
The High ‘Lipolytic Jump’ of Immobilized Amano A Lipase from Aspergillus niger in Developed ‘ESS Catalytic Triangles’ Containing Natural Origin Substrates
 , , ,
, , ,
Abstract
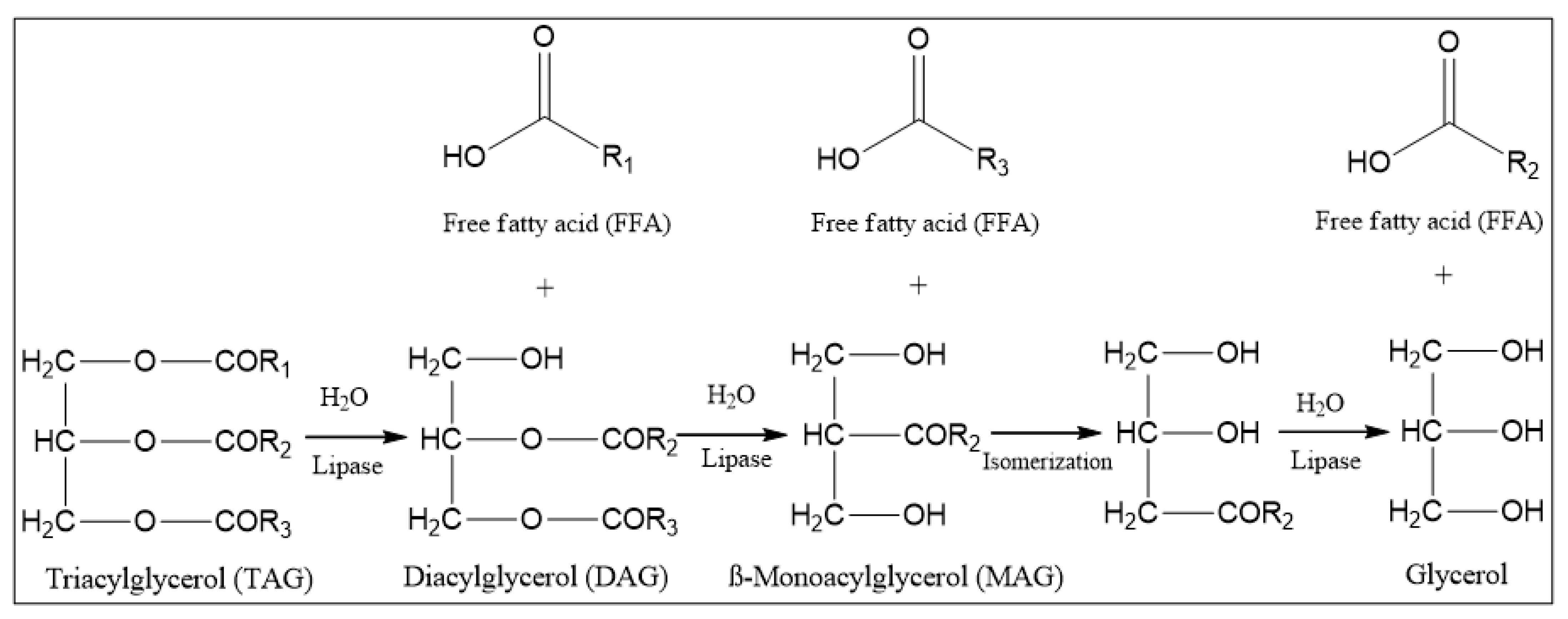
:1. Introduction
2. Results and Discussion
2.1. Effect of Substrates on the Lipolytic Activity of Immobilized AA-ANL
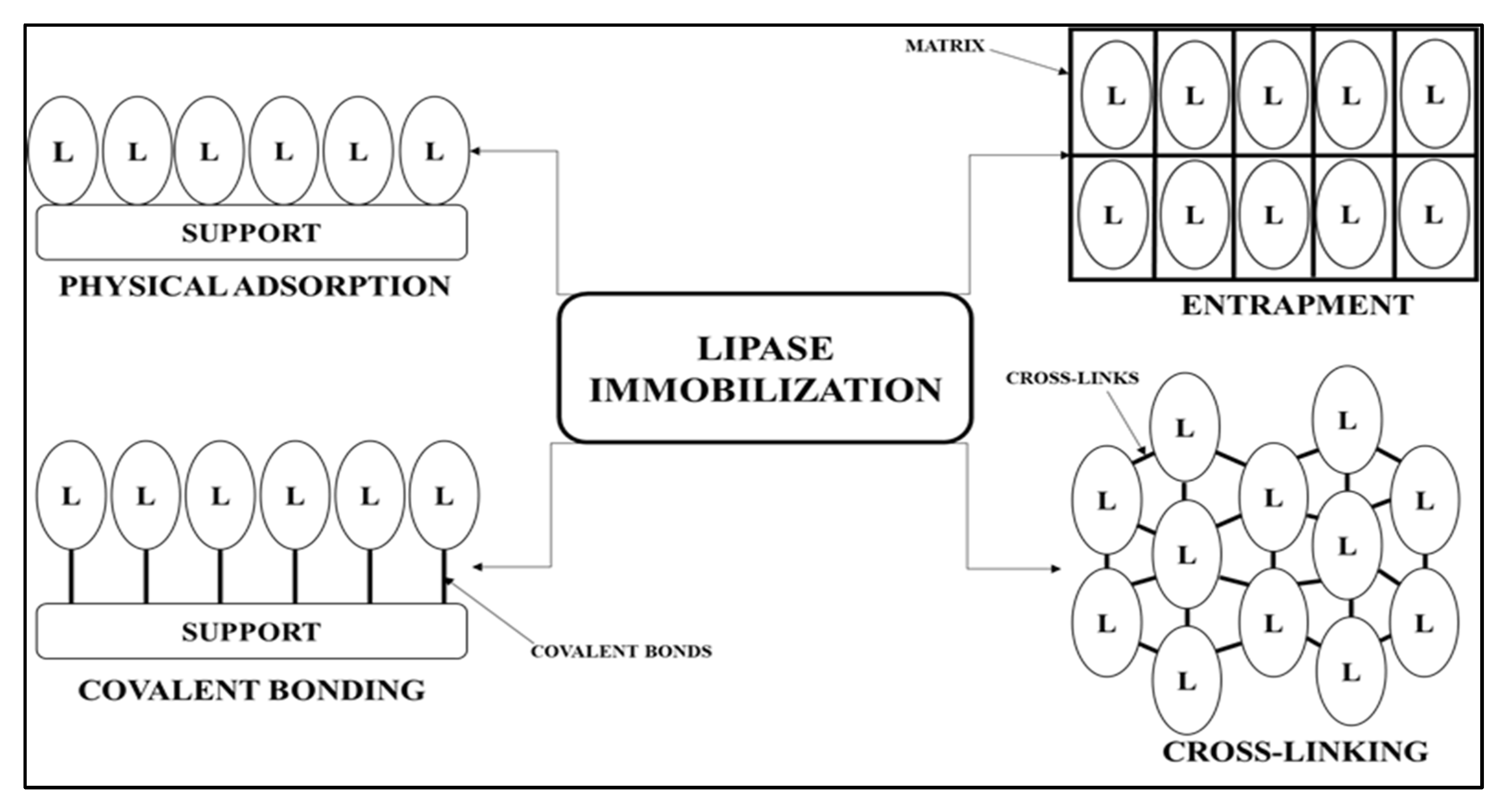
2.2. The Effect of the Support on AA-ANL Lipolytic Activity
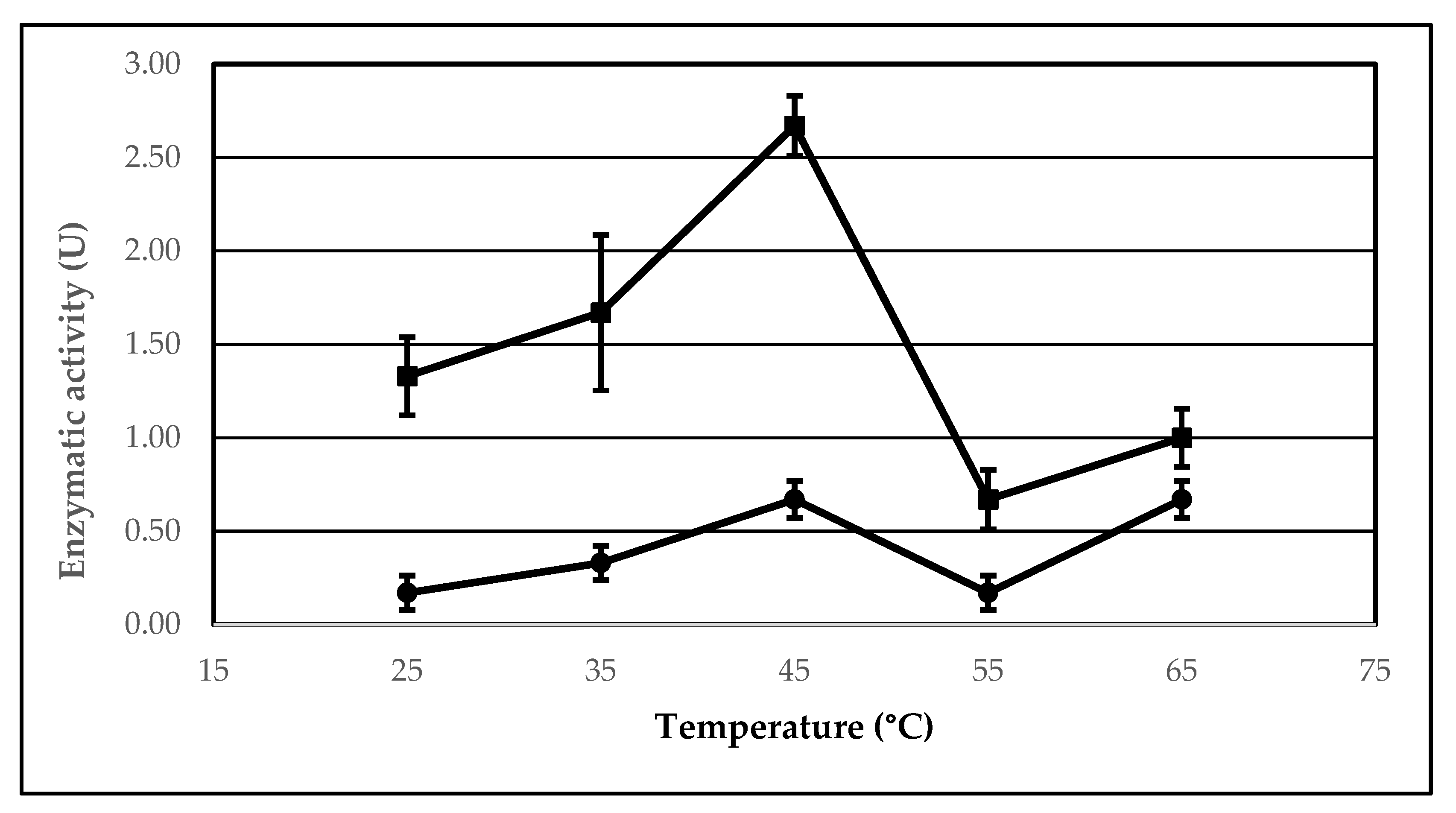
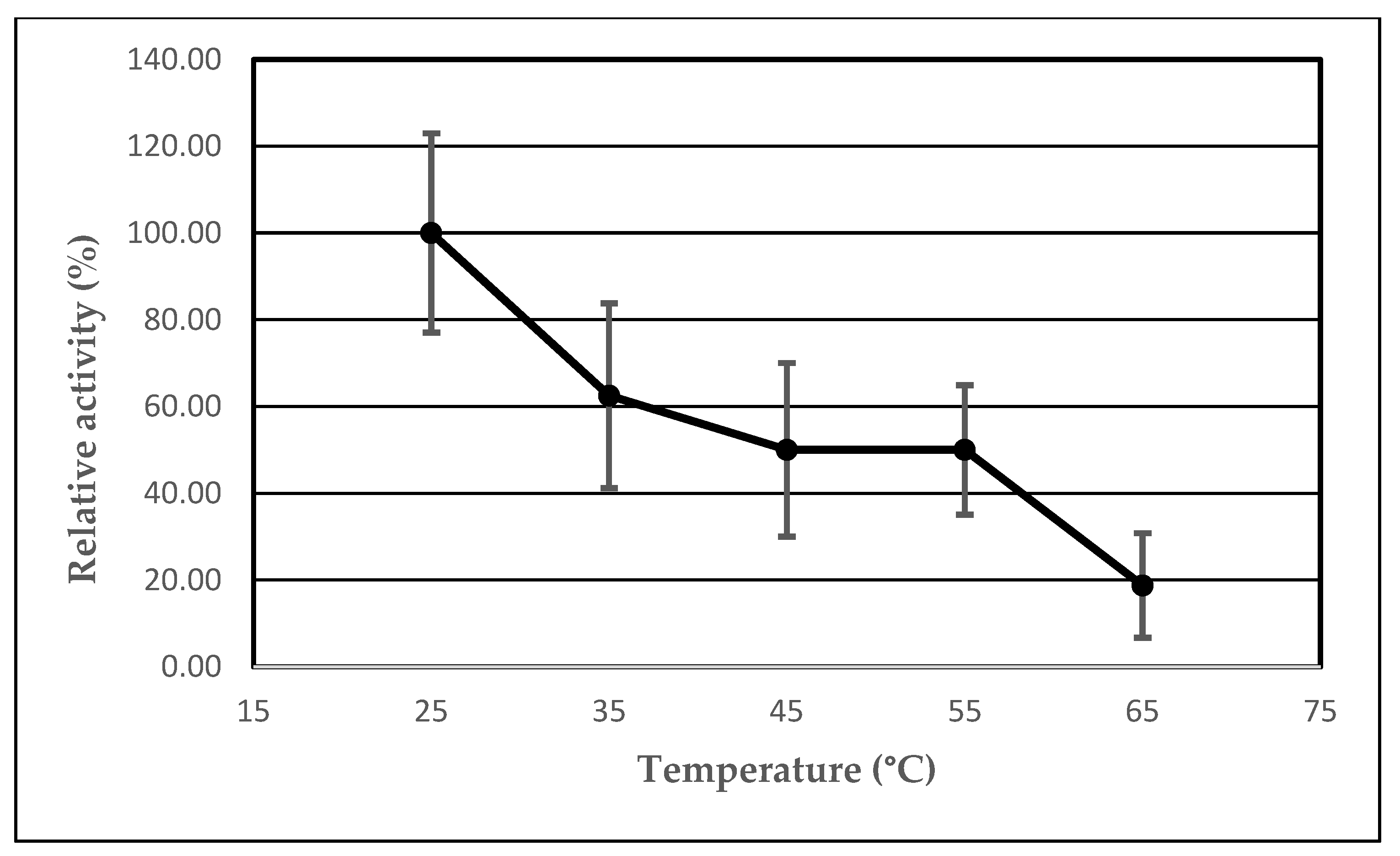
2.3. The Effect of Temperature on the Lipolytic Activity of the Immobilized AA-ANL
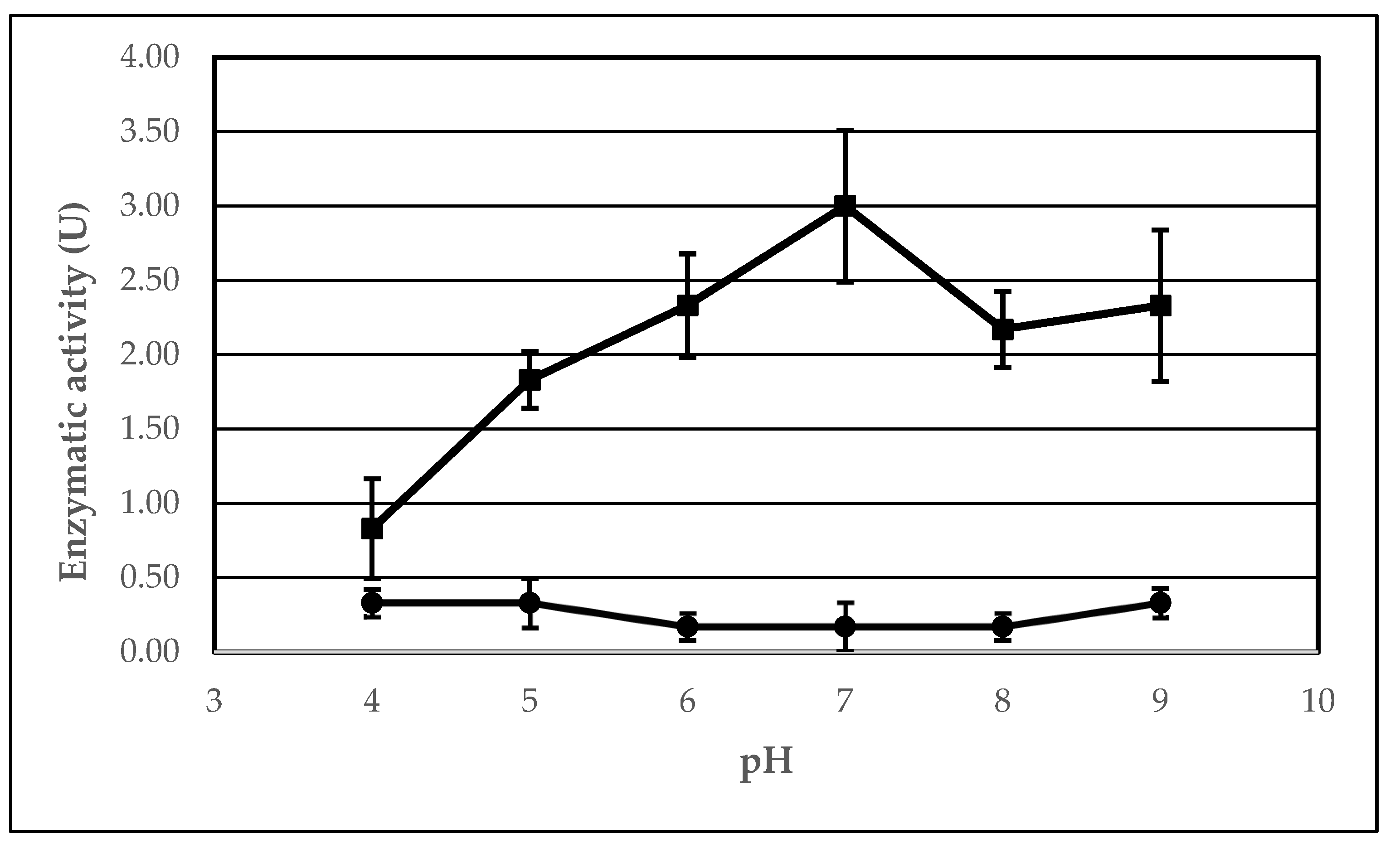
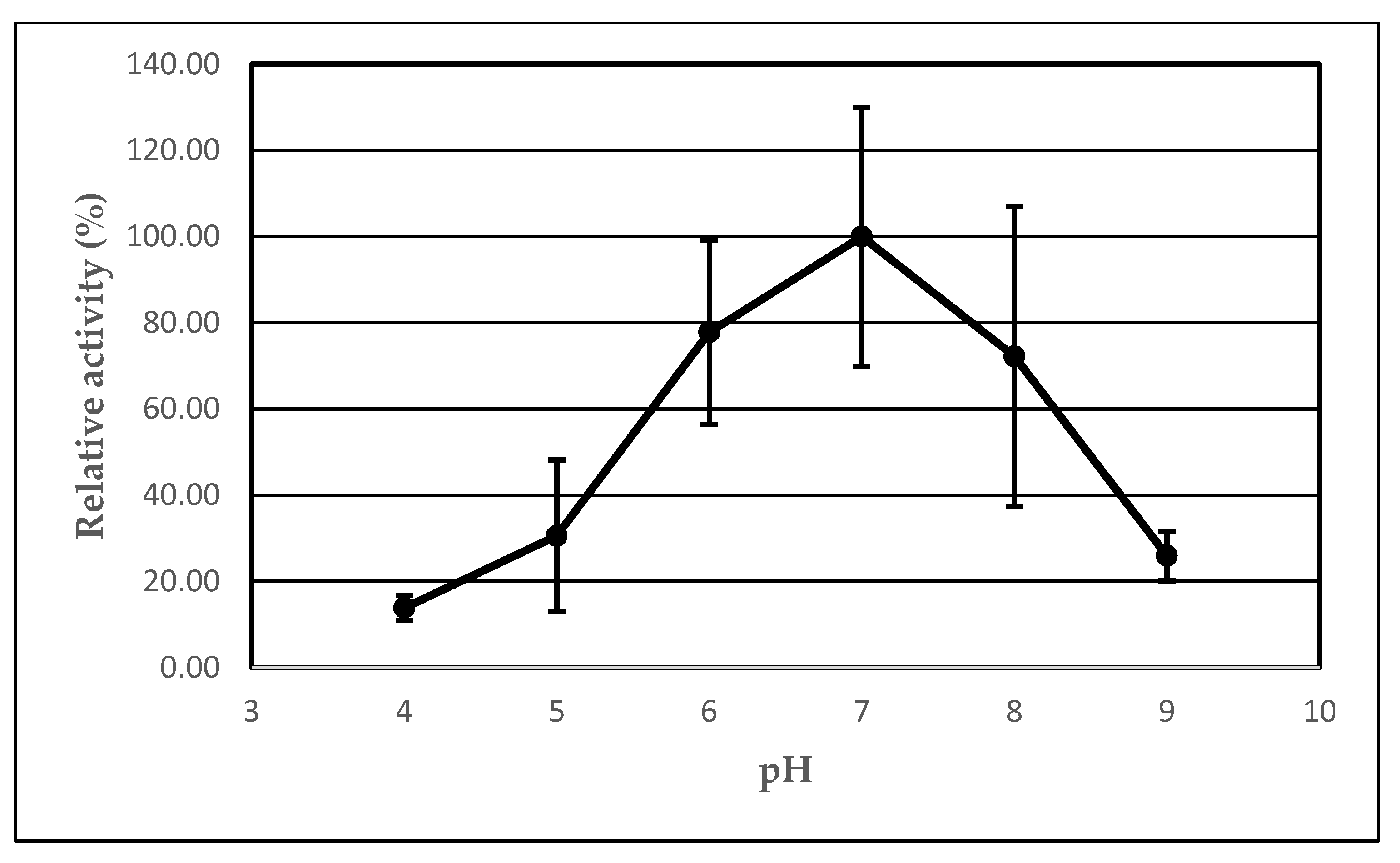
2.4. The Effect of pH on the Lipolytic Activity of Immobilized AA-ANL
2.5. The Effect of Substrate Mixtures on the Lipolytic Activity of Immobilized AA-ANL
2.6. Reuse of Immobilized AA-ANL
2.7. Storage Stability of the Immobilized AA-ANL
2.8. Kinetic Studies
3. Materials and Methods
3.1. Materials
3.2. Immobilization of AA-ANL onto Polyacrylic Supports IB-150A, IB-D152, and IB-EC1
3.3. Determination of the Amount of Immobilized AA-ANL
3.4. Determination of the Lipolytic Activity of Immobilized AA-ANL
3.5. Effect of the Support on Lipolytic Activity of Immobilized AA-ANL
3.6. Effect of Temperature and pH on the Lipolytic Activity of Immobilized AA-ANL
3.7. Effect of Substrate Mixtures on the Lipolytic Activity of Immobilized AA-ANL
3.8. Reusability of Immobilized AA-ANL
3.9. Storage Stability of Immobilized AA-ANL
3.10. Kinetic Studies
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Tacias-Pascacio, V.G.; Ortiz, C.; Rueda, N.; Berenguer-Murcia, A.; Acosta, N.; Aranaz, I.; Civera, C.; Fernandez-Lafuente, R.; Alcantara, A.R. Dextran Aldehyde in Biocatalysis: More Than a Mere Immobilization System. Catalysts 2019, 9, 622. [Google Scholar] [CrossRef]
- Alcalde, M.; Ferrer, M.; Plou, F.J.; Ballesteros, A. Environmental biocatalysis: From remediation with enzymes to novel green processes. Trends Biotechnol. 2006, 24, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Soumanou, M.M.; Bornscheuer, U.T.; Menge, U.; Schmid, R.D. Synthesis of structured triglycerides from peanut oil with immobilized lipase. J. Am. Oil Chem. 1997, 74, 427–433. [Google Scholar] [CrossRef]
- Busch, S.; Horlacher, P.; Both, S.; Westfechtel, A.; Schorken, U. Green synthesis routes toward triglycerides of conjugated linoleic acid. Eur. J. Lipid Sci. Technol 2011, 113, 92–99. [Google Scholar] [CrossRef]
- Siódmiak, T.; Ziegler-Borowska, M.; Marszałł, M.P. Lipase-immobilized magnetic chitosan nanoparticles for kinetic resolution of (R,S)-ibuprofen. J. Mol. Catal. B Enzym. 2013, 94, 7–14. [Google Scholar] [CrossRef]
- Marszałł, M.P.; Siódmiak, T. Immobilization of Candida rugosa lipase onto magnetic beads for kinetic resolution of (R,S)- ibuprofen. Catal. Commun. 2012, 24, 80–84. [Google Scholar] [CrossRef]
- Graffner-Nordberg, M.; Sjodin, K.; Tunek, A.; Hallberg, A. Synthesis and enzymatic hydrolysis of esters, constituting simple models of soft drugs. Chem. Pharm. Bull. 1998, 46, 591–601. [Google Scholar] [CrossRef]
- Song, X.; Qi, X.Y.; Hao, B.; Qu, Y.B. Studies of substrate specificities of lipases from different sources. Eur. J. Lipid Sci. Technol. 2008, 110, 1095–1101. [Google Scholar] [CrossRef]
- Peter, F.; Preda, G. Characterisation of pancreatic lipase substrate specificity in organic reaction media by a kinetic method. J. Mol. Catal. B Enzym. 2002, 19, 467–472. [Google Scholar] [CrossRef]
- Sanchez, D.A.; Tonetto, G.M.; Ferreira, M.L. Burkholderia cepacia lipase: A versatile catalyst in synthesis reactions. Biotechnol. Bioeng. 2018, 115, 6–24. [Google Scholar] [CrossRef]
- Stauch, B.; Fisher, S.J.; Cianci, M. Open and closed states of Candida antarctica lipase B: Protonation and the mechanism of interfacial activation. J. Lipid Res. 2015, 56, 2348–2358. [Google Scholar] [CrossRef] [PubMed]
- Zisis, T.; Freddolino, P.L.; Turunen, P.; van Teeseing, M.C.F.; Rowan, A.E.; Blank, K.G. Interfacial Activation of Candida antarctica Lipase B: Combined Evidence from Experiment and Simulation. Biochemistry 2015, 54, 5969–5979. [Google Scholar] [CrossRef] [PubMed]
- Secundo, F.; Carrea, G.; Tarabiono, C.; Gatti-Lafranconi, P.; Brocca, S.; Lotti, M.; Jaeger, K.E.; Puls, M.; Eggert, T. The lid is a structural and functional determinant of lipase activity and selectivity. J. Mol. Catal. B Enzym. 2006, 39, 166–170. [Google Scholar] [CrossRef]
- Chandra, P.; Enespa; Singh, R.; Arora, P.K. Microbial lipases and their industrial applications: A comprehensive review. Microb. Cell Fact. 2020, 19, 169. [Google Scholar] [CrossRef]
- Bezerra, C.S.; Lemos, C.; de Sousa, M.; Goncalves, L.R.B. Enzyme immobilization onto renewable polymeric matrixes: Past, present, and future trends. J. Appl. Polym. Sci. 2015, 132, 42125. [Google Scholar] [CrossRef]
- Siódmiak, T.; Mangelings, D.; Vander Heyden, Y.; Ziegler-Borowska, M.; Marszałł, M.P. High Enantioselective Novozym 435-Catalyzed Esterification of (R,S)-Flurbiprofen Monitored with a Chiral Stationary Phase. Appl. Biochem. Biotechnol. 2015, 175, 2769–2785. [Google Scholar] [CrossRef] [PubMed]
- Jose, C.; Toledo, M.V.; Briand, L.E. Enzymatic kinetic resolution of racemic ibuprofen: Past, present and future. Crit. Rev. Biotechnol. 2016, 36, 891–903. [Google Scholar] [CrossRef]
- Kourist, R.; de Maria, P.D.; Miyamoto, K. Biocatalytic strategies for the asymmetric synthesis of profens—recent trends and developments. Green Chem. 2011, 13, 2607–2618. [Google Scholar] [CrossRef]
- Carvalho, A.; Fonseca, T.D.; de Mattos, M.C.; de Oliveira, M.D.F.; de Lemos, T.L.G.; Molinari, F.; Romano, D.; Serra, I. Recent Advances in Lipase-Mediated Preparation of Pharmaceuticals and Their Intermediates. Int. J. Mol. Sci. 2015, 16, 29682–29716. [Google Scholar] [CrossRef]
- Rios, N.S.; Neto, D.M.A.; dos Santos, J.C.S.; Fechine, P.B.A.; Fernandez-Lafuente, R.; Goncalves, L.R.B. Comparison of the immobilization of lipase from Pseudomonas fluorescens on divinylsulfone or p-benzoquinone activated support. Int. J. Biol. Macromol. 2019, 134, 936–945. [Google Scholar] [CrossRef] [PubMed]
- Zdarta, J.; Sałek, K.; Kołodziejczak-Radzimska, A.; Siwińska-Stefańska, K.; Szwarc-Rzepka, K.; Norman, M.; Klapiszewski, Ł.; Bartczak, P.; Kaczorek, E.; Jesionowski, T. Immobilization of Amano Lipase A onto Stober silica surface: Process characterization and kinetic studies. Open Chem. 2015, 13, 138–148. [Google Scholar] [CrossRef]
- Le, Z.G.; Guo, L.T.; Jiang, G.F.; Yang, X.B.; Liu, H.Q. Henry reaction catalyzed by Lipase A from Aspergillus niger. Green Chem. Lett. Rev. 2013, 6, 277–281. [Google Scholar] [CrossRef]
- Yildiz, T. An Enzymatic and Environmentally Friendly Route for the Synthesis of Chiral beta-Hydroxy Ketones. ChemistrySelect 2019, 4, 7927–7931. [Google Scholar] [CrossRef]
- Yildiz, T.; Yasa, H.; Hasdemir, B.; Yusufoglu, A.S. Different bio/Lewis acid-catalyzed stereoselective aldol reactions in various mediums. Monatsh. Chem. 2017, 148, 1445–1452. [Google Scholar] [CrossRef]
- Dunne, A.; Palomo, J.M. Efficient and green approach for the complete deprotection of O-acetylated biomolecules. RSC Adv. 2016, 6, 88974–88978. [Google Scholar] [CrossRef]
- Xu, L.; Ke, C.X.; Huang, Y.; Yan, Y.J. Immobilized Aspergillus niger Lipase with SiO2 Nanoparticles in Sol-Gel Materials. Catalysts 2016, 6, 149. [Google Scholar] [CrossRef]
- Feng, K.L.; Huang, Z.C.; Peng, B.; Dai, W.J.; Li, Y.Q.; Zhu, X.A.; Chen, Y.J.; Tong, X.; Lan, Y.Q.; Cao, Y. Immobilization of Aspergillus niger lipase onto a novel macroporous acrylic resin: Stable and recyclable biocatalysis for deacidification of high-acid soy sauce residue oil. Bioresour. Technol. 2020, 298, 122553. [Google Scholar] [CrossRef]
- Contesini, F.J.; Calzado, F.; Madeira, J.V.; Rubio, M.V.; Zubieta, M.P.; de Melo, R.R.; Goncalves, T.A. Aspergillus Lipases: Biotechnological and Industrial Application. Fungal Metab. 2017, 639–666. [Google Scholar] [CrossRef]
- Silva, W.S.D.; Lapis, A.A.M.; Suarez, P.A.Z.; Neto, B.A.D. Enzyme-mediated epoxidation of methyl oleate supported by imidazolium-based ionic liquids. J. Mol. Catal. B Enzym. 2011, 68, 98–103. [Google Scholar] [CrossRef]
- Carvalho, P.D.; Contesini, F.J.; Ikegaki, M. Enzymatic resolution of (R,S)-ibuprofen and (R,S)-ketoprofen by microbial lipases from native and commercial sources. Braz. J. Microbiol. 2006, 37, 329–337. [Google Scholar] [CrossRef]
- El-Ghonemy, D.H.; Ali, T.H.; Hassanein, N.M.; Abdellah, E.M.; Fadel, M.; Awad, G.E.A.; Abdou, D.A.M. Thermo-alkali-stable lipase from a novel Aspergillus niger: Statistical optimization, enzyme purification, immobilization and its application in biodiesel production. Prep. Biochem. Biotechnol. 2021, 51, 225–240. [Google Scholar] [CrossRef] [PubMed]
- Cong, S.Z.; Tian, K.M.; Zhang, X.; Lu, F.P.; Singh, S.; Prior, B.; Wang, Z.X. Synthesis of flavor esters by a novel lipase from Aspergillus niger in a soybean-solvent system. 3 Biotech 2019, 9, 244. [Google Scholar] [CrossRef] [PubMed]
- Pera, L.M.; Romero, C.M.; Baigori, M.D.; Castro, G.R. Catalytic properties of lipase extracts from Aspergillus niger. Food Technol. Biotechnol. 2006, 44, 247–252. [Google Scholar]
- Qiao, H.Z.; Zhang, F.; Guan, W.T.; Zuo, J.J.; Feng, D.Y. Optimisation of combi-lipases from Aspergillus niger for the synergistic and efficient hydrolysis of soybean oil. J. Anim. Sci. 2017, 88, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Kuboki, A.; Ishihara, T.; Kobayashi, E.; Ohta, H.; Ishii, T.; Inoue, A.; Mitsuda, S.; Miyazaki, T.; Kajihara, Y.; Sugai, T. Synthesis of regioselectively protected forms of cytidine based on enzyme-catalyzed deacetylation as the key step. Biosci. Biotechnol. Biochem. 2000, 64, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Silano, V.; Baviera, J.M.B.; Bolognesi, C.; Bruschweiler, B.J.; Cocconcelli, P.S.; Crebelli, R.; Gott, D.M.; Grob, K.; Lampi, E.; Mortensen, A.; et al. Enzyme, Safety evaluation of the food enzyme triacylglycerol lipase from Aspergillus niger (strain LFS). Efsa J. 2019, 17, e05630. [Google Scholar] [PubMed]
- Carvalho, P.D.; Contesini, F.J.; Bizaco, R.; Calafatti, S.A.; Macedo, G.A. Optimization of enantioselective resolution of racemic ibuprofen by native lipase from Aspergillus niger. J. Ind. Microbiol. Biotechnol. 2006, 33, 713–718. [Google Scholar] [CrossRef]
- Thangaraj, B.; Solomon, P.R. Immobilization of Lipases—A Review. Part I: Enzyme Immobilization. ChemBioEng Rev. 2019, 6, 157–166. [Google Scholar] [CrossRef]
- Thangaraj, B.; Solomon, P.R. Immobilization of Lipases—A Review. Part II: Carrier Materials. ChemBioEng Rev. 2019, 6, 167–194. [Google Scholar] [CrossRef]
- Mokhtar, N.F.; Rahman, R.A.; Noor, N.D.M.; Shariff, F.M.; Ali, M.S.M. The Immobilization of Lipases on Porous Support by Adsorption and Hydrophobic Interaction Method. Catalysts 2020, 10, 744. [Google Scholar] [CrossRef]
- Boudrant, J.; Woodley, J.M.; Fernandez-Lafuente, R. Parameters necessary to define an immobilized enzyme preparation. Process Biochem. 2020, 90, 66–80. [Google Scholar] [CrossRef]
- Garcia-Galan, C.; Berenguer-Murcia, A.; Fernandez-Lafuente, R.; Rodrigues, R.C. Potential of Different Enzyme Immobilization Strategies to Improve Enzyme Performance. Adv. Synth. Catal. 2011, 353, 2885–2904. [Google Scholar] [CrossRef]
- Mateo, C.; Palomo, J.M.; Fernandez-Lorente, G.; Guisan, J.M.; Fernandez-Lafuente, R. Improvement of enzyme activity, stability and selectivity via immobilization techniques. Enzym. Microb. Technol. 2007, 40, 1451–1463. [Google Scholar] [CrossRef]
- Poppe, J.K.; Costa, A.P.O.; Brasil, M.C.; Rodrigues, R.C.; Ayub, M.A.Z. Multipoint covalent immobilization of lipases on aldehyde-activated support: Characterization and application in transesterification reaction. J. Mol. Catal. B Enzym. 2013, 94, 57–62. [Google Scholar] [CrossRef]
- Zucca, P.; Sanjust, E. Inorganic Materials as Supports for Covalent Enzyme Immobilization: Methods and Mechanisms. Molecules 2014, 19, 14139–14194. [Google Scholar] [CrossRef]
- Siódmiak, T.; Haraldsson, G.G.; Dulęba, J.; Ziegler-Borowska, M.; Siódmiak, J.; Marszałł, M.P. Evaluation of Designed Immobilized Catalytic Systems: Activity Enhancement of Lipase B from Candida antarctica. Catalysts 2020, 10, 876. [Google Scholar] [CrossRef]
- Alagoz, D.; Celik, A.; Yildirim, D.; Tukel, S.S.; Binay, B. Covalent immobilization of Candida methylica formate dehydrogenase on short spacer arm aldehyde group containing supports. J. Mol. Catal. B Enzym. 2016, 130, 40–47. [Google Scholar] [CrossRef]
- Matte, C.R.; Bussamara, R.; Dupont, J.; Rodrigues, R.C.; Hertz, P.F.; Ayub, M.A.Z. Immobilization of Thermomyces lanuginosus Lipase by Different Techniques on Immobead 150 Support: Characterization and Applications. Appl. Biochem. Biotechnol. 2014, 172, 2507–2520. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi, S.; Yousefi, M.; Nikseresht, A.; Omidi, H. Covalent binding and in-situ immobilization of lipases on a flexible nanoporous material. Process Biochem. 2021, 102, 92–101. [Google Scholar] [CrossRef]
- Elgharbawy, A.A.; Hayyan, A.; Hayyan, M.; Rashid, S.N.; Nor, M.R.M.; Zulkifli, M.Y.; Alias, Y.; Mirghani, M.E.S. Shedding Light on Lipase Stability in Natural Deep Eutectic Solvents. Chem. Biochem. Eng. Q. 2018, 32, 359–370. [Google Scholar] [CrossRef]
- Sharma, A.; Bhandari, K.; Jain, A.; Chaurasia, S.P.; Dalai, A.K. Lipase catalyzed esterification of Docosahexaenoic acid (DHA) with immobilized Pseudomonas cepacia and Thermomyces lanuginosus. Indian J. Chem. Technol. 2021, 28, 139–149. [Google Scholar]
- Diwan, B.; Gupta, P. Synthesis of MCFA and PUFA rich oils by enzymatic structuring of flax oil with single cell oils. LWT 2020, 133, 109928. [Google Scholar] [CrossRef]
- Hebda, P.; Wiśniowska, L.; Szafrański, P.W.; Cegła, M. Multigram-scale enzymatic kinetic resolution of trans-2-azidocyclohexyl acetate and chiral reversed-phase HPLC analysis of trans-2-azidocyclohexanol. Chirality 2022, 34, 428–437. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, A.L.L.; Assuncao, J.C.D.; Pascoal, C.V.P.; Bezerra, M.L.S.; Silva, A.C.S.; de Souza, B.V.; Rodrigues, F.E.A.; Ricardo, N.; Arruda, T. Waste of Nile Tilapia (Oreochromis niloticus) to Biodiesel Production by Enzymatic Catalysis.Optimization Using Factorial Experimental Design. Ind. Eng. Chem. Res. 2021, 60, 3554–3560. [Google Scholar] [CrossRef]
- Sanchez, D.A.; Alnoch, R.C.; Tonetto, G.M.; Krieger, N.; Ferreira, M.L. Immobilization and bioimprinting strategies to enhance the performance in organic medium of the metagenomic lipase LipC12. J. Biotechnol. 2021, 342, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Kahar, U.M.; Chan, K.G.; Sani, M.H.; Noh, N.I.M.; Goh, K.M. Effects of single and co-immobilization on the product specificity of type I pullulanase from Anoxybacillus sp SK3–4. Int. J. Biol. Macromol. 2017, 104, 322–332. [Google Scholar] [CrossRef]
- Gonzalez-Coronel, L.A.; Cobas, M.; Rostro-Alanis, M.D.; Parra-Saldivar, R.; Hernandez-Luna, C.; Pazos, M.; Sanroman, M.A. Immobilization of laccase of Pycnoporus sanguineus CS43. New Biotechnol. 2017, 39, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Gennari, A.; Mobayed, F.H.; Rafael, R.D.; Rodrigues, R.C.; Sperotto, R.A.; Volpato, G.; de Souza, C.F.V. Modification of Immobead 150 Support for Protein Immobilization: Effects on the Properties of Immobilized Aspergillus oryzae beta-Galactosidase. Biotechnol. Prog. 2018, 34, 934–943. [Google Scholar] [CrossRef]
- Binay, B.; Alagoz, D.; Yildirim, D.; Celik, A.; Tukel, S.S. Highly stable and reusable immobilized formate dehydrogenases: Promising biocatalysts for in situ regeneration of NADH. Beilstein J. Org. Chem. 2016, 12, 271–277. [Google Scholar] [CrossRef]
- Zdarta, J.; Jesionowski, T. Luffa cylindrica Sponges as a Thermally and Chemically Stable Support for Aspergillus niger Lipase. Biotechnol. Prog. 2016, 32, 657–665. [Google Scholar] [CrossRef]
- Dulęba, J.; Siódmiak, T.; Marszałł, M.P. Amano Lipase PS from Burkholderia cepacia—Evaluation of the Effect of Substrates and Reaction Media on the Catalytic Activity. Curr. Org. Chem. 2020, 24, 798–807. [Google Scholar] [CrossRef]
- Dulęba, J.; Siódmiak, T.; Marszałł, M.P. The influence of substrate systems on the enantioselective and lipolytic activity of immobilized Amano PS from Burkholderia cepacia lipase (APS-BCL). Process. Biochem. 2022, 130, 126–137. [Google Scholar] [CrossRef]
- Alagawany, M.; Elnesr, S.S.; Farag, M.R.; El-Sabrout, K.; Alqaisi, O.; Dawood, M.A.O.; Soomro, H.; Abdelnour, S.A. Nutritional significance and health benefits of omega-3,-6 and-9 fatty acids in animals. Anim. Biotechnol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Cholewski, M.; Tomczykowa, M.; Tomczyk, M. A Comprehensive Review of Chemistry, Sources and Bioavailability of Omega-3 Fatty Acids. Nutrients 2018, 10, 1662. [Google Scholar] [CrossRef] [PubMed]
- Dyerberg, J.; Madsen, P.; Moller, J.M.; Aardestrup, I.; Schmidt, E.B. Bioavailability of marine n-3 fatty acid formulations. Prostaglandins Leukot. Essent. Fatty Acids 2010, 83, 137–141. [Google Scholar] [CrossRef]
- Freeman, M.P.; Hibbeln, J.R.; Wisner, K.L.; Davis, J.M.; Mischoulon, D.; Peet, M.; Keck, P.E.; Mayangell, L.B.; Richardson, A.J.; Lake, J.; et al. Omega-3 fatty acids: Evidence basis for treatment and future research in psychiatry. J. Clin. Psychiatry 2006, 67, 1954–1967. [Google Scholar] [CrossRef]
- Curioni, C.C.; Alves, N.N.R.; Zago, L. Omega-3 supplementation in the treatment of overweight and obese children and adolescents: A systematic review. J. Funct. Foods 2019, 52, 340–347. [Google Scholar] [CrossRef]
- Das, U.N. Can Bioactive Lipids Inactivate Coronavirus (COVID-19)? Arch. Med. Res. 2020, 51, 282–286. [Google Scholar] [CrossRef]
- Yang, Z.Z.; Jin, W.H.; Cheng, X.Y.; Dong, Z.; Chang, M.; Wang, X.S. Enzymatic enrichment of n-3 polyunsaturated fatty acid glycerides by selective hydrolysis. Food Chem. 2021, 346, 128743. [Google Scholar] [CrossRef]
- Osadnik, K.; Jaworska, J. Analysis of ω-3 fatty acid content of polish fish oil drug and dietary supplements. Acta Pol. Pharm. 2016, 73, 875–883. [Google Scholar]
- Ciriminna, R.; Meneguzzo, F.; Delisi, R.; Pagliaro, M. Enhancing and improving the extraction of omega-3 from fish oil. Sustain. Chem. Pharm. 2017, 5, 54–59. [Google Scholar] [CrossRef]
- Hooper, L.; Al-Khudairy, L.; Abdelhamid, A.S.; Rees, K.; Brainard, J.S.; Brown, T.J.; Ajabnoor, S.M.; O’Brien, A.T.; Winstanley, L.E.; Donaldson, D.H.; et al. Omega-6 fats for the primary and secondary prevention of cardiovascular disease. Cochrane Database Syst. Rev. 2018, 7, CD011094. [Google Scholar] [PubMed]
- Halfen, S.; Jacometo, C.B.; Mattei, P.; Fenstenseifer, S.R.; Pfeifer, L.F.M.; Del Pino, F.A.B.; Santos, M.A.Z.; de Pereira, C.M.P.; Schmitt, E.; Correa, M.N. Diets Rich in Polyunsaturated Fatty Acids With Different Omega-6/Omega-3 Ratio Decrease Liver Content of Saturated Fatty Acids Across Generations of Wistar Rats. Braz. Arch. Biol. Technol. 2016, 59. [Google Scholar] [CrossRef]
- Zhang, Y.F.; Ge, J.; Liu, Z. Enhanced Activity of Immobilized or Chemically Modified Enzymes. ACS Catal. 2015, 5, 4503–4513. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Alagoz, D.; Tukel, S.S.; Yildirim, D. Immobilization of pectinase on silica-based supports: Impacts of particle size and spacer arm on the activity. Int. J. Biol. Macromol. 2016, 87, 426–432. [Google Scholar] [CrossRef]
- Ameri, A.; Asadi, F.; Shakibaie, M.; Forootanfar, H.; Ranjbar, M. Hydroxyapatite/Glycyrrhizin/Lithium-Based Metal-Organic Framework (HA/GL/Li-MOF) Nanocomposite as Support for Immobilization of Thermomyces lanuginosus Lipase. Appl. Biochem. Biotechnol. 2022, 194, 2108–2134. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Lorente, G.; Palomo, J.M.; Fuentes, M.; Mateo, C.; Guisan, J.M.; Fernandez-Lafuente, R. Self-assembly of Pseudomonas fluorescens lipase into bimolecular aggregates dramatically affects functional properties. Biotechnol. Bioeng. 2003, 82, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Gocen, T.; Bayari, S.H.; Guven, M.H. Effects of chemical structures of omega-6 fatty acids on the molecular parameters and quantum chemical descriptors. J. Mol. Struct. 2018, 1174, 142–150. [Google Scholar] [CrossRef]
- Akanbi, T.O.; Barrow, C.J. Candida antarctica lipase A effectively concentrates DHA from fish and thraustochytrid oils. Food Chem. 2017, 229, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.Y.; Smith, C.E.; Lai, C.Q.; Irvin, M.R.; Parnell, L.D.; Lee, Y.C.; Pham, L.D.; Aslibekyan, S.; Claas, S.A.; Tsai, M.Y.; et al. The effects of omega-3 polyunsaturated fatty acids and genetic variants on methylation levels of the interleukin-6 gene promoter. Mol. Nutr. Food Res. 2016, 60, 410–419. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, G.N.; Ramakrishnan, C.; Sasisekharan, V. Stereochemistry of polypeptide chain configurations. J. Mol. Biol. 1963, 7, 95–99. [Google Scholar] [CrossRef]
- Holt, A.P.; Bocharova, V.; Cheng, S.W.; Kisliuk, A.M.; White, B.T.; Saito, T.; Uhrig, D.; Mahalik, J.P.; Kumar, R.; Imel, A.E.; et al. Controlling Interfacial Dynamics: Covalent Bonding versus Physical Adsorption in Polymer Nanocomposites. ACS Nano 2016, 10, 6843–6852. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.N.; Jiang, Y.J.; Zhou, L.Y.; Gao, J. Immobilization of Candida antarctica lipase B by adsorption in organic medium. New Biotechnol. 2010, 27, 53–58. [Google Scholar] [CrossRef]
- dos Santos, E.A.L.; Lima, A.S.; Soares, C.M.F.; Santana, L. Lipase from Aspergillus niger obtained from mangaba residue fermentation: Biochemical characterization of free and immobilized enzymes on a sol-gel matrix. Acta Sci. Technol. 2017, 39, 1–8. [Google Scholar] [CrossRef]
- Muley, A.B.; Awasthi, S.; Bhalerao, P.P.; Jadhav, N.L.; Singhal, R.S. Preparation of cross-linked enzyme aggregates of lipase from Aspergillus niger: Process optimization, characterization, stability, and application for epoxidation of lemongrass oil. Bioprocess. Biosyst. Eng. 2021, 44, 1383–1404. [Google Scholar] [CrossRef] [PubMed]
- da Silva, V.C.F.; Contesini, F.J.; Carvalho, P.D. Characterization and Catalytic Activity of Free and Immobilized Lipase from Aspergillus niger: A Comparative Study. J. Braz. Chem. Soc. 2008, 19, 1468–1474. [Google Scholar] [CrossRef]
- Zubiolo, C.; Santos, R.C.A.; Carvalho, N.B.; Soares, C.M.F.; Lima, A.S.; Santana, L. Encapsulation in a sol-gel matrix of lipase from Aspergillus niger obtained by bioconversion of a novel agricultural residue. Bioprocess. Biosyst. Eng. 2014, 37, 1781–1788. [Google Scholar] [CrossRef]
- Singh, A.K.; Mukhopadhyay, M. Immobilization of lipase on carboxylic acid-modified silica nanoparticles for olive oil glycerolysis. Bioprocess. Biosyst. Eng. 2018, 41, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Yong, Y.; Bai, Y.X.; Li, Y.F.; Lin, L.; Cui, Y.J.; Xia, C.G. Characterization of Candida rugosa lipase immobilized onto magnetic microspheres with hydrophilicity. Process. Biochem. 2008, 43, 1179–1185. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oil | ω3/ω6 | ω6/ω9 | ω3/ω9 | ω6/ω3 | ω9/ω3 | ω9/ω6 |
|---|---|---|---|---|---|---|
| Peanut | - | 0.09 | - | - | - | 10.95 |
| Blackberry | 1.71 | - | - | 0.59 | - | - |
| Rapeseed | 0.47 | 0.29 | 0.13 | 2.13 | 7.47 | 3.50 |
| Pumpkin seed | - | 1.43 | - | - | - | 0.70 |
| Walnut | 0.59 | 1.00 | 0.59 | 1.70 | 1.70 | 1.00 |
| Sesame | - | 1.03 | - | - | - | 0.97 |
| Avocado | - | 0.19 | - | - | - | 5.31 |
| Rice | - | 0.73 | - | - | - | 1.38 |
| Corn | - | 1.75 | - | - | - | 0.57 |
| Black seed | - | 2.36 | - | - | - | 0.42 |
| Hemp | 0.31 | 4.64 | 1.45 | 3.19 | 0.69 | 0.22 |
| Safflower | - | 5.31 | - | - | - | 0.19 |
| Grape seed | - | 3.44 | - | - | - | 0.29 |
| Hazelnut | - | 0.12 | - | - | - | 8.09 |
| Evening primrose | - | 10.09 | - | - | - | 0.10 |
| Argan | - | 0.66 | - | - | - | 1.51 |
| Milk thistle | - | 2.04 | - | - | - | 0.49 |
| Borage | 0.02 | - | - | 58.00 | - | - |
| Apricot kernel | - | 0.38 | - | - | - | 2.61 |
| Olive | - | 0.10 | - | - | - | 9.71 |
| Fish | 6.67 | 0.60 | 4.00 | 0.15 | 0.25 | 1.67 |
| Sunflower | - | 0.04 | - | - | - | 24.21 |
| Linseed | 0.04 | - | - | 24.23 | - | - |
| Oils | LAB 1 (mg) | LL 2 (mg/g) | Iy 3 (%) | Ie 4 (%) | UB 5 (μmol/min) | UI 6 (μmol/min) |
|---|---|---|---|---|---|---|
| Peanut | 6.1 ± 0.1 | 122.0 ± 2.0 | 61.0 ± 1.0 | 250.00 | 0.67 ± 0.10 | 1.67 ± 0.10 |
| Blackberry | 5.8 ± 0.1 | 116.0 ± 2.0 | 58.0 ± 1.0 | 158.82 | 2.83 ± 0.17 | 4.50 ± 0.35 |
| Rapeseed | 5.9 ± 0.2 | 118.0 ± 4.0 | 59.0 ± 2.0 | 113.33 | 2.50 ± 0.10 | 2.83 ± 0.29 |
| Pumpkin seed | 4.0 ± 0.3 | 80.0 ± 6.0 | 40.0 ± 3.0 | 2400.00 | 0.17 ± 0.09 | 4.00 ± 0.25 |
| Walnut | 5.7 ± 0.1 | 114.0 ± 2.0 | 57.0 ± 1.0 | 440.00 | 0.83 ± 0.17 | 3.67 ± 0.42 |
| Sesame | 6.5 ± 0.1 | 130.0 ± 2.0 | 65.0 ± 1.0 | 566.67 | 0.50 ± 0.10 | 2.83 ± 0.25 |
| Avocado | 6.3 ± 0.2 | 126.0 ± 4.0 | 63.0 ± 2.0 | 428.57 | 1.17 ± 0.10 | 5.00 ± 0.17 |
| Rice | 4.5 ± 0.1 | 90.0 ± 2.0 | 45.0 ± 1.0 | 134.38 | 5.33 ± 0.51 | 7.17 ± 0.09 |
| Corn | 6.1 ± 0.1 | 122.0 ± 2.0 | 61.0 ± 1.0 | 1266.67 | 0.50 ± 0.10 | 6.33 ± 0.26 |
| Black cumin | 6.2 ± 0.1 | 124.0 ± 2.0 | 62.0 ± 1.0 | 240.00 | 1.67 ± 0.10 | 4.00 ± 0.29 |
| Hemp | 4.5 ± 0.2 | 90.0 ± 4.0 | 45.0 ± 2.0 | 516.67 | 1.00 ± 0.10 | 5.17 ± 0.17 |
| Safflower | 5.2 ± 0.2 | 104.0 ± 4.0 | 52.0 ± 2.0 | 377.78 | 1.50 ± 0.10 | 5.67 ± 0.25 |
| Grape seed | 5.0 ± 0.1 | 100.0 ± 2.0 | 50.0 ± 1.0 | 126.83 | 6.83 ± 0.35 | 8.67 ± 0.17 |
| Hazelnut | 4.4 ± 0.2 | 88.0 ± 4.0 | 44.0 ± 2.0 | 433.33 | 0.50 ± 0.10 | 2.17 ± 0.17 |
| Evening primrose | 4.4 ± 0.1 | 88.0 ± 2.0 | 44.0 ± 1.0 | 314.29 | 1.17 ± 0.17 | 3.67 ± 0.10 |
| Argan | 4.6 ± 0.2 | 92.0 ± 4.0 | 46.0 ± 2.0 | 600.00 | 0.50 ± 0.10 | 3.00 ± 0.17 |
| Milk thistle | 4.6 ± 0.2 | 92.0 ± 4.0 | 46.0 ± 2.0 | 343.75 | 2.67 ± 0.10 | 9.17 ± 0.25 |
| Borage | 4.8 ± 0.1 | 98.0 ± 2.0 | 48.0 ± 1.0 | 980.00 | 0.83 ± 0.09 | 8.17 ± 0.10 |
| Apricot kernel | 5.0 ± 0.1 | 100.0 ± 2.0 | 50.0 ± 1.0 | 310.00 | 1.67 ± 0.26 | 5.17 ± 0.35 |
| Olive | 4.5 ± 0.2 | 90.0 ± 4.0 | 45.0 ± 2.0 | 38.24 | 11.33 ± 0.25 | 4.33 ± 0.17 |
| Fish | 4.0 ± 0.3 | 80.0 ± 6.0 | 40.0 ± 3.0 | 1433.33 | 0.50 ± 0.10 | 7.17 ± 0.17 |
| Sunflower | 4.7 ± 0.1 | 94.0 ± 2.0 | 47.0 ± 1.0 | 120.00 | 1.67 ± 0.26 | 2.00 ± 0.25 |
| Linseed | 4.4 ± 0.2 | 88.0 ± 4.0 | 44.0 ± 2.0 | 175.00 | 1.17 ± 0.17 | 9.17 ± 0.10 |
| Support | Interaction Type | Functional Group | Molecular Size (μm) | LAB 1 (mg) | LL 2 (mg/g) | Iy 3 (%) | UB 4 (μmol/min) | UI 5 (μmol/min) | Ie 6 (%) |
|---|---|---|---|---|---|---|---|---|---|
| IB-150A | Covalent | Epoxide, apolar | 150–300 | 3.7 ± 0.1 | 74.0 ± 2.0 | 37.0 ± 1.0 | 0.17 ± 0.09 | 0.33 ± 0.09 | 200.00 |
| IB-D152 | Cationic | Carboxylic acid | 350–700 | 1.3 ± 0.1 | 26.0 ± 2.0 | 13.0 ± 1.0 | 1.00 ± 0.17 | 2.33 ± 0.10 | 233.00 |
| IB-EC1 | Non-ionic, adsorption | Carboxylic ester | 350–700 | 5.0 ± 0.1 | 100.0 ± 2.0 | 50.0 ± 1.0 | 0.17 ± 0.09 | 0.17 ± 0.09 | 100.00 |
| Temperature (°C) | Ie 1 (%) | UB 2 (μmol/min) | UI 3 (μmol/min) | Arel 4 (%) |
|---|---|---|---|---|
| 25 | 800.00 | 0.17 ± 0.09 | 1.33 ± 0.21 | 100.00 ± 22.95 |
| 35 | 500.00 | 0.33 ± 0.09 | 1.67 ± 0.42 | 62.50 ± 21.31 |
| 45 | 400.00 | 0.67 ± 0.10 | 2.67 ± 0.17 | 50.00 ± 20.01 |
| 55 | 400.00 | 0.17 ± 0.09 | 0.67 ± 0.17 | 50.00 ± 14.93 |
| 65 | 150.00 | 0.67 ± 0.10 | 1.00 ± 0.16 | 18.75 ± 12.04 |
| pH | Ie 1 (%) | UB 2 (μmol/min) | UI 3 (μmol/min) | Arel 4 (%) |
|---|---|---|---|---|
| 4 | 250.00 | 0.33 ± 0.10 | 0.83 ± 0.33 | 13.89 ± 2.93 |
| 5 | 550.00 | 0.33 ± 0.17 | 1.83 ± 0.19 | 30.56 ± 17.65 |
| 6 | 1400.00 | 0.17 ± 0.09 | 2.33 ± 0.35 | 77.78 ± 21.33 |
| 7 | 1800.00 | 0.17 ± 0.09 | 3.00 ± 0.51 | 100.00 ± 30.02 |
| 8 | 1300.00 | 0.17 ± 0.09 | 2.17 ± 0.25 | 72.22 ± 34.74 |
| 9 | 700.00 | 0.33 ± 0.10 | 2.33 ± 0.51 | 38.89 ± 5.79 |
| Oil Mixture 1:1 | LAB 1 | LL 2 | Iy 3 (%) | Ie 4 (%) | UB 5 (μmol/min) | UI 6 (μmol/min) |
|---|---|---|---|---|---|---|
| (a) Milk thistle + Sesame (+) | 5.6 ± 0.1 | 112.0 ± 2.0 | 56.0 ± 1.0 | 1800.00 | 0.17 ± 0.10 | 3.00 ± 0.10 |
| (b) Sunflower + Avocado (+/−) | 5.7 ± 0.1 | 114.0 ± 2.0 | 57.0 ± 1.0 | 42.00 | 9.17 ± 0.26 | 3.83 ± 0.33 |
| (c) Milk thistle + Sunflower (+/−) | 5.8 ± 0.1 | 116.0 ± 2.0 | 58.0 ± 1.0 | 400.00 | 1.00 ± 0.25 | 4.00 ± 0.15 |
| (d) Pumpkin seed + Fish (+ +) | 5.5 ± 0.1 | 110.0 ± 2.0 | 55.0 ± 1.0 | 1600.00 | 0.17 ± 0.10 | 2.67 ± 0.26 |
| (e) Olive + Rapeseed (−) | 5.5 ± 0.1 | 110.0 ± 2.0 | 55.0 ± 1.0 | 500.00 | 0.33 ± 0.09 | 1.67 ± 0.09 |
| (f) Pumpkin seed + Olive (+ +/−) | 5.3 ± 0.1 | 106.0 ± 2.0 | 53.0 ± 1.0 | 1400.00 | 0.17 ± 0.10 | 2.33 ± 0.16 |
| Support Reuse Cycle | UB 1 (μmol/min) | UI 2 (μmol/min) | Ie 3 (%) | Arel 4 (%) |
|---|---|---|---|---|
| 1 | 1.83 ± 0.19 | 4.33 ± 0.25 | 236.00 | 100.00 ± 0.06 |
| 2 | n/a | 4.06 ± 0.18 | 221.00 | 94.00 ± 1.36 |
| 3 | n/a | 3.78 ± 0.22 | 206.00 | 87.00 ± 10.59 |
| 4 | n/a | 3.78 ± 0.12 | 206.00 | 87.00 ± 3.78 |
| Storage Time (h) | UB 1 (μmol/min) | UI 2 (μmol/min) | Ie 3 (%) |
|---|---|---|---|
| 24 | 2.50 ± 0.10 | 4.67 ± 0.10 | 187.00 |
| 168 | 2.17 ± 0.17 | 0.50 ± 0.17 | 23.00 |
| Lipase | Km (mg/mL) | Vmax (U/mg) | kcat (s−1) | R2 |
|---|---|---|---|---|
| AA-ANL | 29.20 ± 0.08 | 0.061 ± 0.005 | 0.003 ± 0.001 | 0.933 |
| AA-ANL-IB-150A | 1827.95 ± 0.17 | 1.648 ± 0.019 | 0.082 ± 0.006 | 0.986 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siódmiak, T.; Dulęba, J.; Kocot, N.; Wątróbska-Świetlikowska, D.; Marszałł, M.P. The High ‘Lipolytic Jump’ of Immobilized Amano A Lipase from Aspergillus niger in Developed ‘ESS Catalytic Triangles’ Containing Natural Origin Substrates. Catalysts 2022, 12, 853. https://doi.org/10.3390/catal12080853
Siódmiak T, Dulęba J, Kocot N, Wątróbska-Świetlikowska D, Marszałł MP. The High ‘Lipolytic Jump’ of Immobilized Amano A Lipase from Aspergillus niger in Developed ‘ESS Catalytic Triangles’ Containing Natural Origin Substrates. Catalysts. 2022; 12(8):853. https://doi.org/10.3390/catal12080853
Chicago/Turabian StyleSiódmiak, Tomasz, Jacek Dulęba, Natalia Kocot, Dorota Wątróbska-Świetlikowska, and Michał Piotr Marszałł. 2022. "The High ‘Lipolytic Jump’ of Immobilized Amano A Lipase from Aspergillus niger in Developed ‘ESS Catalytic Triangles’ Containing Natural Origin Substrates" Catalysts 12, no. 8: 853. https://doi.org/10.3390/catal12080853