
Clostridium thermocellum as a Promising Source of Genetic Material for Designer Cellulosomes: An Overview



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Reservoir of Recombinant Enzymes for In Vitro Reconstitution of Artificial Cellulosomes
2.1. Impact of Processive Endoglucanase Component in Designer Cellulosome on Biomass Decomposition
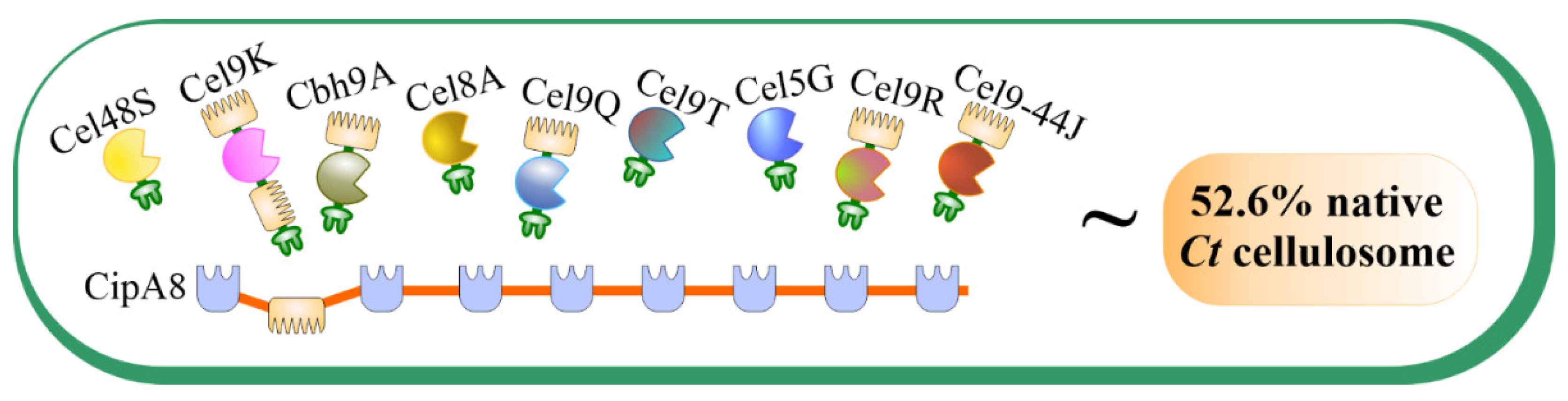
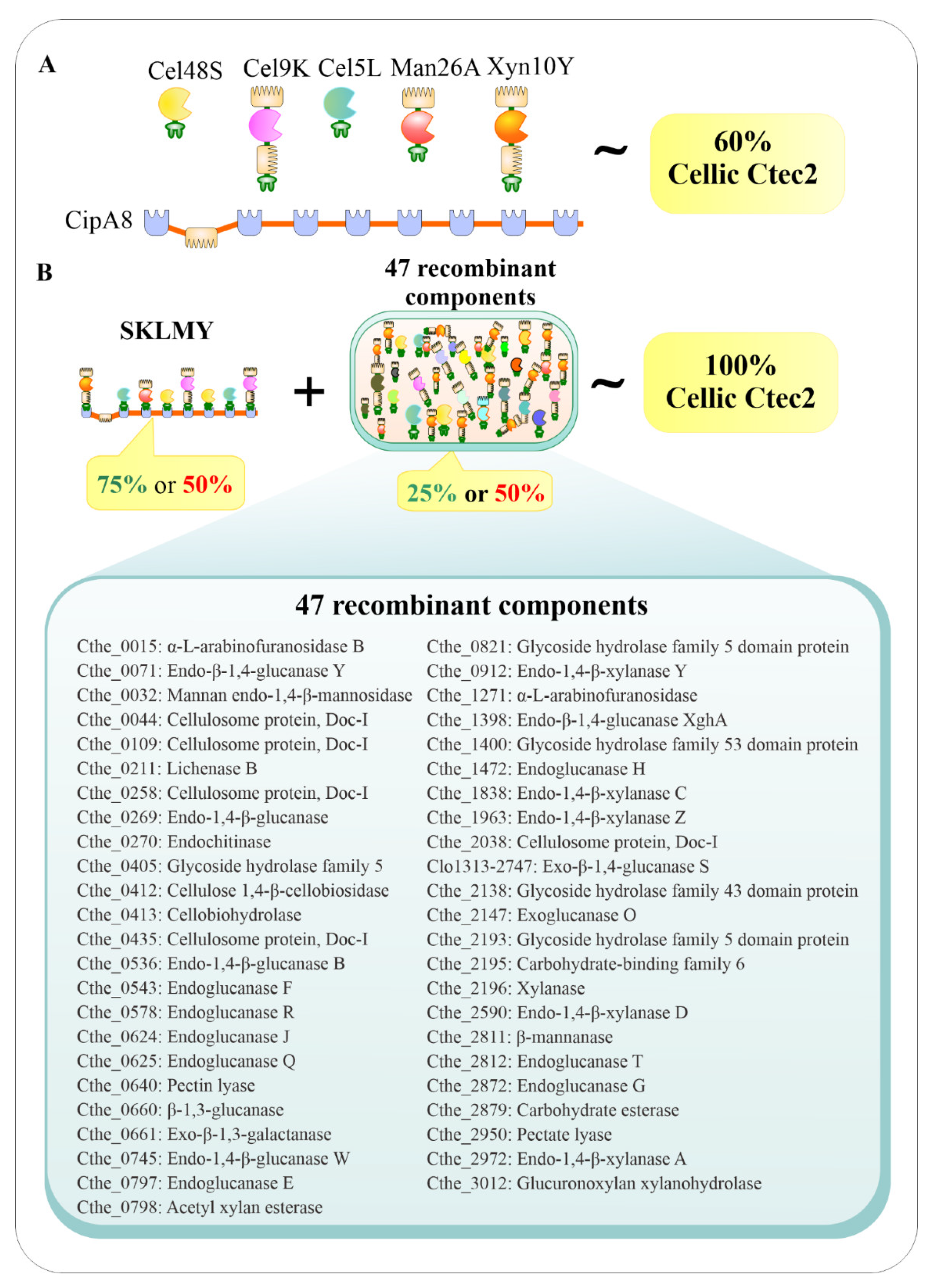
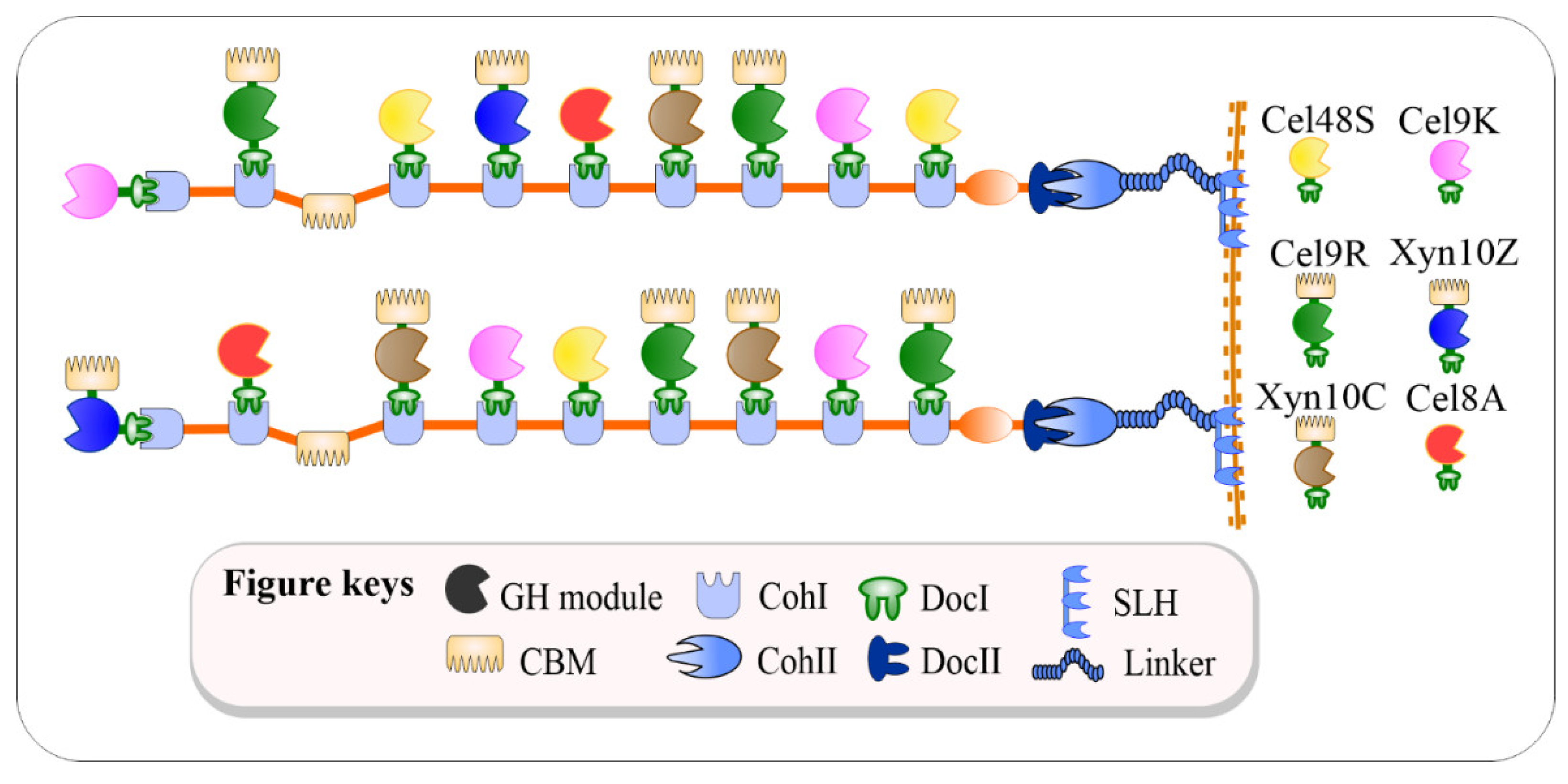
2.2. Preparation of Recombinant C. thermocellum Cellulosomal Enzyme Library
3. Incorporation of Novel Enzymes into Artificial Cellulosomes
3.1. Integration of Laccase into Designer Cellulosomes
3.2. Incorporation of Lytic Polysaccharide Monooxygenases (LPMOs) into Designer Cellulosomes
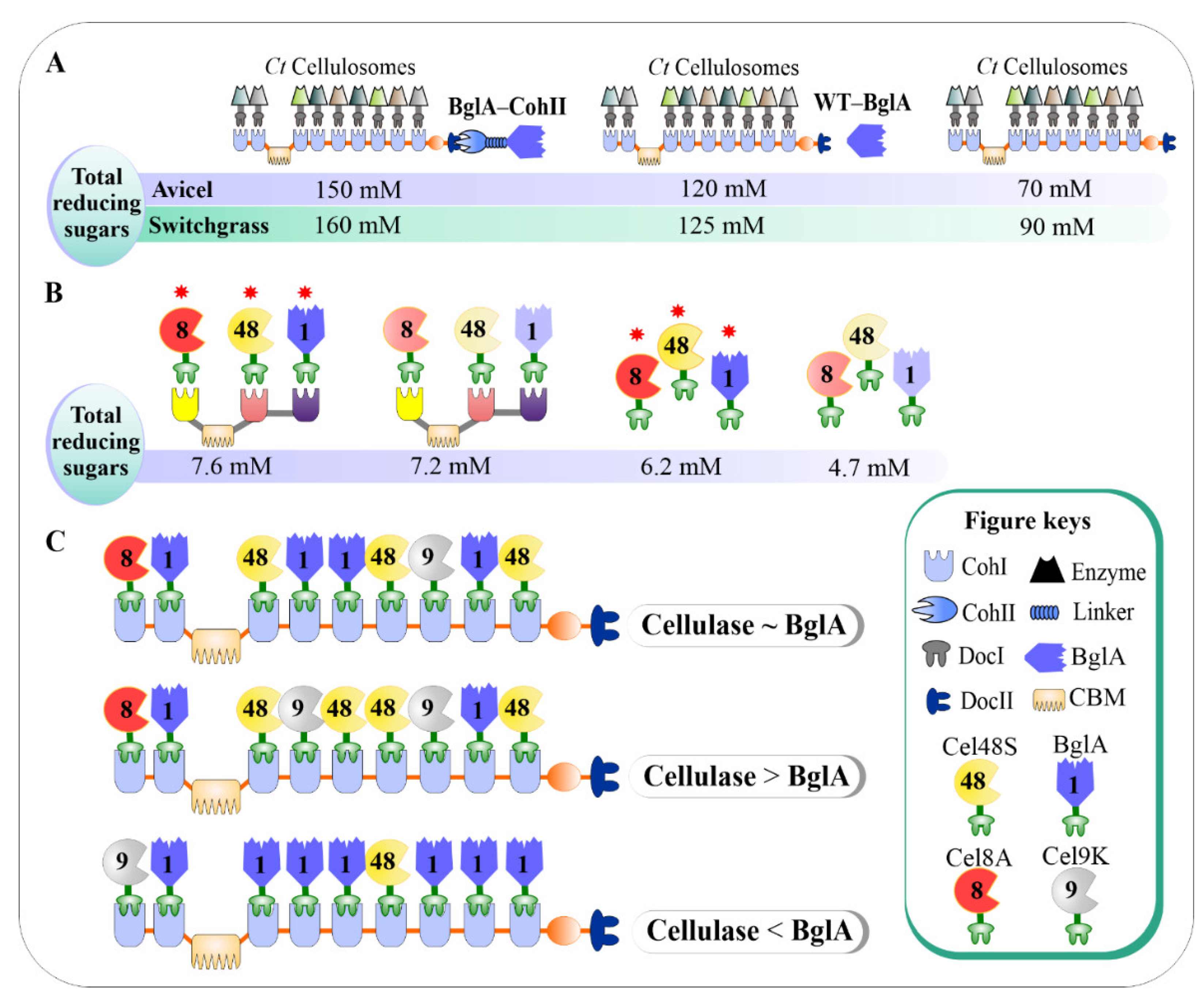
3.3. Incorporation of β-glucosidase into Designer Cellulosomes
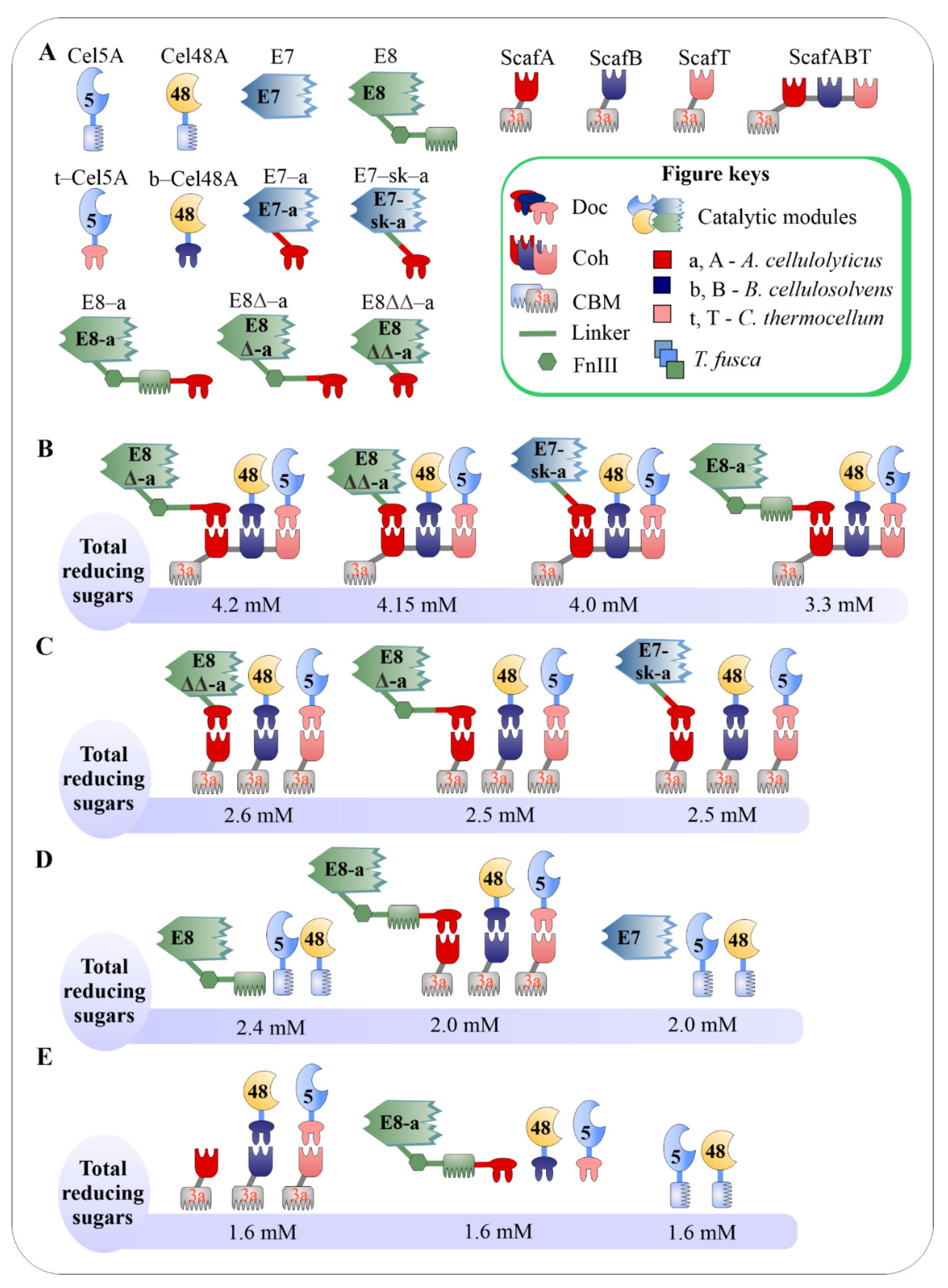
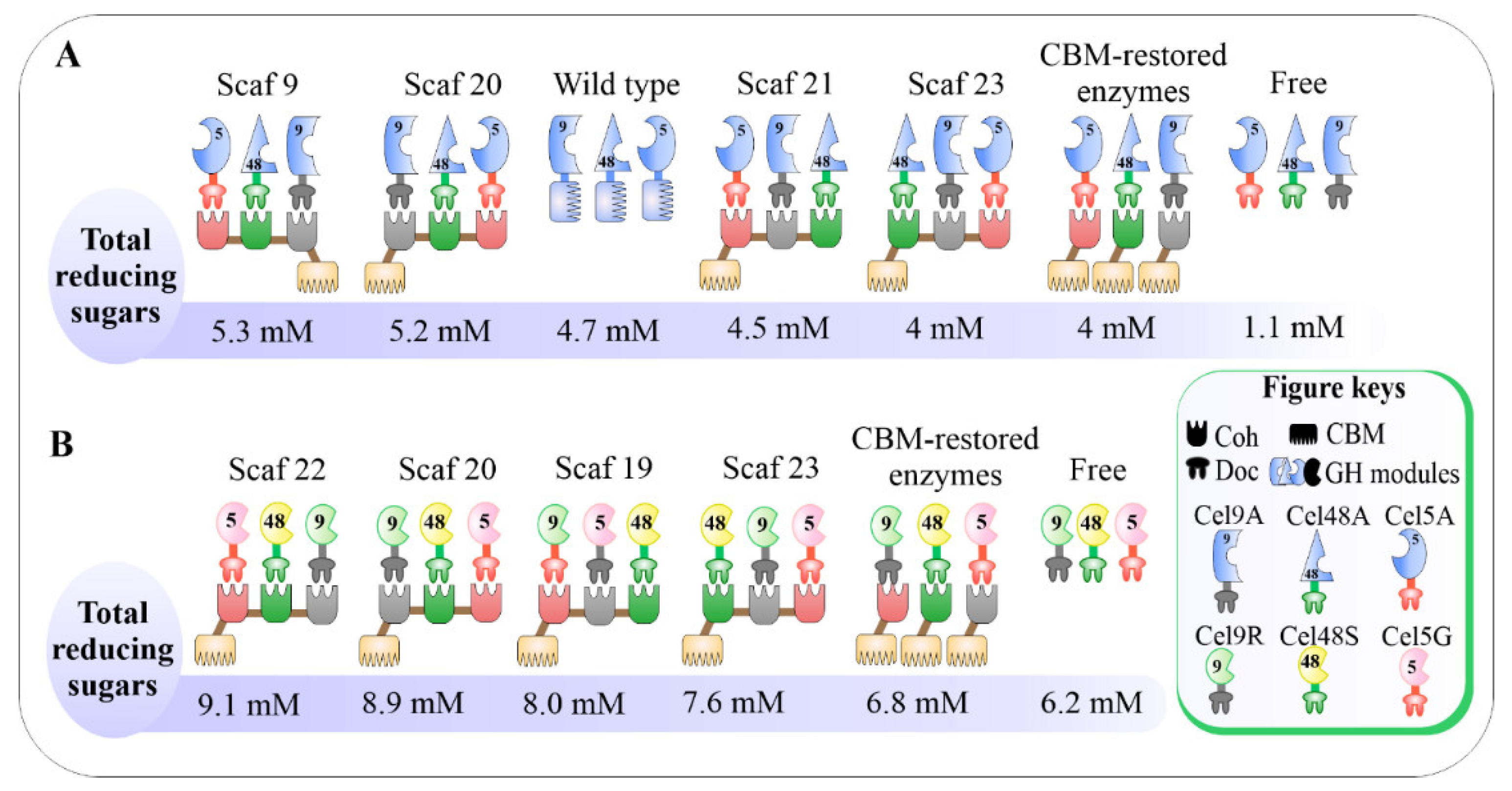
3.4. Impacts of Catalytic Enzyme Positions on Enzymatic Activity of Designer Cellulosomes
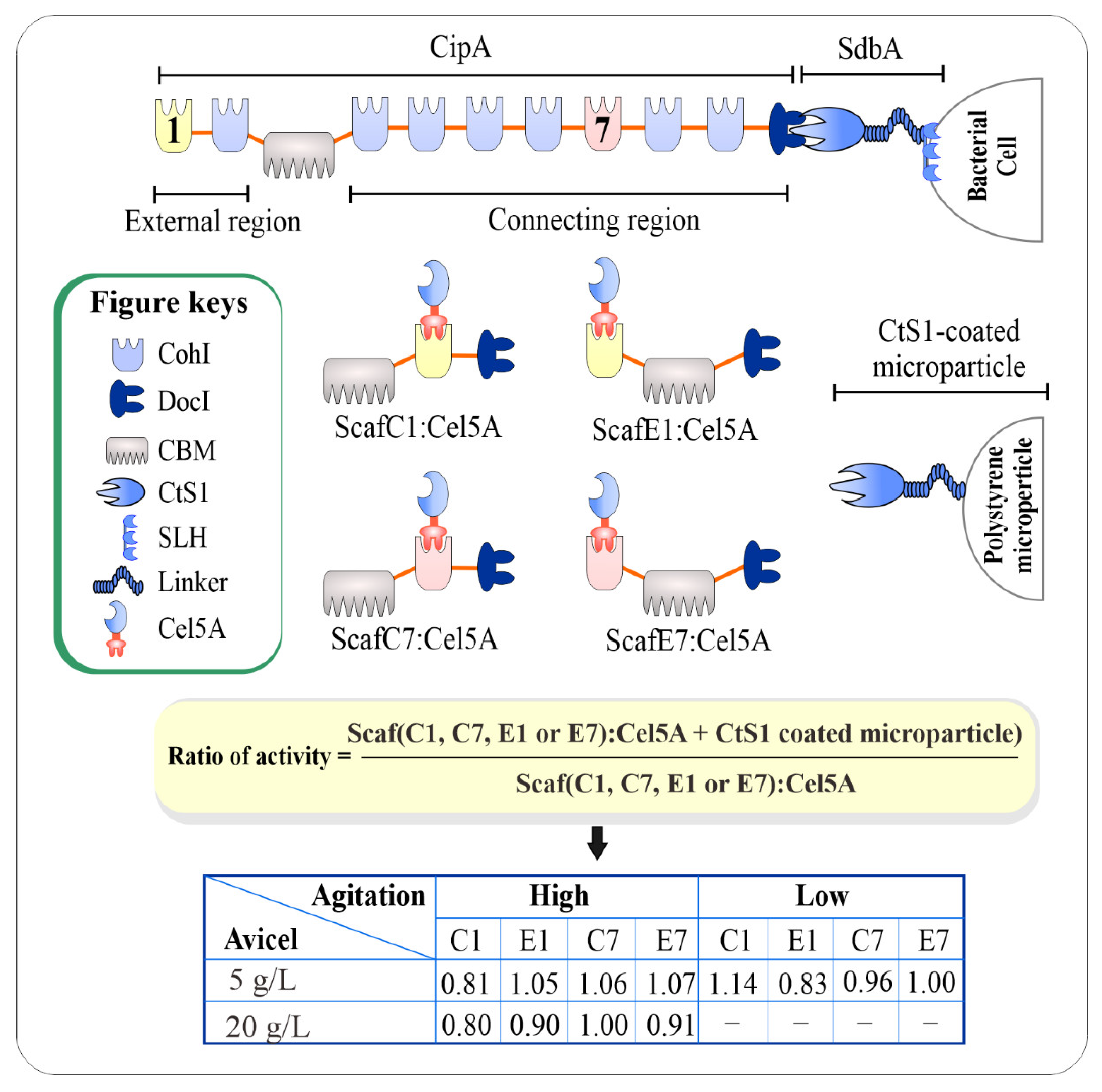
3.5. Impact of Mechanical Stability of CohI on Hydrolytic Activity of Designer Cellulosomes
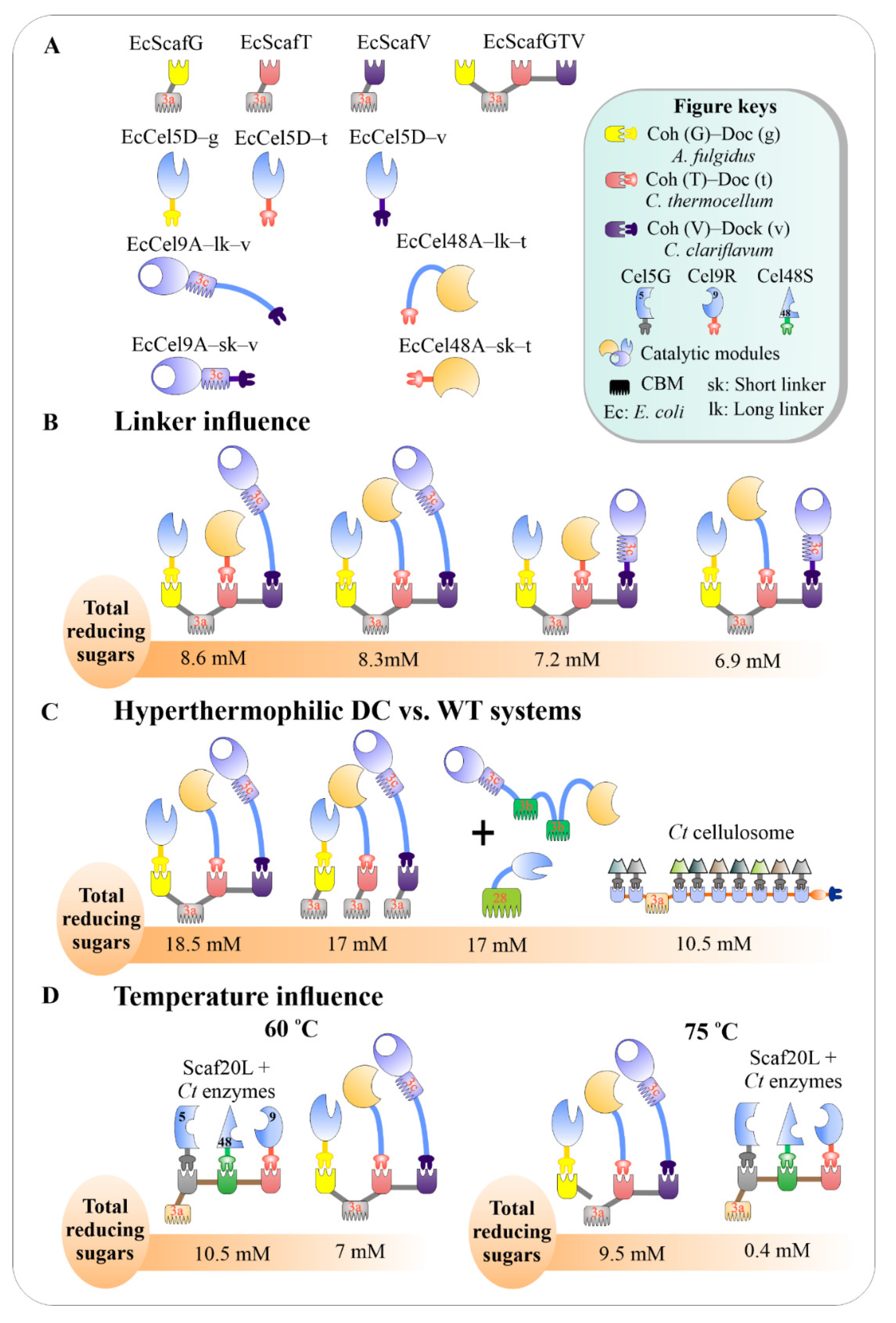
3.6. Incorporation of Hyper-Thermophilic Enzymes into Designer Cellulosomes
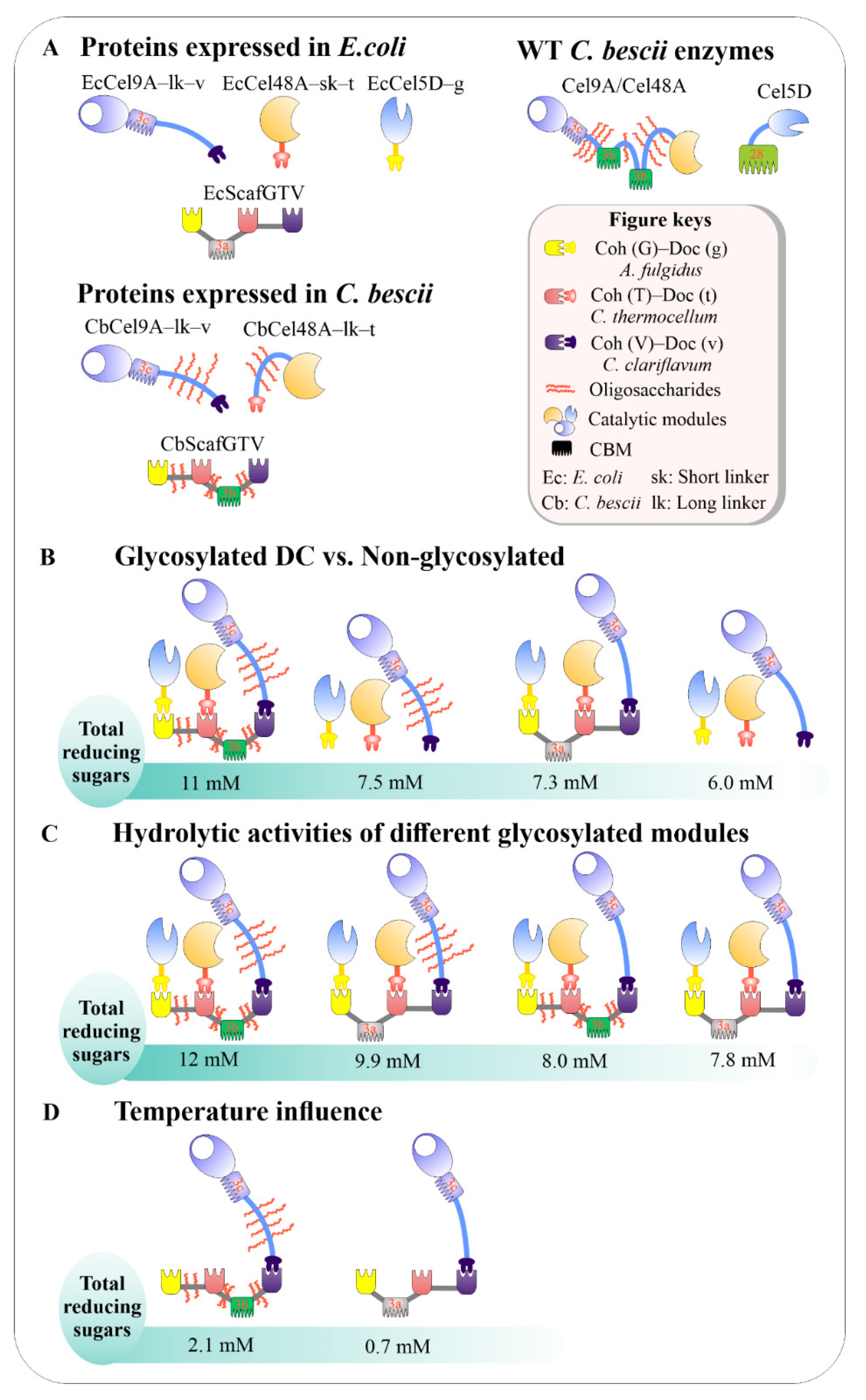
3.7. Effects of Glycosylation on Enzyme Activity
4. Conversion of Non-Cellulolytic Biofuel Microbes into Consolidated Bioprocessing Microbes
4.1. Creation of Cellulolytic Bacillus Subtilis
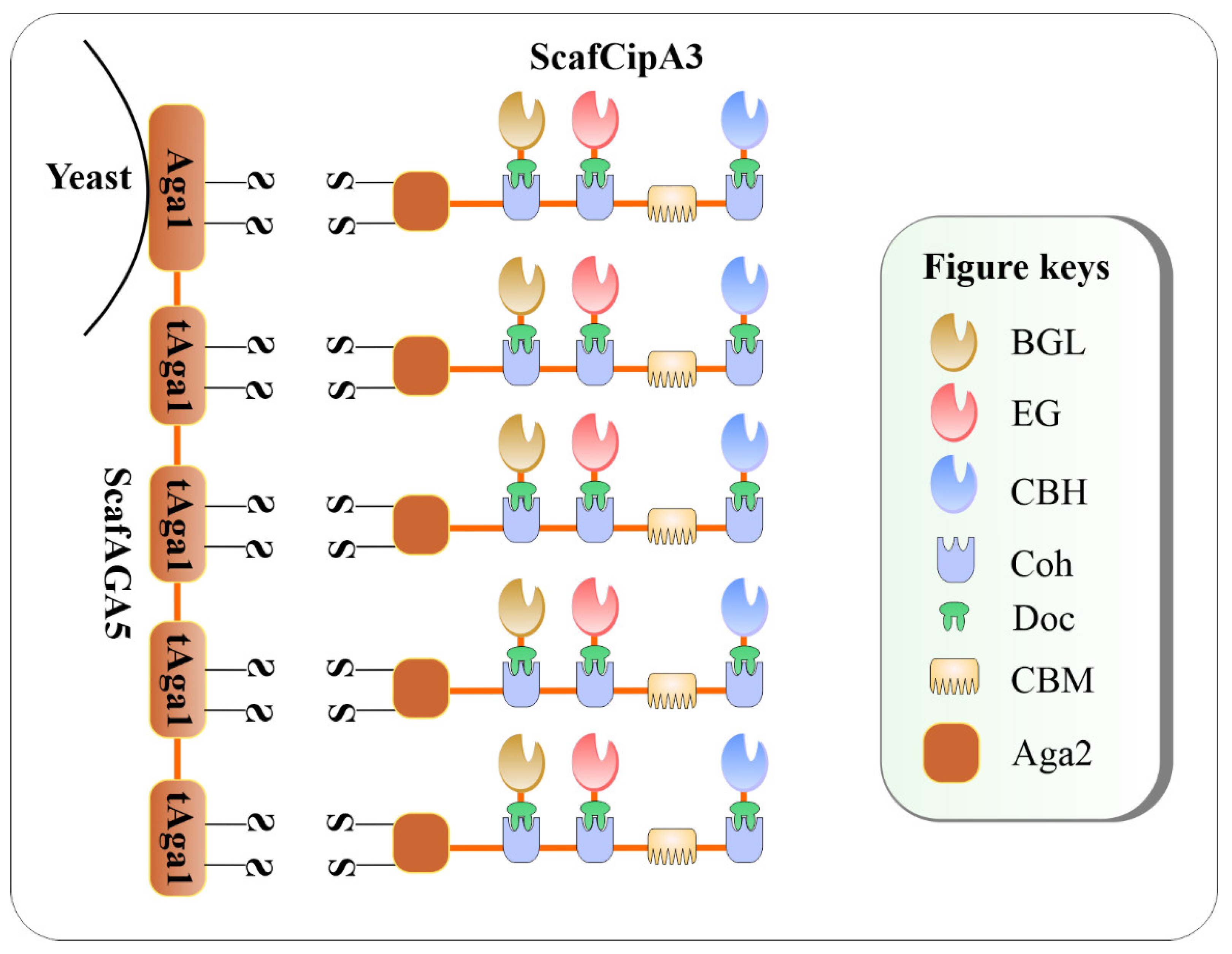
4.2. CBP-Enabling Saccharomyces cerevisiae
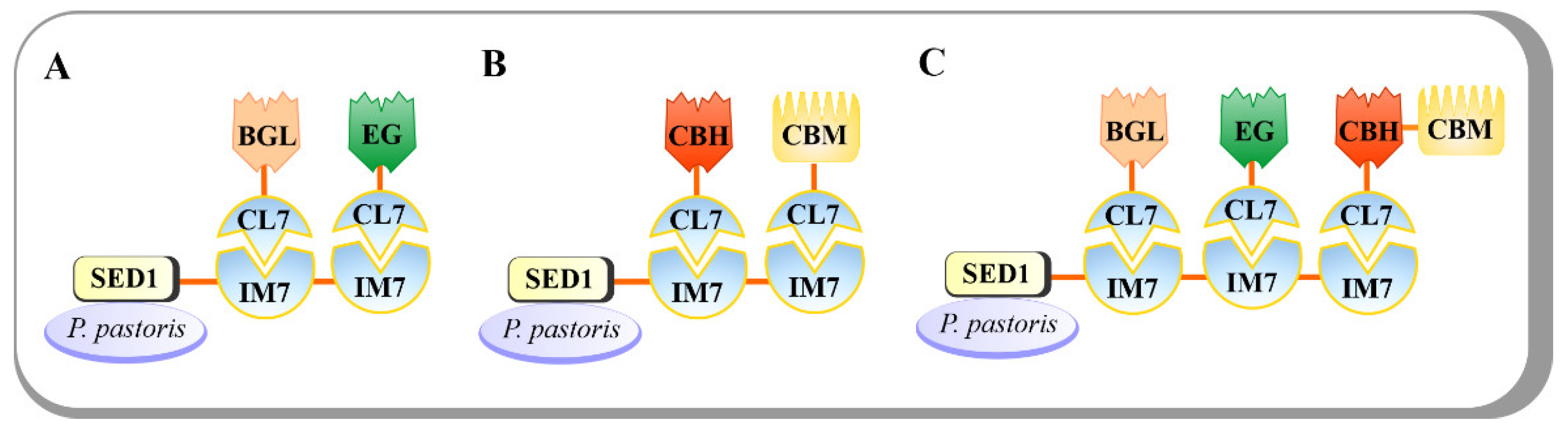
4.3. CBP-Enabling Pichia pastoris
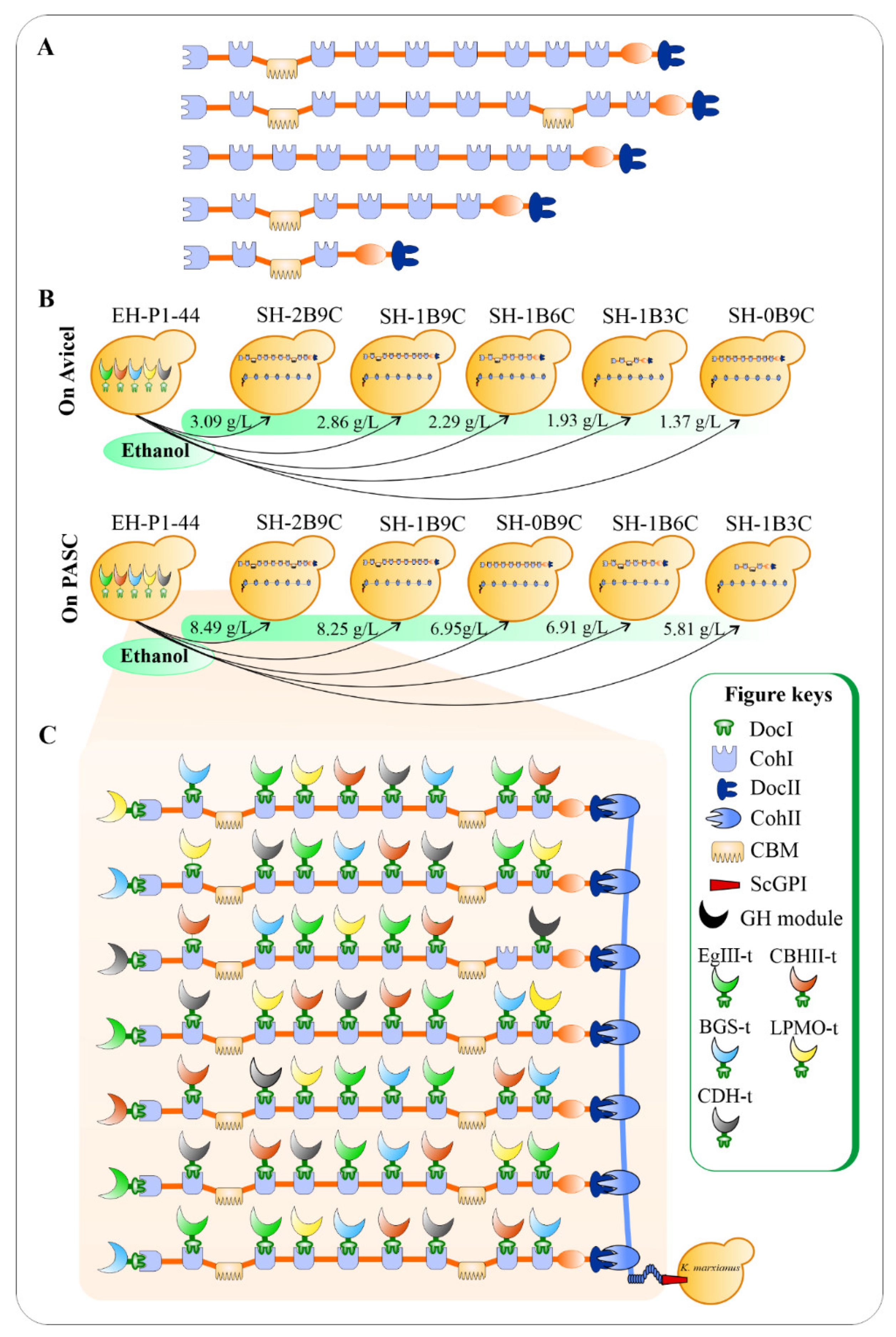
4.4. Consolidated Bioprocessing-Enabling K. marxianus
5. Promise, Limitations and Future Directions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Zhivin-Nissan, O.; Dassa, B.; Morag, E.; Kupervaser, M.; Levin, Y.; Bayer, E. Unraveling Essential Cellulosomal Components of the (Pseudo)Bacteroides Cellulosolvens Reveals an Extensive Reservoir of Novel Catalytic Enzymes. Biotechnol. Biofuels 2019, 12, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Pauly, M.; Keegstra, K. Cell-wall Carbohydrates and Their Modification as a Resource for Biofuels. Plant J. 2008, 54, 559–568. [Google Scholar] [CrossRef]
- Raman, B.; Pan, C.; Hurst, G.; Rodriguez, M., Jr.; McKeown, C.; Lankford, P.; Samatova, N.; Mielenz, J. Impact of Pretreated Switchgrass and Biomass Carbohydrates on Clostridium Thermocellum ATCC 27405 Cellulosome Composition: A Quantitative Proteomic Analysis. PLoS ONE 2009, 4, e5271. [Google Scholar] [CrossRef] [Green Version]
- Mechaly, A.; Fierobe, H.-P.; Belaich, A.; Belaich, J.-P.; Lamed, R.; Shoham, Y.; Bayer, E. Cohesin-Dockerin Interaction in Cellulosome Assembly-A Single Hydroxyl Group of a Dockerin Domain Distinguishes Between Nonrecognition and High Affinity Recognition. J. Biol. Chem. 2001, 276, 9883–9888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayer, E.; Lamed, R.; White, B.; Flint, H. From Cellulosomes to Cellulosomics. Chem. Rec. 2008, 8, 364–377. [Google Scholar] [CrossRef] [PubMed]
- Fierobe, H.-P.; Mingardon, F.; Mechaly, A.; Bélaïch, A.; Rincon, M.; Pagès, S.; Lamed, R.; Tardif, C.; Bélaïch, J.-P.; Bayer, E. Action of Designer Cellulosomes on Homogeneous Versus Complex Substrates. J. Biol. Chem. 2005, 280, 16325–16334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moraïs, S.; Morag, E.; Barak, Y.; Goldman, D.; Hadar, Y.; Lamed, R.; Shoham, Y.; Wilson, D.; Bayer, E. Deconstruction of Lignocellulose into Soluble Sugars by Native and Designer Cellulosomes. mBio 2012, 3, e00508-12. [Google Scholar] [CrossRef] [Green Version]
- Davidi, L.; Moraïs, S.; Artzi, L.; Knop, D.; Hadar, Y.; Arfi, Y.; Bayer, E. Toward Combined Delignification and Saccharification of Wheat Straw by a Laccase-Containing Designer Cellulosome. Proc. Natl. Acad. Sci. USA 2016, 113, 10854–10859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunnoo, M.; Cazade, P.-A.; Galera-Prat, A.; Nash, M.; Czjzek, M.; Cieplak, M.; Alvarez, B.; Aguilar, M.; Karpol, A.; Gaub, H.; et al. Nanoscale Engineering of Designer Cellulosomes. Adv. Mater. 2016, 28, 5619–5647. [Google Scholar] [CrossRef] [PubMed]
- Leis, B.; Held, C.; AndreeBen, B.; Liebl, W.; Graubner, S.; Schulte, L.-P.; Schwarz, W.; Zverlov, V. Optimizing the Composition of a Synthetic Cellulosome Complex for the Hydrolysis of Softwood Pulp: Identification of the Enzymatic Core Functions and Biochemical Complex Characterization. Biotechnol. Biofuels 2018, 11, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Fan, L.; Zhang, Z.; Yu, X.; Xue, Y.; Tan, T. Self-Surface Assembly of Cellulosomes with Two Miniscaffoldins on Saccharomyces Cerevisiae for Cellulosic Ethanol Production. Proc. Natl. Acad. Sci. USA 2012, 109, 13260–13265. [Google Scholar] [CrossRef] [Green Version]
- Nataf, Y.; Bahari, L.; Kahel-Raifer, H.; Borovok, I.; Lamed, R.; Bayer, E.; Sonenshein, A.; Shoham, Y. Clostridium Thermocellum Cellulosomal Genes Are Regulated by Extracytoplasmic Polysaccharides via Alternative Sigma Factor. Proc. Natl. Acad. Sci. USA 2010, 107, 18646–18651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leis, B.; Held, C.; Bergkemper, F.; Dennemarck, K.; Steinbauer, R.; Reiter, A.; Mechelke, M.; Moerch, M.; Graubner, S.; Liebl, W.; et al. Comparative Characterization of All Cellulosomal Cellulases from Clostridium Thermocellum Reveals High Diversity in Endoglucanase Product Formation Essential for Complex Activity. Biotechnol. Biofuels 2017, 10, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lytle, B.; Myers, C.; Kruus, K.; Wu, J.-H. Interactions of the CelS Binding Ligand with Various Receptor Domains of the Clostridium Thermocellum Cellulosomal Scaffolding Protein, CipA. J. Bacteriol. 1996, 178, 1200–1203. [Google Scholar] [CrossRef] [Green Version]
- Pagès, S.; Bélaïch, A.; Bélaïch, J.-P.; Morag, E.; Lamed, R.; Shoham, Y.; Bayer, E. Species-specificity of the Cohesin-dockerin Interaction between Clostridium Thermocellum and Clostridium Cellulolyticum: Prediction of Specificity Determinants of the Dockerin Domain. Proteins 1997, 29, 517–527. [Google Scholar] [CrossRef]
- Moraïs, S.; Barak, Y.; Caspi, J.; Hadar, Y.; Lamed, R.; Shoham, Y.; Wilson, D.; Bayer, E. Cellulase-Xylanase Synergy in Designer Cellulosomes for Enhanced Degradation of a Complex Cellulosic Substrate. mBio 2010, 1, e00285-10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahn, A.; Moraïs, S.; Galanopoulou, A.; Chung, D.; Sarai, N.; Hengge, N.; Hatzinikolaou, D.; Himmel, M.; Bomble, Y.; Bayer, E. Creation of a Functional Hyperthermostable Designer Cellulosome. Biotechnol. Biofuels 2019, 12, 1–15. [Google Scholar] [CrossRef]
- Gold, N.; Martin, V.J. Global View of the Clostridium Thermocellum Cellulosome Revealed by Quantitative Proteomic Analysis. J. Bacteriol. 2007, 189, 6787–6795. [Google Scholar] [CrossRef] [Green Version]
- Dykstra, A.; Brice, L.; Rodriguez Jr, M.; Raman, B.; Izquierdo, J.; Cook, K.; Lynd, L.; Hettich, R. Development of a Multipoint Quantitation Method to Simultaneously Measure Enzymatic and Structural Components of the Clostridium Thermocellum Cellulosome Protein Complex. J. Proteome Res. 2014, 13, 692–701. [Google Scholar] [CrossRef]
- Olson, D.; Tripathi, S.; Giannone, R.; Lo, J.; Caiazza, N.; Hogsett, D.; Hettich, R.; Guss, A.; Dubrovsky, G.; Lynd, L. Deletion of the Cel48S Cellulase from Clostridium Thermocellum. Proc. Natl. Acad. Sci. USA 2010, 107, 17727–17732. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.-J.; Liu, S.; Dong, S.; Li, R.; Feng, Y.; Cui, Q. Determination of the Native Features of the Exoglucanase Cel48S from Clostridium Thermocellum. Biotechnol. Biofuels 2018, 11, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.; Anandharaj, M.; Ho, C.; Tsuge, K.; Tsai, T.; Ke, H.; Lin, Y.; Ha-Tran, D.; Li, W.; Huang, C. Biomimetic Strategy for Constructing Clostridium Thermocellum Cellulosomal Operons in Bacillus Subtilis. Biotechnol. Biofuels 2018, 11, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Hirano, K.; Saito, T.; Shinoda, S.; Haruki, M.; Hirano, N. In Vitro Assembly and Cellulolytic Activity of a β-Glucosidase-Integrated Cellulosome Complex. FEMS Microbiol. Lett. 2019, 366, fnz209. [Google Scholar] [CrossRef] [PubMed]
- Moraïs, S.; Stern, J.; Kahn, A.; Galanopoulou, A.; Yoav, S.; Shamshoum, M.; Smith, M.; Hatzinikolaou, D.; Arnold, F.; Bayer, E. Enhancement of Cellulosome-Mediated Deconstruction of Cellulose by Improving Enzyme Thermostability. Biotechnol. Biofuels 2016, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, H.-T.; Gao, Y.; Yang, Y.-M.; Xiao, W.-J.; Liu, S.-H.; Xia, W.-C.; Liu, Z.-L.; Yi, L.; Jiang, Z.-B. Synergistic Effect of Cellulase and Xylanase during Hydrolysis of Natural Lignocellulosic Substrates. Bioresour. Technol. 2016, 219, 710–715. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, P.; Mahajan, R. Cellulase and Xylanase Synergism in Industrial Biotechnology. Appl. Microbiol. Biotechnol. 2019, 103, 8711–8724. [Google Scholar] [CrossRef]
- Huang, C.; Zhao, C.; Li, H.; Xiong, L.; Chen, X.; Luo, M.; Chen, X. Comparison of Different Pretreatments on the Synergistic Effect of Cellulase and Xylanase during the Enzymatic Hydrolysis of Sugarcane Bagasse. RSC Adv. 2018, 8, 30725–30731. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves, G.; Takasugi, Y.; Jia, L.; Mori, Y.; Noda, S.; Tanaka, T.; Ichinose, H.; Kamiya, N. Synergistic Effect and Application of Xylanases as Accessory Enzymes to Enhance the Hydrolysis of Pretreated Bagasse. Enzyme Microb. Technol. 2015, 72, 16–24. [Google Scholar] [CrossRef] [Green Version]
- Moraïs, S.; Barak, Y.; Caspi, J.; Hadar, Y.; Lamed, R.; Shoham, Y.; Wilson, D.; Bayer, E. Contribution of a Xylan-Binding Module to the Degradation of a Complex Cellulosic Substrate by Designer Cellulosomes. Appl. Environ. Microbiol. 2010, 76, 3787–3796. [Google Scholar] [CrossRef] [Green Version]
- del Pulgar, E.; Saadeddin, A. The Cellulolytic System of Thermobifida Fusca. Crit. Rev. Microbiol. 2014, 40, 236–247. [Google Scholar] [CrossRef]
- Stern, J.; Kahn, A.; Vazana, Y.; Shamshoum, M.; Moraïs, S.; Lamed, R.; Bayer, E. Significance of Relative Position of Cellulases in Designer Cellulosomes for Optimized Cellulolysis. PLoS ONE 2015, 10, e0127326. [Google Scholar] [CrossRef]
- Stern, J.; Moraïs, S.; Lamed, R.; Bayer, E. Adaptor Scaffoldins: An Original Strategy for Extended Designer Cellulosomes, Inspired from Nature. mBio 2016, 7, e00083-16. [Google Scholar] [CrossRef] [Green Version]
- Arfi, Y.; Shamshoum, M.; Rogachev, I.; Peleg, Y.; Bayer, E. Integration of Bacterial Lytic Polysaccharide Monooxygenases into Designer Cellulosomes Promotes Enhanced Cellulose Degradation. Proc. Natl. Acad. Sci. USA 2014, 111, 9109–9114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.-Y.; Lee, C.-C.; Chen, H.-S.; Yang, C.-H.; Wang, S.-P.; Wu, J.-H.; Meng, M. Modification of Lignin in Sugarcane Bagasse by a Monocopper Hydrogen Peroxide-Generating Oxidase from Thermobifida Fusca. Process Biochem. 2016, 51, 1486–1495. [Google Scholar] [CrossRef]
- Gaber, Y.; Rashad, B.; Hussein, R.; Abdelgawad, M.; Ali, N.; Dishisha, T.; Várnai, A. Heterologous Expression of Lytic Polysaccharide Monooxygenases (LPMOs). Biotechnol. Adv. 2020, 43, 107583. [Google Scholar] [CrossRef]
- Brenelli, L.; Squina, F.; Felby, C.; Cannella, D. Laccase-Derived Lignin Compounds Boost Cellulose Oxidative Enzymes AA9. Biotechnol. Biofuels 2018, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Westereng, B.; Cannella, D.; Agger, J.; Jørgensen, H.; Andersen, M.; Eijsink, V.; Felby, C. Enzymatic Cellulose Oxidation Is Linked to Lignin by Long-Range Electron Transfer. Sci. Rep. 2015, 5, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Wang, B.; Xiao, Q.; Wu, S. A Kinetics Modeling Study on the Inhibition of Glucose on Cellulosome of Clostridium Thermocellum. Bioresour. Technol. 2015, 190, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Lamed, R.; Kenig, R.; Morag, E.; Calzada, J.; De Micheo, F.; Bayer, E. Efficient Cellulose Solubilization by a Combined Cellulosome Beta-Glucosidase System. Appl. Biochem. Biotechnol. 1991, 27, 173–183. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, S.; Li, R.; Hong, W.; Xiao, Y.; Feng, Y.; Cui, Q.; Liu, Y.-J. Efficient Whole-Cell-Catalyzing Cellulose Saccharification Using Engineered Clostridium Thermocellum. Biotechnol. Biofuels 2017, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Yoav, S.; Stern, J.; Salama-Alber, O.; Frolow, F.; Anbar, M.; Karpol, A.; Hadar, Y.; Morag, E.; Bayer, E. Directed Evolution of Clostridium Thermocellum β-Glucosidase A Towards Enhanced Thermostability. Int. J. Mol. Sci. 2019, 20, 4701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinoda, S.; Kurosaki, M.; Kokuzawa, T.; Hirano, K.; Takano, H.; Ueda, K.; Haruki, M.; Hirano, N. Comparative Biochemical Analysis of Cellulosomes Isolated from Clostridium Clariflavum DSM 19732 and Clostridium Thermocellum ATCC 27405 Grown on Plant Biomass. Appl. Biochem. Biotechnol. 2019, 187, 994–1010. [Google Scholar] [CrossRef] [PubMed]
- Gefen, G.; Anbar, M.; Morag, E.; Lamed, R.; Bayer, E. Enhanced Cellulose Degradation by Targeted Integration of a Cohesin-Fused β-Glucosidase into the Clostridium Thermocellum Cellulosome. Proc. Natl. Acad. Sci. USA 2012, 109, 10298–10303. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Xiao, Y.; Feng, Y.; Li, B.; Li, W.; Cui, Q. The Spatial Proximity Effect of Beta-Glucosidase and Cellulosomes on Cellulose Degradation. Enzyme Microb. Technol. 2018, 115, 52–61. [Google Scholar] [CrossRef]
- Waeonukul, R.; Kosugi, A.; Tachaapaikoon, C.; Pason, P.; Ratanakhanokchai, K.; Prawitwong, P.; Deng, L.; Saito, M.; Mori, Y. Efficient Saccharification of Ammonia Soaked Rice Straw by Combination of Clostridium Thermocellum Cellulosome and Thermoanaerobacter Brockii β-Glucosidase. Bioresour. Technol. 2012, 107, 352–357. [Google Scholar] [CrossRef]
- Tang, H.; Wang, J.; Wang, S.; Shen, Y.; Petranovic, D.; Hou, J.; Bao, X. Efficient Yeast Surface-Display of Novel Complex Synthetic Cellulosomes. Microb. Cell Factories 2018, 17, 1–13. [Google Scholar] [CrossRef]
- Qi, K.; Chen, C.; Yan, F.; Feng, Y.; Bayer, E.; Kosugi, A.; Cui, Q.; Liu, Y.-J. Coordinated β-Glucosidase Activity with the Cellulosome Is Effective for Enhanced Lignocellulose Saccharification. Bioresour. Technol. 2021, 337, 125441. [Google Scholar] [CrossRef]
- Wu, B.; Zheng, S.; Pedroso, M.; Guddat, L.; Chang, S.; He, B.; Schenk, G. Processivity and Enzymatic Mechanism of a Multifunctional Family 5 Endoglucanase from Bacillus Subtilis BS-5 with Potential Applications in the Saccharification of Cellulosic Substrates. Biotechnol. Biofuels 2018, 11, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, A.; Dias, F.; Nagy, T.; Prates, J.; Proctor, M.; Smith, N.; Bayer, E.; Davies, G.; Ferreira, L.; Romão, M.; et al. Evidence for a Dual Binding Mode of Dockerin Modules to Cohesins. Proc. Natl. Acad. Sci. USA 2007, 104, 3089–3094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valbuena, A.; Oroz, J.; Hervás, R.; Vera, A.; Rodríguez, D.; Menéndez, M.; Sulkowska, J.; Cieplak, M.; Carrión-Vázquez, M. On the Remarkable Mechanostability of Scaffoldins and the Mechanical Clamp Motif. Proc. Natl. Acad. Sci. USA 2009, 106, 13791–13796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galera-Prat, A.; Moraïs, S.; Vazana, Y.; Bayer, E.; Carrión-Vázquez, M. The Cohesin Module Is a Major Determinant of Cellulosome Mechanical Stability. J. Biol. Chem. 2018, 293, 7139–7147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galera-Prat, A.; Vera, A.; Moraïs, S.; Vazana, Y.; Bayer, E.; Carrión-Vázquez, M. Impact of Scaffoldin Mechanostability on Cellulosomal Activity. Biomater. Sci. 2020, 8, 3601–3610. [Google Scholar] [CrossRef] [PubMed]
- Hong, W.; Zhang, J.; Feng, Y.; Mohr, G.; Lambowitz, A.; Cui, G.-Z.; Liu, Y.-J.; Cui, Q. The Contribution of Cellulosomal Scaffoldins to Cellulose Hydrolysis by Clostridium Thermocellum Analyzed by Using Thermotargetrons. Biotechnol. Biofuels 2014, 7, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vazana, Y.; Moraïs, S.; Barak, Y.; Lamed, R.; Bayer, E. Chapter 23: Designer Cellulosomes For Enhanced Hydrolysis of Cellulosic Substrates. In Cellulases; Gilbert, H., Ed.; Method in Enzymology; Elsevier Inc.: Amsterdam, The Netherlands, 2012; Volume 510, ISBN 978-0-12-415931-0. [Google Scholar]
- Basen, M.; Rhaesa, A.; Kataeva, I.; Prybol, C.; Scott, I.; Poole, F.; Adams, M. Degradation of High Loads of Crystalline Cellulose and of Unpretreated Plant Biomass by the Thermophilic Bacterium Caldicellulosiruptor Bescii. Bioresour. Technol. 2014, 152, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Kataeva, I.; Foston, M.; Yang, S.-J.; Pattathil, S.; Biswal, A.; Pool, F., II; Basen, M.; Rhaesa, A.; Thomas, T.; Azadi, P.; et al. Carbohydrate and Lignin Are Simultaneously Solubilized from Unpretreated Switchgrass by Microbial Action at High Temperature. Energy Environ. Sci. 2013, 6, 2186–2195. [Google Scholar] [CrossRef]
- Young, J.; Chung, D.; Bomble, Y.; Himmel, M.; Westpheling, J. Deletion of Caldicellulosiruptor Bescii CelA Reveals Its Crucial Role in the Deconstruction of Lignocellulosic Biomass. Biotechnol. Biofuels 2014, 7, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Brunecky, R.; Chung, D.; Sarai, N.; Hengge, N.; Russell, J.; Young, J.; Mittal, A.; Pason, P.; Wall, T.; Michener, W.; et al. High Activity CAZyme Cassette for Improving Biomass Degradation in Thermophiles. Biotechnol. Biofuels 2018, 11, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caspi, J.; Barak, Y.; Haimovitz, R.; Irwin, D.; Lamed, R.; Wilson, D.; Bayer, E. Effect of Linker Length and Dockerin Position on Conversion of a Thermobifida Fusca Endoglucanase to the Cellulosomal Mode. Appl. Environ. Microbiol. 2009, 75, 7335–7342. [Google Scholar] [CrossRef] [Green Version]
- Vazana, Y.; Moraïs, S.; Barak, Y.; Lamed, R.; Bayer, E. Interplay between Clostridium Thermocellum Family 48 and Family 9 Cellulases in Cellulosomal versus Noncellulosomal States. Appl. Environ. Microbiol. 2010, 76, 3236–3243. [Google Scholar] [CrossRef] [Green Version]
- Kahn, A.; Moraïs, S.; Chung, D.; Sarai, N.; Hengge, N.; Kahn, A.; Himmel, M.; Bayer, E.; Bomble, Y. Glycosylation of Hyperthermostable Designer Cellulosome Components Yields Enhanced Stability and Cellulose Hydrolysis. FEBS J. 2020, 287, 4370–4388. [Google Scholar] [CrossRef]
- Lambertz, C.; Garvey, M.; Klinger, J.; Heesel, D.; Klose, H.; Fischer, R.; Commandeur, U. Challenges and Advances in the Heterologous Expression of Cellulolytic Enzymes: A Review. Biotechnol. Biofuels 2014, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Cui, W.; Han, L.; Suo, F.; Liu, Z.; Zhou, L.; Zhou, Z. Exploitation of Bacillus Subtilis as a Robust Workhorse for Production of Heterologous Proteins and Beyond. World J. Microbiol. Biotechnol. 2018, 34, 1–19. [Google Scholar] [CrossRef]
- Nishizaki, T.; Tsuge, K.; Itaya, M.; Doi, N.; Yanagawa, H. Metabolic Engineering of Carotenoid Biosynthesis in Escherichia Coli by Ordered Gene Assembly in Bacillus Subtilis. Appl. Environ. Microbiol. 2007, 73, 1355–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuge, K.; Matsui, K.; Itaya, M. One Step Assembly of Multiple DNA Fragments with a Designed Order and Orientation in Bacillus Subtilis Plasmid. Nucleic Acids Res. 2003, 31, e133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Y.; Ang, E.; Zhao, H. Engineered Pentafunctional Minicellulosome for Simultaneous Saccharification and Ethanol Fermentation in Saccharomyces Cerevisiae. Appl. Environ. Microbiol. 2014, 80, 6677–6684. [Google Scholar] [CrossRef] [Green Version]
- Karbalaei, M.; Rezaee, S.; Farsiani, H. Pichia Pastoris: A Highly Successful Expression System for Optimal Synthesis of Heterologous Proteins. J. Cell. Physiol. 2020, 235, 5867–5881. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Wang, X.; Sun, W.; Chen, L.; Li, S.; Wu, K.; Ma, L.; Liu, Y. Engineering Pichia Pastoris with Surface-Display Minicellulosomes for Carboxymethyl Cellulose Hydrolysis and Ethanol Production. Biotechnol. Biofuels 2020, 13, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Vassylyeva, M.; Klyuyev, S.; Vassylyev, A.; Wesson, H.; Zhang, Z.; Renfrow, M.; Wang, H.; Higgins, N.; Chow, L.; Vassylyev, D. Efficient, Ultra-High-Affinity Chromatography in a One-Step Purification of Complex Proteins. Proc. Natl. Acad. Sci. USA 2017, 114, 5138–5147. [Google Scholar] [CrossRef] [Green Version]
- Hirano, K.; Nihei, S.; Hasegawa, H.; Haruki, M.; Hirano, N. Stoichiometric Assembly of the Cellulosome Generates Maximum Synergy for the Degradation of Crystalline Cellulose, as Revealed by In Vitro Reconstitution of the Clostridium Thermocellum Cellulosome. Appl. Environ. Microbiol. 2015, 81, 4756–4766. [Google Scholar] [CrossRef] [Green Version]
- Ha-Tran, D.; Lai, R.-Y.; Nguyen, T.; Huang, E.; Lo, S.-C.; Huang, C. Construction of Engineered RuBisCO Kluyveromyces Marxianus for a Dual Microbial Bioethanol Production System. PLoS ONE 2021, 16, e0247135. [Google Scholar] [CrossRef]
- Andreu, C.; del Olmo, M. Whole-Cell Biocatalysis in Seawater: New Halotolerant Yeast Strains for the Regio- and Stereoselectivity Reduction of 1-Phenylpropane-1,2-Dione in Saline-Rich Media. ChemBioChem 2020, 21, 1621–1628. [Google Scholar] [CrossRef] [PubMed]
- Anandharaj, M.; Lin, Y.-J.; Rani, R.; Nadendla, E.; Ho, M.-C.; Huang, C.; Cheng, J.-F.; Chang, J.-J.; Li, W.-H. Constructing a Yeast to Express the Largest Cellulosome Complex on the Cell Surface. Proc. Natl. Acad. Sci. USA 2020, 117, 2385–2394. [Google Scholar] [CrossRef] [PubMed]
- Olson, D.; Giannone, R.; Hettich, R.; Lynd, L. Role of the CipA Scaffoldin Protein in Cellulose Solubilization, as Determined by Targeted Gene Deletion and Complementation in Clostridium Thermocellum. J. Bacteriol. 2013, 195, 733–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ha-Tran, D.M.; Nguyen, T.T.M.; Huang, C.-C. Clostridium thermocellum as a Promising Source of Genetic Material for Designer Cellulosomes: An Overview. Catalysts 2021, 11, 996. https://doi.org/10.3390/catal11080996
Ha-Tran DM, Nguyen TTM, Huang C-C. Clostridium thermocellum as a Promising Source of Genetic Material for Designer Cellulosomes: An Overview. Catalysts. 2021; 11(8):996. https://doi.org/10.3390/catal11080996
Chicago/Turabian StyleHa-Tran, Dung Minh, Trinh Thi My Nguyen, and Chieh-Chen Huang. 2021. "Clostridium thermocellum as a Promising Source of Genetic Material for Designer Cellulosomes: An Overview" Catalysts 11, no. 8: 996. https://doi.org/10.3390/catal11080996