalent attachment and other stabilization strategies. Biotechnol. Adv. 2021, 52, 107821. [Google Scholar] [CrossRef]
- Ruggieri, F.; Campillo-Brocal, J.C.; Chen, S.; Humble, M.S.; Walse, B.; Logan, D.T.; Berglund, P. Insight into the dimer dissociation process of the Chromobacterium violaceum (S)-selective amine transaminase. Sci. Rep. 2019, 9, 16946. [Google Scholar] [CrossRef] [Green Version]
- Börner, T.; Rämisch, S.; Bartsch, S.; Vogel, A.; Adlercreutz, P.; Grey, C. Three in one: Temperature, solvent and catalytic stability by engineering the cofactor-binding element of amine transaminase. ChemBioChem 2017, 18, 1482–1486. [Google Scholar] [CrossRef]
- Heinks, T.; Paulus, J.; Koopmeiners, S.; Beuel, T.; Sewald, N.; Höhne, M.; Bornscheuer, U.T.; Fischer von Mollard, G. Recombinant l-amino acid oxidase with broad substrate spectrum for co-substrate recycling in (S)-selective transaminase-catalyzed kinetic resolutions. ChemBioChem 2022, 23, e202200329. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Merz, L.M.; van Langen, L.M.; Berglund, P. The role of buffer, pyridoxal 5′-phosphate and light on the stability of the Silicibacter pomeroyi transaminase. ChemCatChem 2022, e202201174. [Google Scholar] [CrossRef]
- Seo, J.-H.; Kyung, D.; Joo, K.; Lee, J.; Kim, B.-G. Necessary and sufficient conditions for the asymmetric synthesis of chiral amines using ω-aminotransferases. Biotechnol. Bioeng. 2011, 108, 253–263. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transaminase | Type | Specific Activity of Biocatalyst [U/g bead] 1 | Binding Efficiency [%] 2 | Activity Recovery [%] 3 |
|---|---|---|---|---|
| ATA-Vfl | HA | 53 | 97.6 | 6 |
| HAGA | 62 | 52.7 | 7 | |
| EP | 49 | 64.0 | 5 | |
| ATA-Bmu | HA | 26 | 97.1 | 17 |
| HAGA | 55 | 69.3 | 19 | |
| EP | 26 | 97.7 | 20 | |
| ATA-3FCR-5M | HA | 44 | 95.4 | 15 |
| HAGA | 44 | 98.2 | 50 | |
| EP | 15 | 77.3 | 2 | |
| ATA-Lsy | HA | 48 | 46.9 | 15 |
| HAGA | 34 | 94.6 | 16 | |
| EP | 36 | 62.6 | 12 |
| Transaminase | Type of Immobilization | Enantiomeric Excess [%ee] [a,c] | Obtained Enantiomer [a,c] | Product Yield [%] [b,c] |
|---|---|---|---|---|
| ATA-Bmu | HAGA | 98.8 | (R) | 72 |
| ATA-Vfl | HAGA | 99.8 | (R) | 79 |
| ATA-3FCR-5M | HAGA | 87.8 | (R) | 50 |
| ATA-Lsy | HA | 98.2 | (S) | 57 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heinks, T.; Montua, N.; Teune, M.; Liedtke, J.; Höhne, M.; Bornscheuer, U.T.; Fischer von Mollard, G. Comparison of Four Immobilization Methods for Different Transaminases. Catalysts 2023, 13, 300. https://doi.org/10.3390/catal13020300
Heinks T, Montua N, Teune M, Liedtke J, Höhne M, Bornscheuer UT, Fischer von Mollard G. Comparison of Four Immobilization Methods for Different Transaminases. Catalysts. 2023; 13(2):300. https://doi.org/10.3390/catal13020300
Chicago/Turabian StyleHeinks, Tobias, Nicolai Montua, Michelle Teune, Jan Liedtke, Matthias Höhne, Uwe T. Bornscheuer, and Gabriele Fischer von Mollard. 2023. "Comparison of Four Immobilization Methods for Different Transaminases" Catalysts 13, no. 2: 300. https://doi.org/10.3390/catal13020300