
Bacterial Competition for the Anode Colonization under Different External Resistances in Microbial Fuel Cells



Abstract
:1. Introduction
2. Results
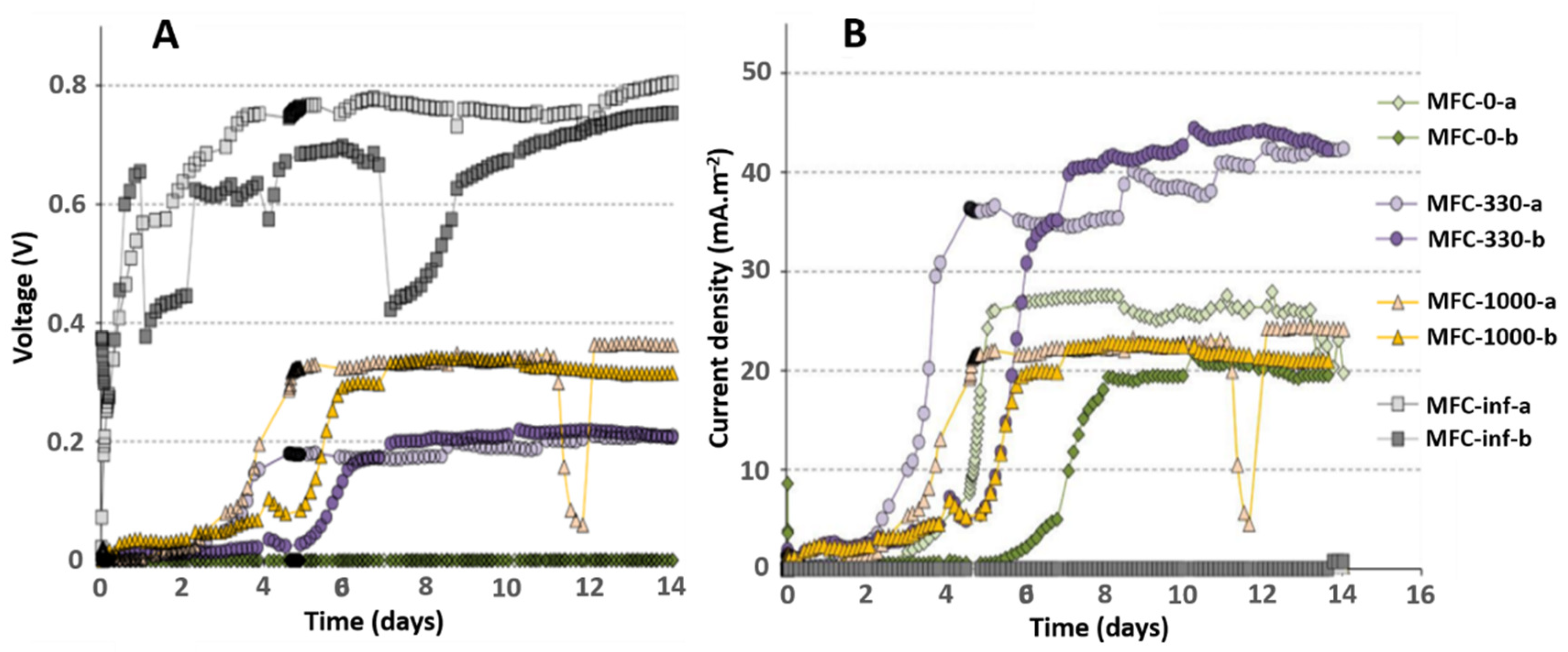
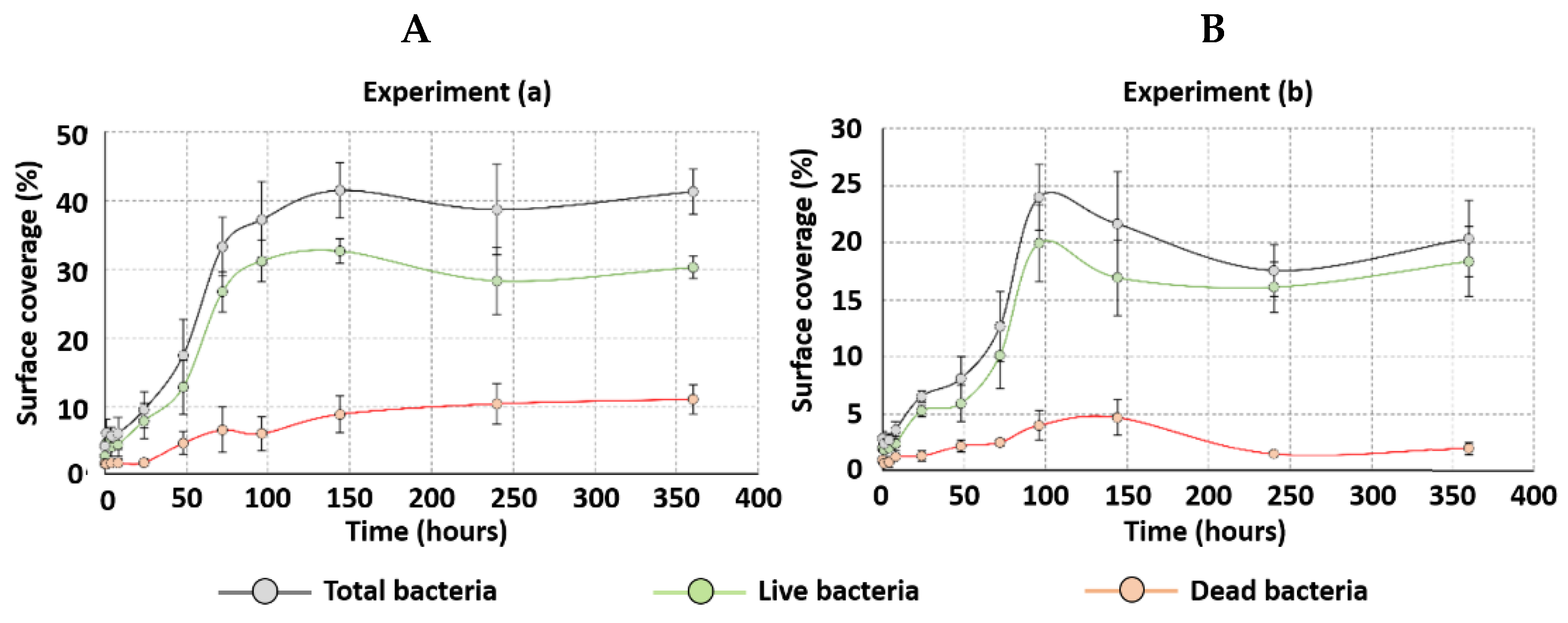
2.1. Impact of External Resistance on Anodic Biofilm Growth
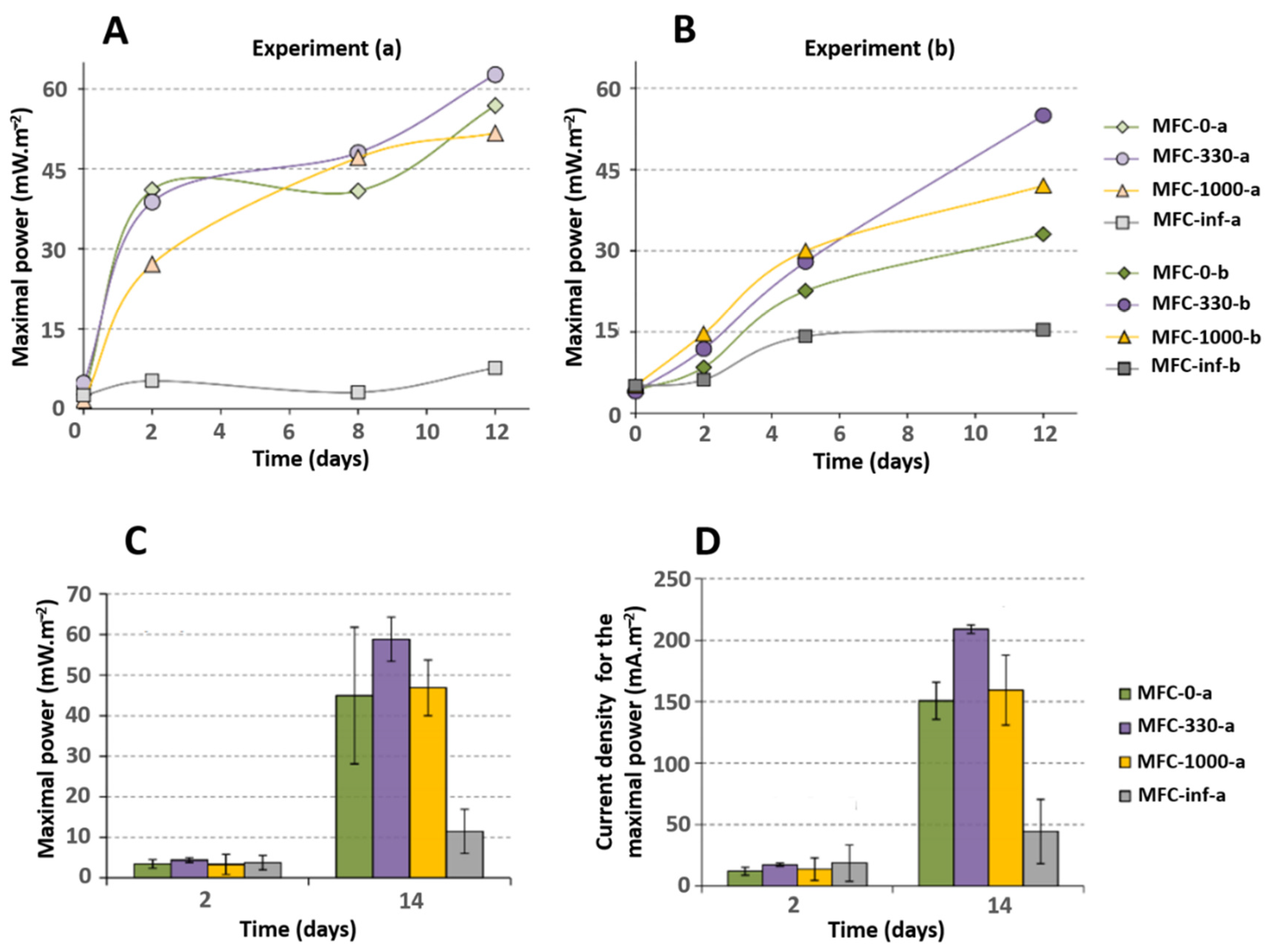
2.2. Impact of External Resistance on Electrical Performance of MFCs
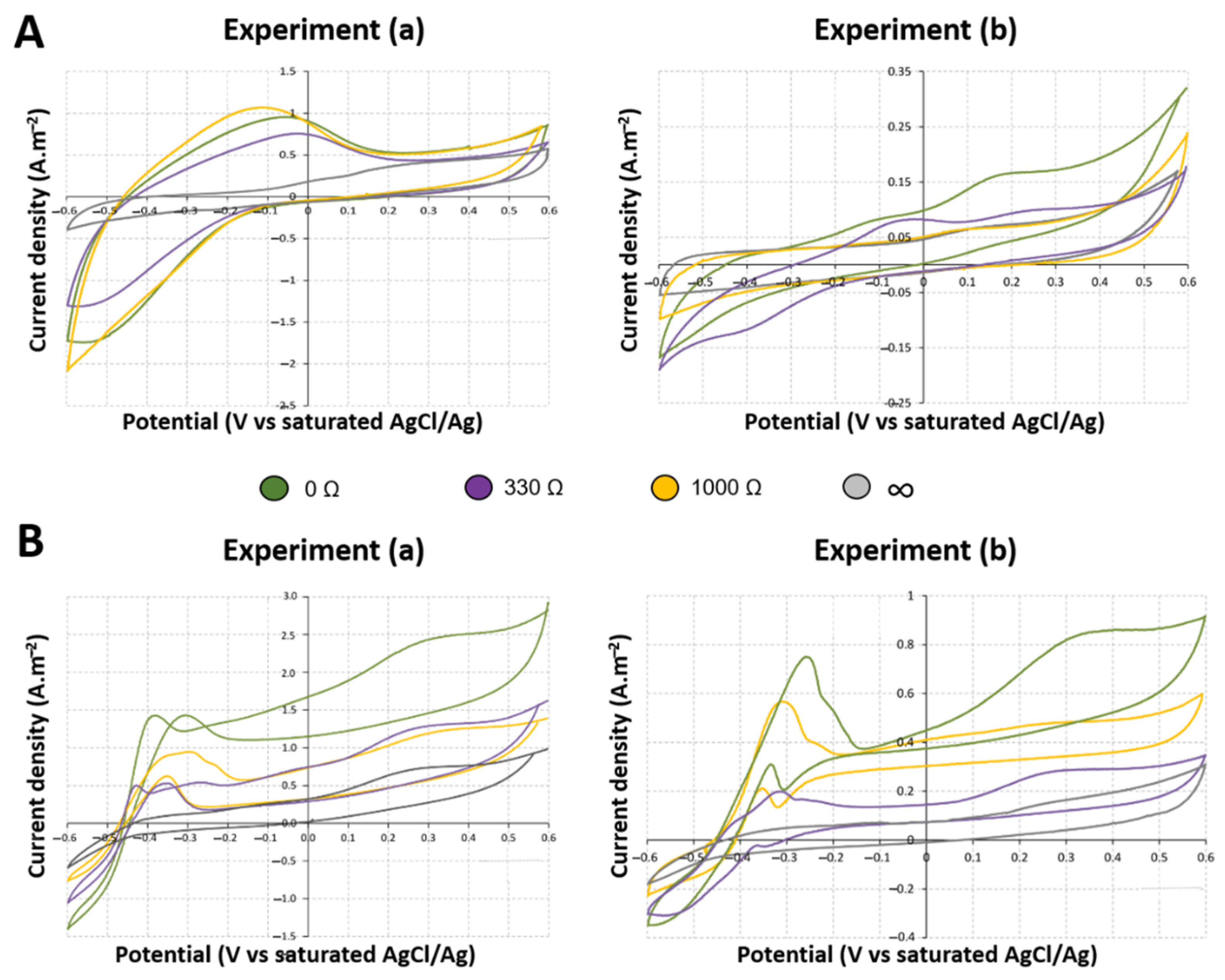
2.3. Influence on Electrochemical Activity of Anodic Biofilms
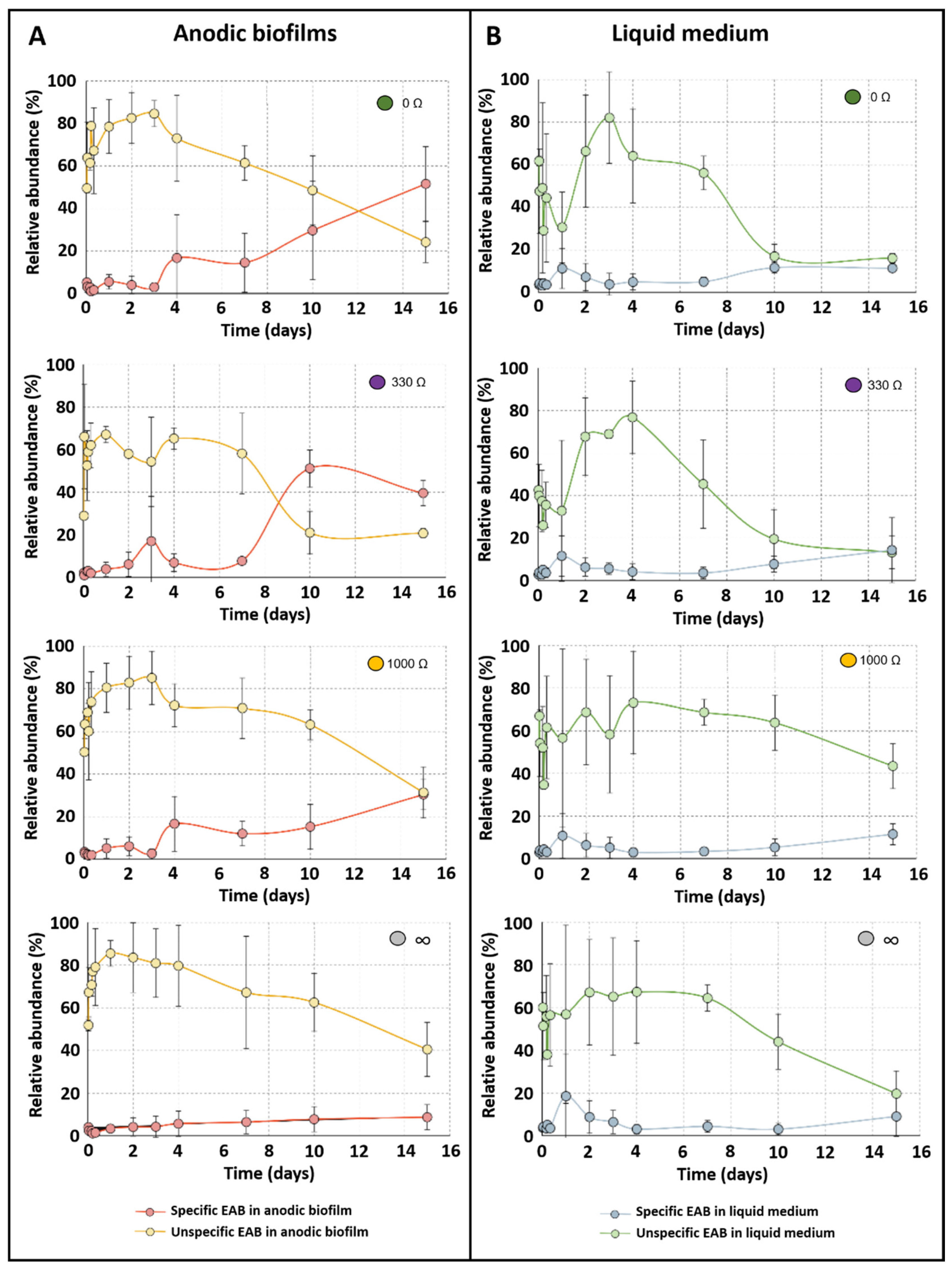
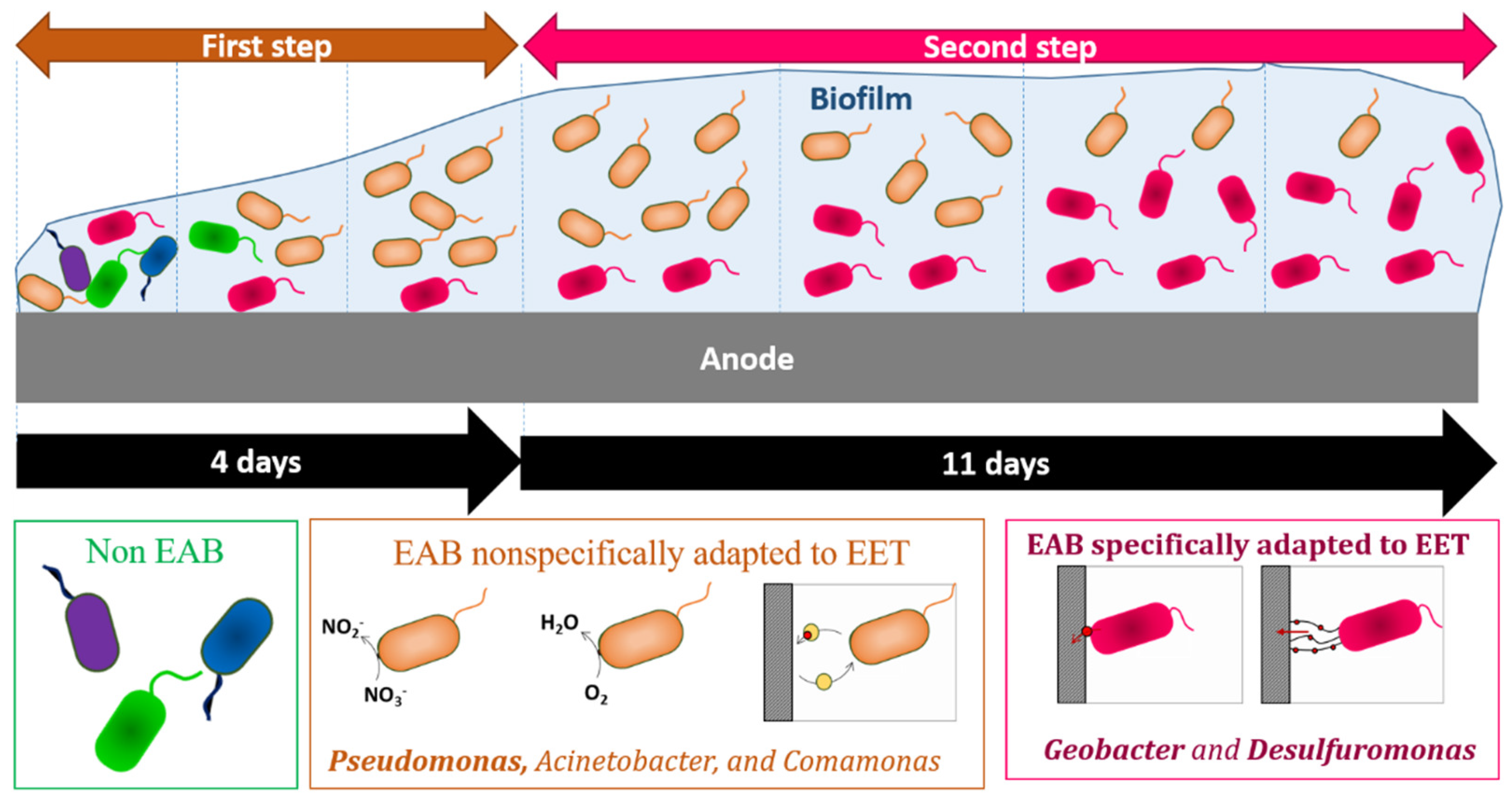
2.4. Impact of External Resistance on Bacterial Community Dynamic of Anodic Biofilms
3. Discussion
4. Materials and Methods
4.1. MFC Steup and Sampling
4.2. Electrochemical Measurements
4.3. Microscopic Observations and Image Analyses
4.4. Sequencing Analyses
4.5. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Saadi, M.; Pézard, J.; Haddour, N.; Erouel, M.; Vogel, T.M.; Khirouni, K. Stainless Steel Coated with Carbon Nanofiber/PDMS Composite as Anodes in Microbial Fuel Cells. Mater. Res. Express 2020, 7, 25504. [Google Scholar] [CrossRef]
- Bensalah, F.; Julien, P.; Haddour, N.; Erouel, M.; Buret, F.; Khirouni, K. Carbon Nano-Fiber/PDMS Composite Used as Corrosion-Resistant Coating for Copper Anodes in Microbial Fuel Cells. Nanomaterials 2021, 11, 3144. [Google Scholar] [CrossRef]
- Jia, H.; Yang, G.; Wang, J.; Ngo, H.H.; Guo, W.; Zhang, H.; Zhang, X. Performance of a Microbial Fuel Cell-Based Biosensor for Online Monitoring in an Integrated System Combining Microbial Fuel Cell and Upflow Anaerobic Sludge Bed Reactor. Bioresour. Technol. 2016, 218, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Lai, B.; Tang, X. Microbial Fuel Cell-Based Biosensors. Biosensors 2019, 9, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, N.; Yan, Z.; Xu, H.; Yao, Z.; Wang, C.; Chen, M.; Zhao, Z.; Peng, Z.; Wang, C.; Jiang, H.L. Development of a Sediment Microbial Fuel Cell-Based Biosensor for Simultaneous Online Monitoring of Dissolved Oxygen Concentrations along Various Depths in Lake Water. Sci. Total Environ. 2019, 673, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Prévoteau, A.; Rabaey, K. Electroactive Biofilms for Sensing: Reflections and Perspectives. ACS Sens. 2017, 2, 1072–1085. [Google Scholar] [CrossRef] [PubMed]
- Pinck, S.; Ostormujof, L.M.; Teychené, S.; Erable, B. Microfluidic Microbial Bioelectrochemical Systems: An Integrated Investigation Platform for a More Fundamental Understanding of Electroactive Bacterial Biofilms. Microorganisms 2020, 8, 1841. [Google Scholar] [CrossRef] [PubMed]
- Greenman, J.; Gajda, I.; You, J.; Mendis, B.A.; Obata, O.; Pasternak, G.; Ieropoulos, I. Microbial Fuel Cells and Their Electrified Biofilms. Biofilm 2021, 3, 100057. [Google Scholar] [CrossRef] [PubMed]
- Lyon, D.Y.; Buret, F.; Vogel, T.M.; Monier, J.M. Is Resistance Futile? Changing External Resistance Does Not Improve Microbial Fuel Cell Performance. Bioelectrochemistry 2010, 78, 2–7. [Google Scholar] [CrossRef]
- Rismani-Yazdi, H.; Christy, A.D.; Carver, S.M.; Yu, Z.; Dehority, B.A.; Tuovinen, O.H. Effect of External Resistance on Bacterial Diversity and Metabolism in Cellulose-Fed Microbial Fuel Cells. Bioresour. Technol. 2011, 102, 278–283. [Google Scholar] [CrossRef]
- Jung, S.; Regan, J.M. Influence of External Resistance on Electrogenesis, Methanogenesis, and Anode Prokaryotic Communities in Microbial Fuel Cells. Appl. Environ. Microbiol. 2011, 77, 564–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koók, L.; Nemestóthy, N.; Bélafi-Bakó, K.; Bakonyi, P. Investigating the Specific Role of External Load on the Performance versus Stability Trade-off in Microbial Fuel Cells. Bioresour. Technol. 2020, 309, 123313. [Google Scholar] [CrossRef] [PubMed]
- Katuri, K.P.; Scott, K.; Head, I.M.; Picioreanu, C.; Curtis, T.P. Microbial Fuel Cells Meet with External Resistance. Bioresour. Technol. 2011, 102, 2758–2766. [Google Scholar] [CrossRef] [PubMed]
- Pinto, R.P.; Srinivasan, B.; Uiot, S.R.; Tartakovsky, B. The Effect of Real-Time External Resistance Optimization on Microbial Fuel Cell Performance. Water Res. 2011, 45, 1571–1578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Zhu, X.; Li, J.; Liao, Q.; Ye, D. Biofilm Formation and Electricity Generation of a Microbial Fuel Cell Started up under Different External Resistances. J. Power Sources 2011, 196, 6029–6035. [Google Scholar] [CrossRef]
- Vilajeliu-Pons, A.; Bañeras, L.; Puig, S.; Molognoni, D.; Vilà-Rovira, A.; Del Amo, E.H.; Balaguer, M.D.; Colprim, J. External Resistances Applied to MFC Affect Core Microbiome and Swine Manure Treatment Efficiencies. PLoS ONE 2016, 11, e0164044. [Google Scholar] [CrossRef] [PubMed]
- Kamau, J.M.; Mbui, D.N.; Mwaniki, J.M.; Mwaura, F.B.; Kamau, G.N. Microbial Fuel Cells: Influence of External Resistors on Power, Current and Power Density. J. Thermodyn. Catal. 2017, 8, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Rossi, R.; Logan, B.E. Impact of External Resistance Acclimation on Charge Transfer and Diffusion Resistance in Bench-Scale Microbial Fuel Cells. Bioresour. Technol. 2020, 318, 123921. [Google Scholar] [CrossRef]
- Paitier, A.; Godain, A.; Lyon, D.; Haddour, N.; Vogel, T.M.; Monier, J.M. Microbial Fuel Cell Anodic Microbial Population Dynamics during MFC Start-Up. Biosens. Bioelectron. 2017, 92, 357–363. [Google Scholar] [CrossRef]
- Hodgson, D.M.; Smith, A.; Dahale, S.; Stratford, J.P.; Li, J.V.; Grüning, A.; Bushell, M.E.; Marchesi, J.R.; Avignone Rossa, C. Segregation of the Anodic Microbial Communities in a Microbial Fuel Cell Cascade. Front. Microbiol. 2016, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Pasternak, G.; Greenman, J.; Ieropoulos, I. Dynamic Evolution of Anodic Biofilm When Maturing under Different External Resistive Loads in Microbial Fuel Cells. Electrochemical Perspective. J. Power Sources 2018, 400, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Freguia, S.; Rabaey, K.; Yuan, Z.; Keller, J. Electron and Carbon Balances in Microbial Fuel Cells Reveal Temporary Bacterial Storage Behavior during Electricity Generation. Environ. Sci. Technol. 2007, 41, 2915–2921. [Google Scholar] [CrossRef] [PubMed]
- Aelterman, P.; Versichele, M.; Marzorati, M.; Boon, N.; Verstraete, W. Loading Rate and External Resistance Control the Electricity Generation of Microbial Fuel Cells with Different Three-Dimensional Anodes. Bioresour. Technol. 2008, 99, 8895–8902. [Google Scholar] [CrossRef] [PubMed]
- Roy, J.N.; Babanova, S.; Garcia, K.E.; Cornejo, J.; Ista, L.K.; Atanassov, P. Electrochimica Acta Catalytic Biofilm Formation by Shewanella Oneidensis MR-1 and Anode Characterization by Expanded Uncertainty. Electrochim. Acta 2014, 126, 3–10. [Google Scholar] [CrossRef]
- Oziat, J.; Cohu, T.; Elsen, S.; Gougis, M.; Malliaras, G.G.; Mailley, P. Electrochemical Detection of Redox Molecules Secreted by Pseudomonas Aeruginosa—Part 1: Electrochemical Signatures of Different Strains. Bioelectrochemistry 2021, 140, 107747. [Google Scholar] [CrossRef]
- Xiao, Y.; Zheng, Y.; Wu, S.; Zhang, E.H.; Chen, Z.; Liang, P.; Huang, X.; Yang, Z.H.; Ng, I.S.; Chen, B.Y.; et al. Pyrosequencing Reveals a Core Community of Anodic Bacterial Biofilms in Bioelectrochemical Systems from China. Front. Microbiol. 2015, 6, 1410. [Google Scholar] [CrossRef] [Green Version]
- Pinto, D.; Coradin, T.; Laberty-Robert, C. Effect of Anode Polarization on Biofilm Formation and Electron Transfer in Shewanella Oneidensis/Graphite Felt Microbial Fuel Cells. Bioelectrochemistry 2018, 120, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Jung, S. Impedance Analysis of Geobacter Sulfurreducens PCA, Shewanella Oneidensis MR-1, and Their Coculture in Bioeletrochemical Systems. Int. J. Electrochem. Sci. 2012, 7, 11091–11100. [Google Scholar]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R.; Al, W.E.T. Naïve Bayesian Classifier for Rapid Assignment of RRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Cole, J.R.; Wang, Q.; Fish, J.A.; Chai, B.; Mcgarrell, D.M.; Sun, Y.; Brown, C.T.; Porras-alfaro, A.; Kuske, C.R.; Tiedje, J.M. Ribosomal Database Project: Data and Tools for High Throughput RRNA Analysis. Nucleic Acids Res. 2014, 42, 633–642. [Google Scholar] [CrossRef] [Green Version]
- Kiely, P.D.; Call, D.F.; Yates, M.D.; Regan, J.M.; Logan, B.E. Anodic Biofilms in Microbial Fuel Cells Harbor Low Numbers of Higher-Power-Producing Bacteria than Abundant Genera. Appl. Microbiol. Biotechnol. 2010, 88, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wei, J.; Liang, P.; Huang, X. Bioresource Technology Electricity Generation and Microbial Community Changes in Microbial Fuel Cells Packed with Different Anodic Materials. Bioresour. Technol. 2011, 102, 10886–10891. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.H.; Boon, N.; Maeyer, K. De Use of Pseudomonas Species Producing Phenazine-Based Metabolites in the Anodes of Microbial Fuel Cells to Improve Electricity Generation. Appl. Microbiol. Biotechnol. 2008, 80, 985–993. [Google Scholar] [CrossRef] [PubMed]
- Freguia, S.; Tsujimura, S.; Kano, K. Electrochimica Acta Electron Transfer Pathways in Microbial Oxygen Biocathodes. Electrochim. Acta 2010, 55, 813–818. [Google Scholar] [CrossRef]
- Kievit, T.R. De Minireview Quorum Sensing in Pseudomonas Aeruginosa Biofilms. Environ. Microbiol. 2009, 11, 279–288. [Google Scholar] [CrossRef]
- Dietrich, L.E.P.; Price-whelan, A.; Petersen, A.; Whiteley, M.; Newman, D.K. The Phenazine Pyocyanin Is a Terminal Signalling Factor in the Quorum Sensing Network of Pseudomonas Aeruginosa. Mol. Microbiol. 2006, 61, 1308–1321. [Google Scholar] [CrossRef]
- Chen, S.; Jing, X.; Tang, J.; Fang, Y.; Zhou, S. Biosensors and Bioelectronics Quorum Sensing Signals Enhance the Electrochemical Activity and Energy Recovery of Mixed-Culture Electroactive Biofilms. Biosens. Bioelectron. 2017, 97, 369–376. [Google Scholar] [CrossRef]
- Cheng, S.; Liu, H.; Logan, B.E. Increased Performance of Single-Chamber Microbial Fuel Cells Using an Improved Cathode Structure. Electrochem. Commun. 2006, 8, 489–494. [Google Scholar] [CrossRef]
- Kang, H.; Jeong, J.; Gupta, P.L.; Jung, S.P. Effects of Brush-Anode Configurations on Performance and Electrochemistry of Microbial Fuel Cells. Int. J. Hydrogen Energy 2017, 42, 27693–27700. [Google Scholar] [CrossRef]
- Kang, H.; Kim, E.; Jung, S.P. Influence of Flowrates to a Reverse Electro-Dialysis (RED) Stack on Performance and Electrochemistry of a Microbial Reverse Electrodialysis Cell (MRC). Int. J. Hydrogen Energy 2017, 42, 27685–27692. [Google Scholar] [CrossRef]
- Nam, T.; Son, S.; Koo, B.; Hoa Tran, H.V.; Kim, J.R.; Choi, Y.; Jung, S.P. Comparative Evaluation of Performance and Electrochemistry of Microbial Fuel Cells with Different Anode Structures and Materials. Int. J. Hydrogen Energy 2017, 42, 27677–27684. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MFC | Relative Abundance of Specific EAB (%) | Genus Number | Shannon Index |
|---|---|---|---|
| MFC-0 | 51 ± 17 | 123 ± 8 | 2.09 ± 0.40 |
| MFC-330 | 39 ± 5 | 127 ± 9 | 2.54 ± 0.12 |
| MFC-1000 | 30 ± 7 | 138 ± 17 | 2.76 ± 0.40 |
| MFC-inf | 9 ± 6 | 163 ± 13 | 3.20 ± 0.23 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Godain, A.; Haddour, N.; Fongarland, P.; Vogel, T.M. Bacterial Competition for the Anode Colonization under Different External Resistances in Microbial Fuel Cells. Catalysts 2022, 12, 176. https://doi.org/10.3390/catal12020176
Godain A, Haddour N, Fongarland P, Vogel TM. Bacterial Competition for the Anode Colonization under Different External Resistances in Microbial Fuel Cells. Catalysts. 2022; 12(2):176. https://doi.org/10.3390/catal12020176
Chicago/Turabian StyleGodain, Alexiane, Naoufel Haddour, Pascal Fongarland, and Timothy M. Vogel. 2022. "Bacterial Competition for the Anode Colonization under Different External Resistances in Microbial Fuel Cells" Catalysts 12, no. 2: 176. https://doi.org/10.3390/catal12020176