1. Introduction
Smoking processes have been used by humanity for thousands of years to preserve perishable foods, such as meat and fish [
1]. Besides the decrease in water activity, the empirically found preserving effect is caused by the condensation of numerous antimicrobial products of the pyrolysis of lignocellulose (for example, formaldehyde, methanol, phenols, and cresols) on the food surface [
2]. Formaldehyde and cresols, together with polycyclic aromatic hydrocarbons and heterocyclic amines, form a group of potent cancerogens, which in trace concentrations may contaminate the smoked goods [
3,
4]. To mitigate the apparent health risks, fractions of liquid smoke absorbates have been introduced into the market more recently [
5,
6]. These show a lesser toxic load, but leave highly toxic fractions, tar and ashes, which require laborious disposal.
Due to modern preservation techniques, such as cold chains, inert gas, and artificial preservatives, the antibiotic effects of smoke are no longer a central purpose. More of interest are the smoky and savory flavor attributes of smoked foods [
7]. Consequently, specific enzymatic processes for generating a natural smoke flavor in mild reaction conditions without the risk of formation of unwanted and toxic by-products should be developed.
Previous works have shown that wood-decaying basidiomycetes, adapted to survive on lignified plant materials, secrete a number of potent enzymes, often distinguished by unique catalytic properties when it comes to the degradation of highly cross-linked and recalcitrant lignocelluloses [
8,
9,
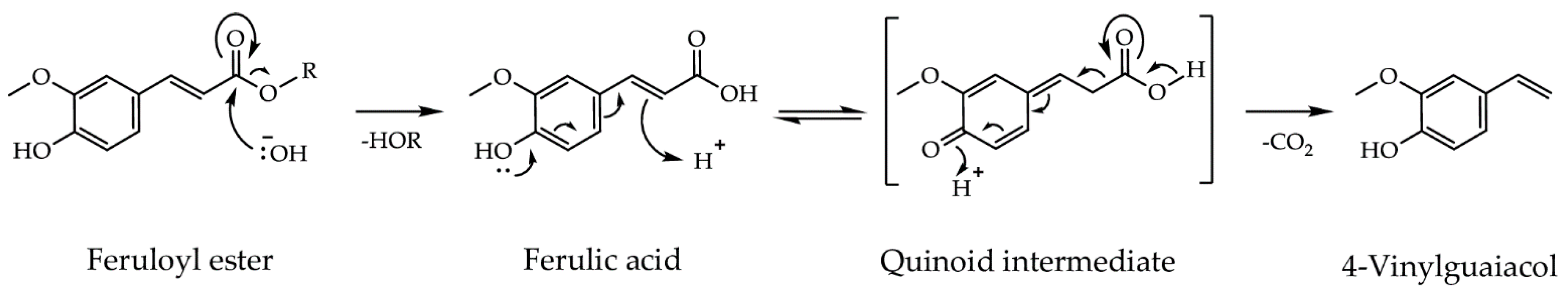
10]. Among these, ferulic acid esterases (FAEs) and decarboxylases (FADs) hydrolyse feruloyl esters, and subsequently decarboxylate ferulic acid (FA), to the smoke flavor 4-vinylguaiacol (4-VG), a reaction sequence chemically analogous to processes during pyrolysis (
Figure 1) [
11,
12]. Thus, 4-VG is an impact flavor compound in conventionally produced wood smoke [
13]. The enzymes Rs
pCAE (
Rhizoctonia solani p-coumaric acid esterase) and ScoFAD (
Schizophyllum commune ferulic acid decarboxylase) were recombinantly produced in the food grade expression host
Komagataella phaffii GS115 and were immobilized on AminoLink Agarose [
11,
12,
14].
The present work describes a small-scale prototype of a continuous process for the production of natural 4-VG with packed bed columns of immobilized enzymes and extracts of the feruloyl ester-rich sugar beet fiber as the starting material [
16,
17]. An online solid phase extraction with an XAD resin and ethanol as eluent was applied to separate 4-VG under food-grade conditions. To facilitate the process and decrease the diffusion distances between the active centers, both enzymes were joined to a His-tagged fusion protein and assessed for catalytic activity.
2. Results
2.1. Solid Phase Extraction of 4-Vinylguaiacol
For food-grade downstream processing, solid phase extraction using ethanol as eluent represents a promising and cost-efficient method. To select a suitable adsorbent, five common Amberlite XAD resins were tested with aqueous solutions of 4-VG (
Table 1a).
All resins adsorbed the analyte from Bis-Tris buffer solution well, except XAD7HP. It was the only resin, which released significant amounts of 4-VG during washing with water, resulting in an overall recovery rate of 0.75. During the elution step, all remaining adsorbents showed recovery rates close to or greater than 0.9. Based on the overall recovery rates of 4-VG during elution, XAD16N, XAD2, and XAD4 were chosen for a second screening (
Table 1b). To mimic the matrix of the planned process, 4-VG was dissolved in aqueous extracts of sugar beet fiber and was applied to the resins. In contrast to the first screening, three consecutive elution steps at 50 °C were used to maximize analyte recovery. As in the first screening, no significant amounts of 4-VG were desorbed during the washing step with water. In all three samples, the majority of the analyte (0.9 for XAD16N, XAD4 and 0.8 for XAD2) was eluted within the first elution step. The second elution released another 0.04 of the applied 4-VG from all resins, whereas the third elution did not significantly increase the overall yield. Altogether, XAD4 showed the highest recovery rate of 4-VG of 0.94 and was thus applied for the following studies.
2.2. Continuous Production of 4-VG from Sugar Beet Fiber Extracts
To model a continuous production process of 4-VG from sugar beet fiber extract, the basidiomycetous enzymes Rs
pCAE and ScoFAD were covalently immobilized on agarose. Columns equipped with the immobilisates were connected in series followed by an XAD4 column with sampling points after each column (Figure 6). The concentrations of FA (sampling valve 1) and 4-VG (sampling valves 2 and 3) were recorded for 77 h (
Figure 2a) and the corresponding conversion rates were calculated (
Figure 2b).
After an equilibration time of about 1 h, FA and 4-VG reached maximum concentrations of 380 and 265 mg L−1, respectively. After 12 h, the concentrations remained constant at about 350 (FA) and 250 mg L−1 (4-VG). During the duration of the experiment, no leakage of 4-VG through the SPE column was detected. Matching the product concentrations, the conversion rates of the applied enzymes also remained on a high level. The RspCAE exhibited an average conversion rate of about 0.8, whereas the ScoFAD reached complete conversion between 30 and 50 h and otherwise remained at an average rate of about 0.95. At the end of the experiment, close to 4000 bed volumes of substrate solution had passed each column.
From the product concentrations, a process productivity curve was calculated that showed the accumulation of 4-VG during the experiment (
Figure 3).
Described by a linear graph with a regression coefficient of 0.9999, the process offered an average productivity of 3.98 mg h−1. Overall, 306 mg 4-VG were produced through the enzymatic process of which, as shown by the missing leakage through the SPE column, was adsorbed onto the XAD4 resin.
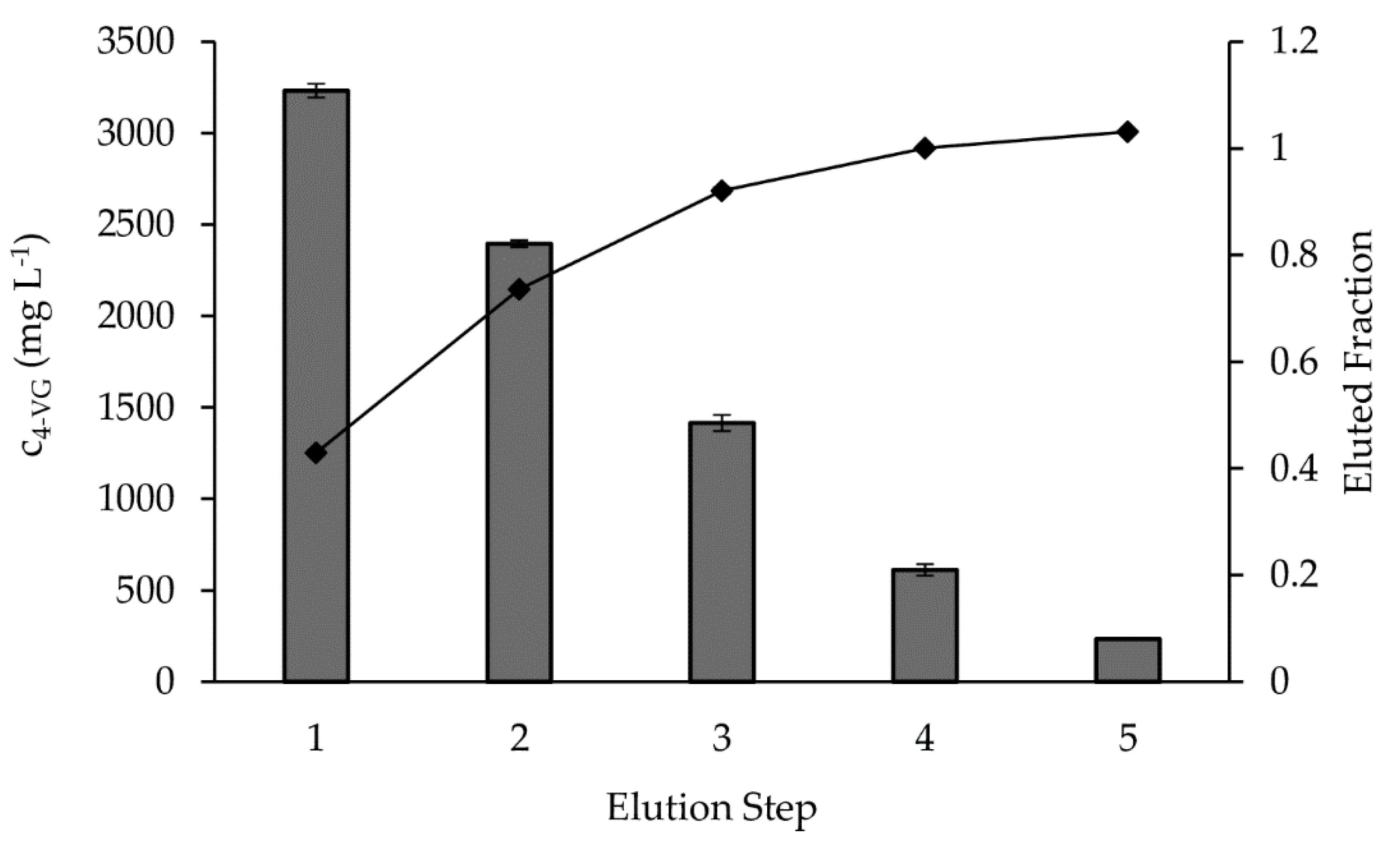
2.3. Elution of 4-Vinylguaiacol from the Solid Phase
As no leakage of 4-VG through the adsorbent column was observed, all product had bound to XAD4. To maximize the product concentration and to minimize ethanol demand, the elution volume was drastically reduced compared to the previous screenings and five subsequent elution steps were applied instead (
Figure 4).
The concentration of 4-VG in the elution fractions decreased from 3200 (fraction 1) to 200 mg L−1 (fraction 5). Calculated from the overall product mass of 306 mg, less than 50% of the total 4-VG was eluted in the first step. The accumulated proportion of eluted analyte increased with each fraction and reached 100% with the last fraction. Thus, the complete product mass was recovered. Overall, the precursor ferulic acid present in the substrate was almost completely converted, and 5.3 mg of pure 4-VG was produced per g of dry sugar beet fiber.
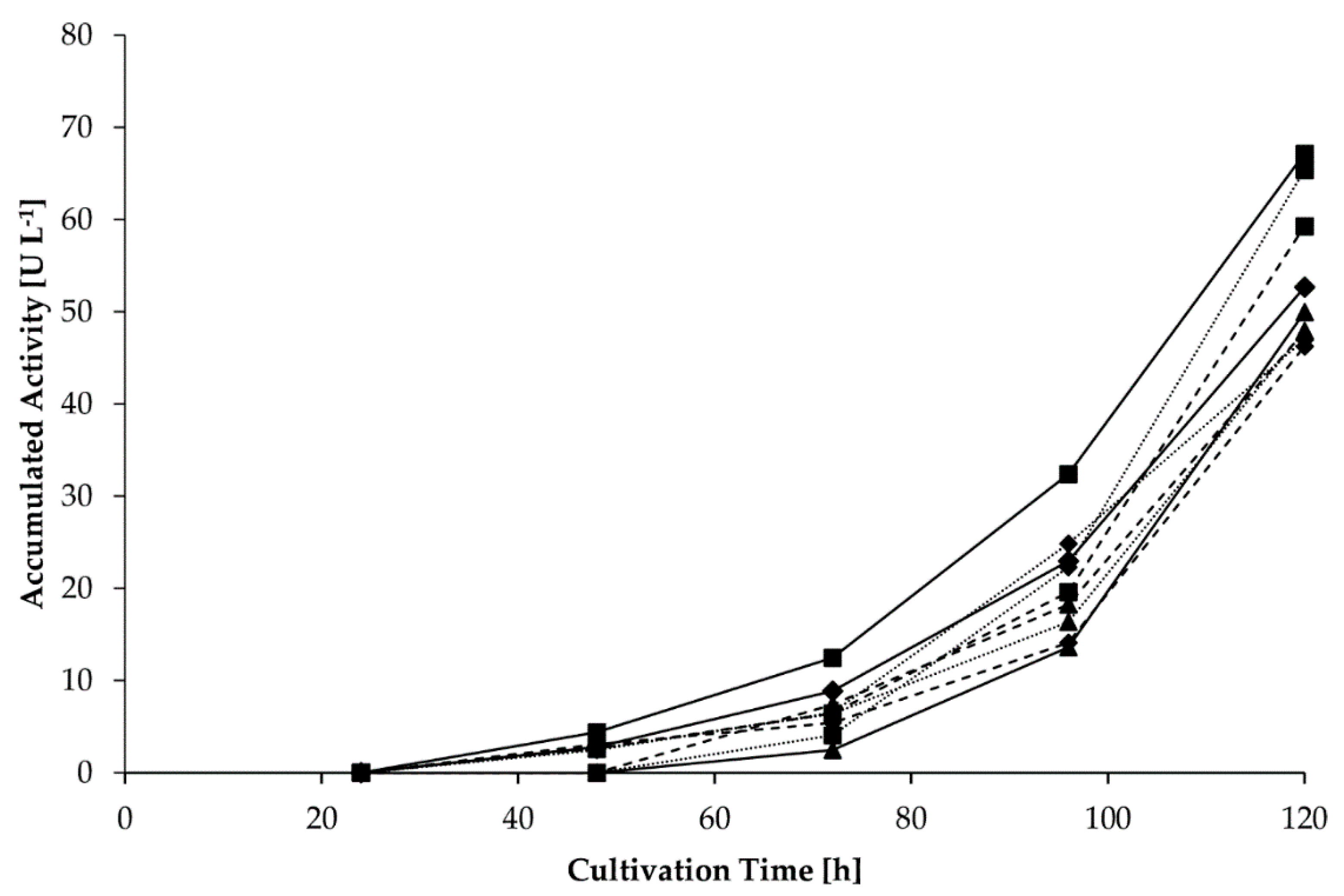
2.4. Heterologous Production of a Fusion Protein with Different Linker Variants
As the substrate is a by-product of the sugar beet industries and XAD4 is reusable, the heterologous enzymes will most likely be a major cost factor in upscaling of the presented bioprocess. Thus, a fusion protein was designed to replace the enzyme cascade. Advantages expected were the simultaneous one-column production and a process acceleration due to shorter diffusion ways between the active centers. Consequently, fusion constructs using flexible (F, GGGGS
n) and rigid (R, EAAAK
n) linkers of different lengths were cloned. As conventional cloning procedures failed, a ligase-based approach was chosen (Figure 7). After successful cloning, mixed cultures of recombinant
Komagataella phaffii were studied for their enzymatic activity to form 4-VG from methyl ferulate (
Table 2).
Activities were observed for the linker variants FFF, FFFF, RR, RRR, and RRRR. Maximal activities were detected for the linkers RRRR and RRR with 38 and 42 U L
−1, respectively. The construct with the linker variant RRRR was used for a second screening, in which the activities of individual clones were determined (
Figure 5).
A maximum activity of 67 U L−1 was detected for the clone A, producing the fusion protein with the linker RRRR after 120 h.
3. Discussion
4-VG was successfully separated from two different aqueous matrices by means of SPE using various XAD resins. The different recovery rates demonstrated the influence of the matrix on the binding behavior of 4-VG. A plausible reason for this may be the high content of saccharides and further competing agents in the sugar beet fiber extract interfering with 4-VG adsorption. Similar effects were observed by Guyot–Declerck
, et al. [
18] when extracting norisoprenoids from honey solutions. Whereas the sugar beet matrix decreased the recovery rates for the resins XAD2 and XAD16N, a higher recovery rate was detected for XAD4, which was consequently chosen for subsequent studies. XAD4 was already successfully applied for the extraction of phenols in various studies, both in native and substituted form [
19,
20]. Throughout the continuous process, no leakage of 4-VG through the resin was detected and, in repetitive elution steps, the natural flavor was quantitatively recovered in high purity under food grade conditions.
The two immobilized enzymes required to produce 4-VG from a lignified plant residue were combined for the first time. The conversion rates of the enzymes throughout the experiment remained constant on conversion rates around 0.8 (Rs
pCAE) and 0.95 (ScoFAD). Considering the highly crosslinked character of lignocellulose and comparing it to similar studies, especially the conversion rate of the esterase, was remarkably high [
21].
This was achieved as a result of the potent RspCAE, a high immobilized activity and a hydrothermal pre-treatment of the sugar beet fiber to increase the accessibility of ferulic acid.
Other studies with immobilized decarboxylases likewise showed the great potential for producing this flavor in a natural manner, but generally lacked the supply of natural ferulic acid as precursor [
22,
23]. Natural FA is predominantly used for production of high-valued natural vanillin, which also increases the market value of natural FA. This study opens the use of a cheap and lignified plant material available in large quantities to supply FA as a precursor for numerous flavor derivatives. For instance, the described bioprocess may be complemented with an oxygenase to form vanillin from 4-VG [
24].
The first active fusion protein of an esterase and a phenolic acid decarboxylase was described. The fusion proteins with longer and more rigid linkers showed the highest activities. For an economic application, the activity of the fusion protein needs to be increased, considering the activities achieved by separately used enzymes of 3300 (Rs
pCAE) and 2600 U L
−1 (ScoFAD), respectively [
11,
12]. Thus, longer and chimeric linkers combining both rigid and flexible characteristics should be tested as described by Huang, et al. [
25]
4. Materials and Methods
4.1. Applied Enzymes
For the subsequent conversion of feruloyl esters to 4-vinylguaiacol, two enzymes were required. First, a hydrolase with esterase activity to cleave feruloyl esters from off their hemicellulose backbone. The
p-coumaric acid esterase Rs
pCAE from the basidiomycete
Rhizoctonia solani, which was already successfully used as an immobilisate to reduce chlorogenic acids in coffee beverages, was found to be suitable for this purpose [
12,
14]. Second, a phenolic acid decarboxylase to cleave off carbon dioxide from the released ferulic acid to form the desired vinylic end-product. The ferulic acid decarboxylase ScoFAD from the edible fungus
Shizophyllum commune was also already successfully immobilized and showed high catalytic efficiency for the conversion of ferulic acid [
11]. Consequently, it was chosen for the second reaction step in this study. Both enzymes were available in large quantities from the food-grade expression host
Komagataella phaffii GS115.
4.2. Preparation and Elution of Solid Phase Materials
The screenings for a suitable solid phase adsorbent was performed in small scale using 10 mg of activated resin in Eppendorf tubes. All resins, including XAD2, XAD4, XAD7HP, XAD16, and XAD1180, were purchased from Sigma Aldrich (St. Luis, MO, USA). For activation, the respective adsorbent was agitated in 96% undenatured ethanol overnight at 20 °C. After removal of the ethanol, the adsorbent was washed with 1 mL double-distilled water by inversion around five times. Discarding the supernatant, 4-vinylguaiacol solved in Bis-Tris buffer solution (50 mM, pH 6) or sugar beet fiber extract at a concentration of 1 mM was loaded. Adsorption was carried out agitating overnight at 20 °C followed by a washing step applying 1 mL of double-distilled water for 1 h at 20 °C. The elution was carried out using 1 mL 96% undenatured ethanol at 20 °C for 3 h and 500 rpm in case of the buffer screening. The elution step was expanded for the screening with sugar beet fiber extracts applying three consecutive elution steps at 50 °C for 1 h and 500 rpm each. All screenings were performed in independent triplicates.
For the online extraction in the continuous bioprocess, the activation and washing steps were upscaled linearly to produce 5 g of activated XAD4. The resin was then loaded with the product of the two-enzyme-conversion for 77 h at a flow rate of 0.25 mL min−1. After washing, the XAD4 was eluted with five consecutive elution steps of adding 40 mL 96% undenaturated ethanol at 50 °C for 1 h and 500 rpm. All samples were measured in triplicates via HPLC.
4.3. Aqueous Extraction of Sugar Beet Fiber
As hemicellulose, which carries the feruloyl esters, are solvable in mild acidic conditions, aqueous extraction was performed to provide a liquid substrate solution suitable for a continuous bioprocess. Per liter of Bis-Tris buffer solution (50 mM, pH 6), 57.75 g of milled sugar beet fiber containing 8.13 mg g−1 ferulic acid were extracted in an autoclave for 2.5 h at 121 °C. The hot raw extracts were hot filtered through generous folding filters and cooled to room temperature. The concentration of total ferulic acid was determined by alkaline hydrolysis mixing 100 µL of extract and 100 µL of 4 M KOH, incubation at 50 °C for 5 h at 500 rpm, neutralization with 50 µL 60% acetic acid and adding 250 µL methanol and 500 µL acetonitrile. The samples were centrifuged at 11,000× g for 5 min and the clear supernatant was used for HPLC analysis. The extract contained 434 mg L−1 ferulic acid, which corresponded to an extraction yield of 0.92.
4.4. HPLC Analysis of Ferulic Acid and 4-Vinylguaiacol
The HPLC method for quantitating the concentration of ferulic acid and 4-vinylguaicol in the SPE screenings and in the process samples was adapted from previous works [
11]. A LC-10 system (Shimadzu Deutschland GmbH, Duisburg, Germany) was used along with a reversed phase column (Chromolith Performance RP-18e, 100 × 4.6 m
2, Merck, Darmstadt, Germany) and a SPD-10A VP UV/Vis detector (Shimadzu Deutschland GmbH, Duisburg, Germany). The wavelengths of the UV-Vis detector were set to 263 nm for the detection of 4-vinylguaiacol and 323 for ferulic acid to allow a simultaneous quantitation of both analytes.
4.5. Immobilisation of Enzymes
The enzymes were covalently immobilized to AminoLink Coupling Resin (ThermoFisher, Waltham, MA, USA) according to a method which was already found suitable for both enzymes [
11,
14]. For the continuous bioprocess, 14.1 mg agarose (dry weight) was applied for each of the enzymes and the corresponding immobilization yields were 0.28 mg mg
−1 (Rs
pCAE) and 0.44 mg mg
−1 (ScoFAD), respectively. The immobilisates formed a bed of a volume of 300 µL.
4.6. Process Management
The setup of the bioprocess included the peristaltic pump Ismatec IPC-4 (Cole Parmer GmbH, Wertheim, Germany) and three columns including both immobilized enzymes and the activated adsorbent connected via tubing (
Figure 6).
The pump rate was set to 0.25 mL min−1 and with a given bed volume of 300 µL of the immobilisates, a dwell time of 1.2 min was obtained. The use of 5 g activated adsorbent XAD4 (dry weight) resulted in a bed volume of around 10 mL, which lead to a calculated dwell time of 20 min. After each process step, a valve was placed for monitoring the concentrations of analytes. Samples were taken in regular intervals and analyzed via HPLC.
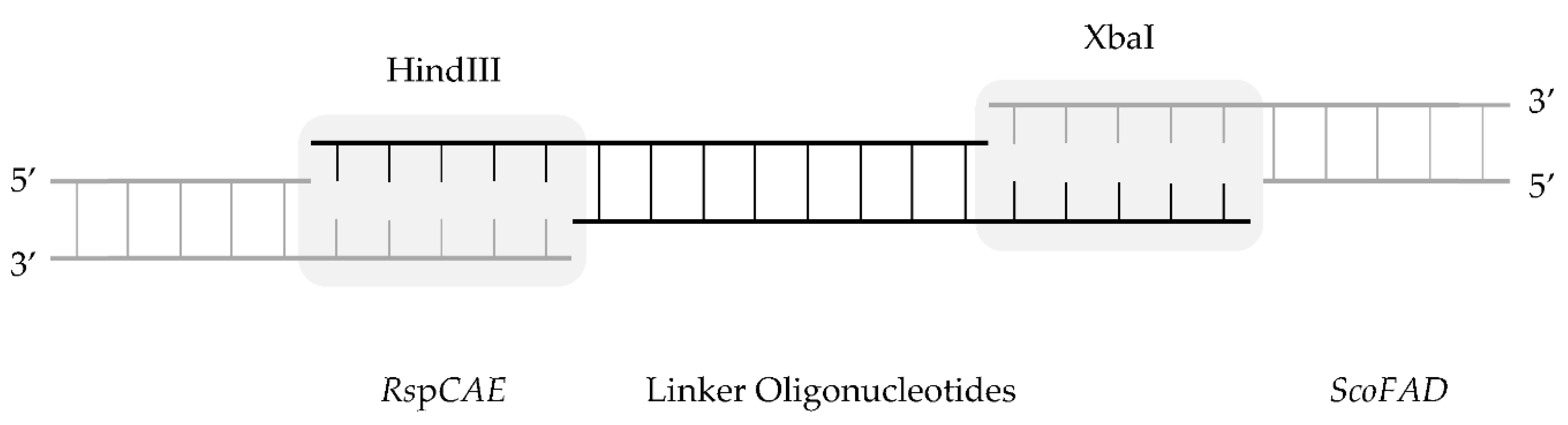
4.7. Cloning of Linker Variants of a Fusion Protein
As conventional cloning techniques failed, a ligase-based approach for the cloning of the linker variants was chosen (
Figure 7).
For each construct, the pre-digested genes of ScoFAD and RspCAE were ligated using two overlapping oligonucleotides coding for the respective linker residues. Thus, the oligosaccharides R_For 5′-AGCTTGAAGCTGCCGCAAAGT-3′ and R_Rev 5′-CTAGACTTTGCGGCAGCTTCA-3′ were applied for the cloning of the 1 × EAAAK variant, RR_For 5′-AGCTTGAAGCTGCCGCAAAGGAGGCAGCTGCCAAAT-3′ and RR_Rev 5′-CTAGATTTGGCAGCTGCCTCCTTTGCGGCAGCTTCA-3′ were applied for the cloning of the 2 × EAAAK variant, RRR_For 5′-AGCTTGAAGCTGCCGCAAAGGAGGCAGCTGCCAAAGAGGCTGCCGCAAAAT-3′ and RRR_Rev 5′-CTAGATTTTGCGGCAGCCTCTTTGGCAGCTGCCTCCTTTGCGGCAGCTTCA-3′ were applied for the cloning of the 3 × EAAAK variant, RRRR_For 5′-AGCTTGAAGCTGCCGCAAAGGAGGCAGCTGCCAAAGAGGCTGCCGCAAAAGAAGCTGCCGCAAAGT-3′ and RRRR_Rev 5′-CTAGACTTTGCGGCAGCTTCTTTTGCGGCAGCCTCTTTGGCAGCTGCCTCCTTTGCGGCAGCTTCA-3′ were applied for the cloning of the 4 × EAAAK variant, F_For 5′-AGCTTGGCGGAGGTGGAAGCT-3′ and F_Rev 5′-CTAGAGCTTCCACCTCCGCCA-3′ were applied for the cloning of the 1 × GGGGS variant, FF_For 5′-AGCTTGGCGGAGGTGGAAGCGGCGGAGGTGGAAGCT-3′ and FF_Rev 5′-CTAGAGCTTCCACCTCCGCCGCTTCCACCTCCGCCA-3′ were applied for the cloning of the 2 × GGGGS variant, FFF_For 5′-AGCTTGGCGGAGGTGGAAGCGGCGGAGGTGGAAGCGGCGGAGGTGGAAGCT-3′ and FFF_Rev 5′-CTAGAGCTTCCACCTCCGCCGCTTCCACCTCCGCCGCTTCCACCTCCGCCA-3′ were applied for the cloning of the 3 × GGGGS variant, and FFFF_For 5′-AGCTTGGTGGAGGTGGAAGCGGAGGTGGAGGTAGCGGTGGAGGTGGAAGCGGAGGTGGAGGTAGCT-3′ and FFFF_Rev 5′-CTAGAGCTACCTCCACCTCCGCTTCCACCTCCACCGCTACCTCCACCTCCGCTTCCACCTCCACCA-3′ were applied for the cloning of the 4 × GGGGS variant. The ligation was carried out at 8 °C overnight incubating 60 ng of the gene coding for RspCAE cut with HindIII, 20 ng of the gene coding for ScoFAD cut with XbaI, 5 pmol of the respective forward and reverse oligonucleotide with 5 U of T4 DNA Ligase.
The ligated constructs were subsequently blunt-end cloned in pPIC9 linearized with SnaBI and inserted into the AOXI locus of the expression host
Komagataella phaffi GS115 using electroporation as described previously [
26]. All enzymes were provided by Thermo Fisher Scientific (Waltham, MA, USA).
5. Conclusions
Lignified plant materials offer an abundant source of phenolic precursors for various biotechnological products including flavors and preservatives, which is not even close to being exhausted yet. This work presents the first comprehensive bioprocess, starting from the raw material, combining heterologously produced enzymes and online extraction, yielding a food-grade, purified and highly concentrated natural flavor compound. Based on this, novel clean and safe smoke flavors may be developed, typically in compositions with other natural flavors, which transport a smoky taste impression without any toxic by-products and using renewable resources as a starting material.




 cFA after hydrolysis with RspCAE,
cFA after hydrolysis with RspCAE,  c4-VG after decarboxylation with ScoFAD,
c4-VG after decarboxylation with ScoFAD,  c4-VG after extraction with XAD4. (b)
c4-VG after extraction with XAD4. (b)  RspCAE,
RspCAE,  ScoFAD.
ScoFAD.



 A,
A,  B,
B,  C,
C,  D,
D,  E,
E,  F,
F,  G,
G,  H,
H,  I.
I.

 ).
).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}