Response Surface Methodology for the Optimization of Keratinase Production in Culture Medium Containing Feathers by Bacillus sp. UPM-AAG1

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Isolation and Screening of Keratinase Producing Bacterium
2.2. Identification of Keratinolytic Microorganism
2.3. Optimization of Keratinase Activity Using Plackett Burman and Response Surface Methodology
2.3.1. Pre-Screening of Significant Parameters Using Plackett-Burman
2.3.2. Optimization of Significant Variables Using Central Composite Design (CCD)
2.3.3. Numerical Optimization
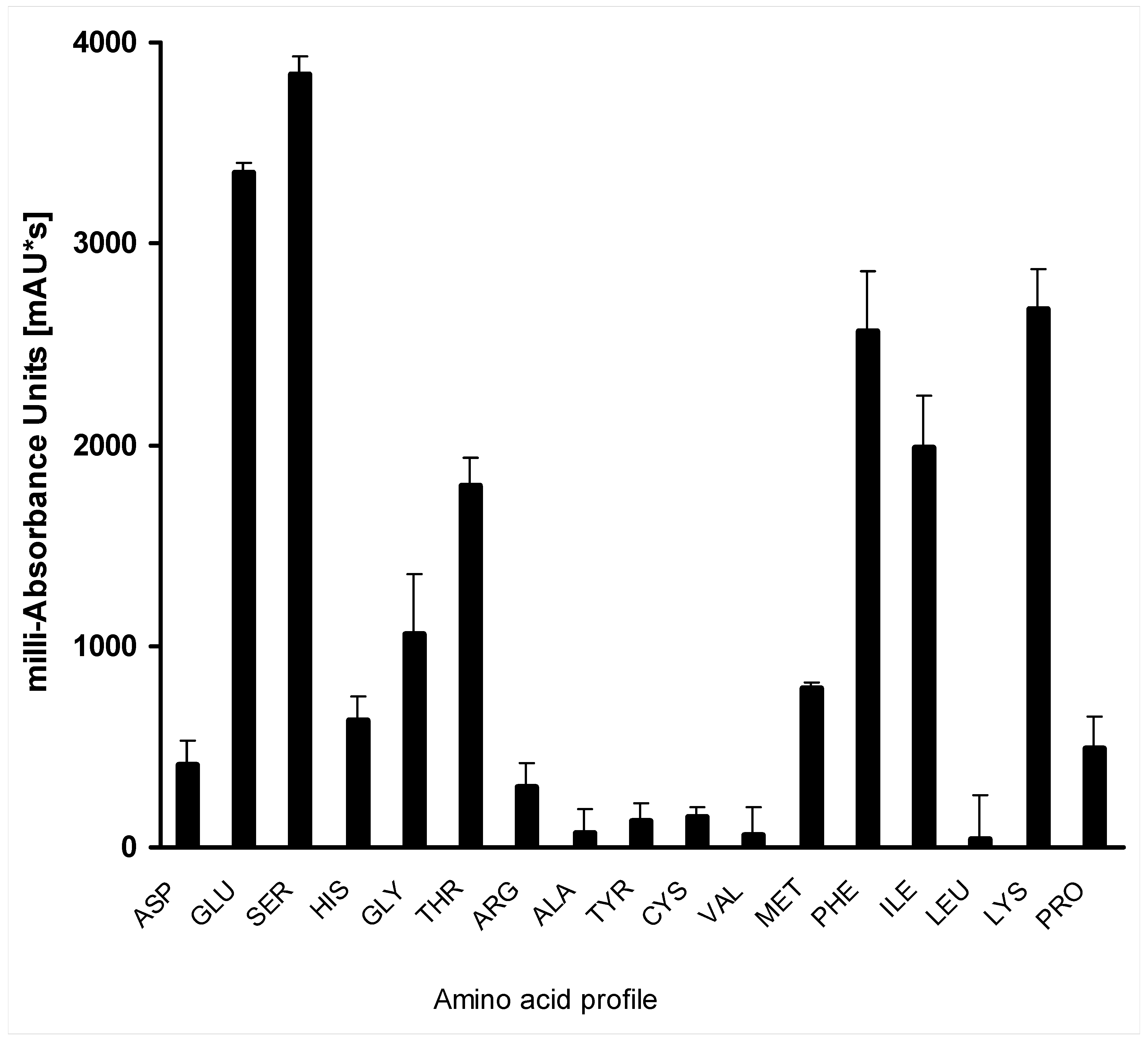
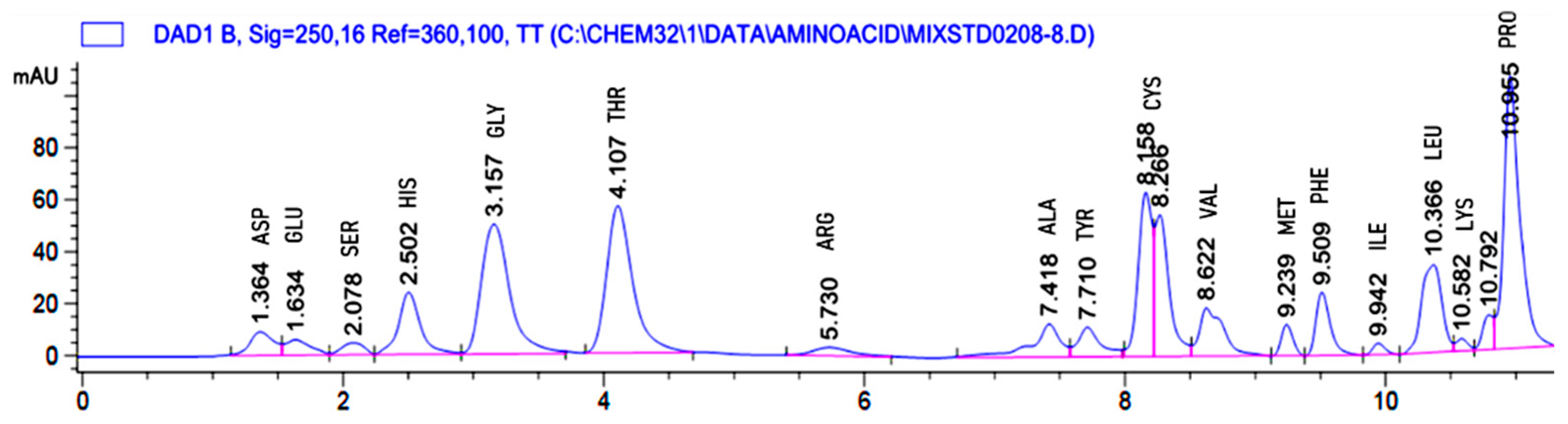
2.4. Amino Acid Profile of Hydrolysate of Bacillus sp. UPM-AAG1 Using High-Performance Liquid Chromatography (HPLC)
3. Discussion
4. Materials and Methods
4.1. Azokeratin and Keratinase Assay
4.2. Isolation and Screening of Bacillus sp. with Keratinolytic Activity
4.3. Morphological, Biochemical and Molecular Identification of Keratinolytic Microorganism
4.4. Sequence Analysis and Phylogenetic Analysis
4.5. Optimization of Keratinase Activity Using Response Surface Methodology
4.6. Amino Acid Profile of Hydrolysate
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Fang, Z.; Zhang, J.; Liu, B.; Du, G.; Chen, J. Biodegradation of wool waste and keratinase production in scale-up fermenter with different strategies by Stenotrophomonas maltophilia BBE11-1. Bioresour. Technol. 2013, 140, 286–291. [Google Scholar] [CrossRef] [Green Version]
- Brandelli, A.; Daroit, D.J.; Riffel, A. Biochemical features of microbial keratinases and their production and applications. Appl. Microbiol. Biotechnol. 2010, 85, 1735–1750. [Google Scholar] [CrossRef] [PubMed]
- Tiwary, E.; Gupta, R. Rapid Conversion of Chicken Feather to Feather Meal Using Dimeric Keratinase from Bacillus licheniformis ER-15. J. Bioprocess. Biotech. 2012, 2, 1000123. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Fattah, A.M.; El-Gamal, M.S.; Ismail, S.; Emran, M.; Hashem, A. Biodegradation of feather waste by keratinase produced from newly isolated Bacillus licheniformis ALW1. J. Genet. Eng. Biotechnol. 2018, 16, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Zhang, J.; Liu, B.; Jiang, L. Cloning, heterologous expression and characterization of two keratinases from Stenotrophomonas maltophilia BBE11-1. Process Biochem. 2014, 49, 647–654. [Google Scholar] [CrossRef]
- Pereira, J.Q.; Lopes, F.C.; Petry, M.V.; da Costa Medina, L.F.; Brandelli, A. Isolation of three novel Antarctic psychrotolerant feather-degrading bacteria and partial purification of keratinolytic enzyme from Lysobacter sp. A03. Int. Biodeterior. Biodegrad. 2014, 88, 1–7. [Google Scholar] [CrossRef]
- Mohamedin, A.H. Isolation, identification and some cultural conditions of a protease- producing thermophilic Streptomyces strain grown on chicken feather as a substrate. Int. Biodeterior. Biodegrad. 1999, 43, 13–21. [Google Scholar] [CrossRef]
- Syeda, D.G.; Leeb, J.C.; Lic, W.-J.; Kimb, C.-J.; Agasard, D. Production, characterization and application of keratinase from Streptomyces gulbargensis. Bioresour. Technol. 2009, 100, 1868–1871. [Google Scholar] [CrossRef]
- Malviya, H.K.; Rajak, R.C.; Hasija, S.K. Synthesis and regulation of extracellular keratinase in three fungi isolated from the grounds of a gelatin factory, Jabalpur, India. Mycopathologia 1992, 120, 1–4. [Google Scholar] [CrossRef]
- Ramakrishnaiah, G.; Mustafa, S.M.; Srihari, G. Studies on Keratinase Producing Fungi Isolated from Poultry Waste and their Enzymatic Activity. Microbiol. Res. 2013, 3, 148–151. [Google Scholar] [CrossRef]
- Lin, X.; Chung, G.L.; Casale, E.S.; Jason, C.H.S. Purification and characterization of a keratinase from a degrading Bacillus licheniformis strain. Appl. Environ. Microbiol. 1992, 58, 3271–3275. [Google Scholar] [CrossRef] [Green Version]
- Suntornsuk, W.; Suntornsuk, L. Feather degradation by Bacillus sp. FK 46 in submerged cultivation. Bioresour. Technol. 2003, 86, 239–243. [Google Scholar] [CrossRef]
- Schallmey, M.; Singh, A.; Ward, O.P. Developments in the use of Bacillus species for industrial production. Can. J. Microbiol. 2004, 50, 1–17. [Google Scholar] [CrossRef]
- Huang, Y.; Busk, P.K.; Herbst, F.-A.; Lange, L. Genome and secretome analyses provide insights into keratin decomposition by novel proteases from the non-pathogenic fungus Onygena corvina. Appl. Microbiol. Biotechnol. 2015, 99, 9635–9649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Refai, H.A.; AbdelNaby, M.A.; Gaballa, A.; El-Araby, M.H.; Abdel Fattah, A.F. Improvement of the newly isolated Bacillus pumilus FH9 keratinolytic activity. Process Biochem. 2005, 40, 2325–2332. [Google Scholar] [CrossRef]
- Reddy, L.V.A.; Wee, Y.J.; Yun, J.S.; Ryu, H.W. Optimization of alkaline protease production by batch culture of Bacillus sp. RKY3 through Plackett-Burman and response surface methodological approaches. Bioresour. Technol. 2008, 99, 2242–2249. [Google Scholar] [CrossRef]
- Pillai, P.; Mandge, S.; Archana, G. Statistical optimization of production and tannery applications of a keratinolytic serine protease from Bacillus subtilis P13. Process Biochem. 2011, 46, 1110–1117. [Google Scholar] [CrossRef]
- Dettmer, A.; Cavalli, É.; Ayub, M.A.Z.; Gutterres, M. Optimization of the unhairing leather processing with enzymes and the evaluation of inter-fibrillary proteins removal: An environment-friendly alternative. Bioprocess Biosyst. Eng. 2012, 35, 1317–1324. [Google Scholar] [CrossRef] [PubMed]
- Lo, W.H.; Too, J.R.; Wu, J.Y. Production of keratinolytic enzyme by an indigenous feather-degrading strain Bacillus cereus WU2. J. Biosci. Bioeng. 2012, 114, 640–647. [Google Scholar] [CrossRef]
- Okoroma, E.A.; Garelick, H.; Abiola, O.O.; Purchase, D. Identification and characterisation of a Bacillus licheniformis strain with profound keratinase activity for degradation of melanised feather. Int. Biodeterior. Biodegrad. 2012, 74, 54–60. [Google Scholar] [CrossRef]
- Sahoo, D.K.; Das, A.; Thatoi, H.; Mondal, K.C.; Mohapatra, P.K.D. Keratinase production and biodegradation of whole chicken feather keratin by a newly isolated bacterium under submerged fermentation. Appl. Biochem. Biotechnol. 2012, 167, 1040–1051. [Google Scholar] [CrossRef] [PubMed]
- Sivakumar, T.; Shankar, T.; Vijayabaskar, P.; Ramasubramanian, V. Optimization for Keratinase Enzyme Production Using Bacillus thuringiensis TS2. Acad. J. Plant Sci. 2012, 5, 102–109. [Google Scholar] [CrossRef]
- E Silva, L.A.D.; Macedo, A.J.; Termignoni, C. Production of keratinase by Bacillus subtilis S14. Ann. Microbiol. 2014, 64, 1725–1733. [Google Scholar] [CrossRef]
- Govarthanan, M.; Selvankumar, T.; Selvam, K.; Sudhakar, C.; Kamala-Kannan, S. Response surface methodology based optimization of keratinase production from alkali-treated feather waste and horn waste using Bacillus sp. MG-MASC-BT. J. Ind. Eng. Chem. 2015, 27, 25–30. [Google Scholar] [CrossRef]
- Sanghvi, G.; Patel, H.; Vaishnav, D.; Oza, T.; Dave, G.; Kunjadia, P.; Sheth, N. A novel alkaline keratinase from Bacillus subtilis DP1 with potential utility in cosmetic formulation. Int. J. Biol. Macromol. 2016, 87, 256–262. [Google Scholar] [CrossRef]
- Reddy, M.R.; Reddy, K.S.; Chouhan, Y.R.; Bee, H.; Reddy, G. Effective feather degradation and keratinase production by Bacillus pumilus GRK for its application as bio-detergent additive. Bioresour. Technol. 2017, 243, 254–263. [Google Scholar] [CrossRef]
- Lateef, A.; Oloke, J.K.; Gueguim Kana, E.B.; Sobowale, B.O.; Ajao, S.O.; Bello, B.Y. Keratinolytic activities of a new feather-degrading isolate of Bacillus cereus LAU 08 isolated from Nigerian soil. Int. Biodeterior. Biodegrad. 2010, 64, 162–165. [Google Scholar] [CrossRef]
- Kothari, D.; Rani, A.; Goyal, A. Keratinase. In Current Developments in Biotechnology and Bioengineering: Production, Isolation and Purification of Industrial Products; Elsevier: London, UK, 2017; pp. 447–469. ISBN 9780444636621. [Google Scholar]
- Walker, R.; Powell, A.A.; Seddon, B. Bacillus isolates from the spermosphere of peas and dwarf French beans with antifungal activity against Botrytis cinerea and Pythium species. J. Appl. Microbiol. 1998, 84, 791–801. [Google Scholar] [CrossRef]
- Monteiro, S.M.; Clemente, J.; Henriques, A.O.; Gomes, R.J.; Carrondo, M.J. A Procedure for High-Yield Spore Production by Bacillus subtilis. Biotechnol. Prog. 2005, 21, 1026–1031. [Google Scholar] [CrossRef]
- Tan, I.S.; Ramamurthi, K.S. Spore formation in Bacillus subtilis. Environ. Microbiol. Rep. 2014, 6, 212–225. [Google Scholar] [CrossRef] [Green Version]
- Riffel, A.; Brandelli, A. Keratinolytic bacteria isolated from feather waste. Braz. J. Microbiol. 2006, 37, 395–399. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.X.; Gong, J.S.; Su, C.; Zhang, D.D.; Tian, H.; Dou, W.F.; Li, H.; Shi, J.S.; Xu, Z.H. Biochemical characterization of a novel surfactant-stable serine keratinase with no collagenase activity from Brevibacillus parabrevis CGMCC 10798. Int. J. Biol. Macromol. 2016, 93, 843–851. [Google Scholar] [CrossRef]
- Laba, W.; Choinska, A.; Rodziewicz, A.; Piegza, M.; Laba, W.; Choinska, A.; Rodziewicz, A.; Piegza, M. Keratinolytic abilities of Micrococcus luteus from poultry waste. Braz. J. Microbiol. 2015, 46, 691–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arokiyaraj, S.; Varghese, R.; Ahmed, B.A.; Duraipandiyan, V.; Al-dhabi, N.A. Optimizing the fermentation conditions and enhanced production of keratinase from Bacillus cereus isolated from halophilic environment. Saudi J. Biol. Sci. 2019, 26, 378–381. [Google Scholar] [CrossRef] [PubMed]
- Vidmar, B.; Vodovnik, M. Microbial Keratinases: Enzymes with Promising Biotechnological Applications. Food Technol. Biotechnol. 2018, 56, 312–328. [Google Scholar] [CrossRef]
- Khodayari, S.; Kafilzadeh, F. Separating Keratinase Producer Bacteria from the Soil of Poultry Farms and Optimization of the Conditions for Maximum Enzyme Production. Eur. J. Exp. Biol. 2018, 8, 1–8. [Google Scholar] [CrossRef]
- Fang, Z.; Zhang, J.; Liu, B.; Du, G.; Chen, J. Biochemical characterization of three keratinolytic enzymes from Stenotrophomonas maltophilia BBE11-1 for biodegrading keratin wastes. Int. Biodeterior. Biodegrad. 2013, 82, 166–172. [Google Scholar] [CrossRef]
- Fakhfakh-Zouari, N.; Haddar, A.; Hmidet, N.; Frikha, F.; Nasri, M. Application of statistical experimental design for optimization of keratinases production by Bacillus pumilus A1 grown on chicken feather and some biochemical properties. Process Biochem. 2010, 45, 617–626. [Google Scholar] [CrossRef]
- Cai, C.; Zheng, X. Medium optimization for keratinase production in hair substrate by a new Bacillus subtilis KD-N2 using response surface methodology. J. Ind. Microbiol. Biotechnol. 2009, 36, 875–883. [Google Scholar] [CrossRef]
- Kumar, R.; Balaji, S.; Uma, T.S. Optimization of influential parameters for extracellular keratinase production by Bacillus subtilis (MTCC9102) in solid state fermentation using horn meal -A biowaste management. Appl. Biochem. Biotechnol. 2010, 160, 30–39. [Google Scholar] [CrossRef]
- Mazotto, A.M.; Cedrola, S.M.L.; Lins, U.; Rosado, A.S.; Silva, K.T.; Chaves, J.Q.; Rabinovitch, L. Keratinolytic activity of Bacillus subtilis AMR using human hair. Soc. Appl. Microbiol. 2010, 50, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Tiwary, E.; Gupta, R. Medium optimization for a novel 58 kDa dimeric keratinase from Bacillus licheniformis ER-15: Biochemical characterization and application in feather degradation and dehairing of hides. Bioresour. Technol. 2010, 101, 6103–6110. [Google Scholar] [CrossRef] [PubMed]
- Fakhfakh, N.; Kanoun, S.; Manni, L.; Nasri, M. Production and biochemical and molecular characterization of a keratinolytic serine protease from chicken feather-degrading Bacillus licheniformis RPk. Can. J. Microbiol. 2009, 55, 427–436. [Google Scholar] [CrossRef]
- Mazotto, A.M.; Coelho, R.R.R.; Cedrola, S.M.L.; De Lima, M.F.; Couri, S.; Paraguai De Souza, E.; Vermelho, A.B. Keratinase production by three Bacillus spp. using feather meal and whole feather as substrate in a submerged fermentation. Enzyme Res. 2011, 2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, R.; Beg, Q.; Lorenz, P. Bacterial alkaline proteases: Molecular approaches and industrial applications. Appl. Microbiol. Biotechnol. 2005, 59, 15–32. [Google Scholar] [CrossRef]
- Mousavi, S.; Salouti, M.; Shapoury, R.; Heidari, Z. Optimization of keratinase production for feather degradation by Bacillus subtilis. Jundishapur J. Microbiol. 2013, 6. [Google Scholar] [CrossRef] [Green Version]
- Yusuf, I.; Ahmad, S.A.; Phang, L.Y.; Syed, M.A.; Shamaan, N.A.; Abdul Khalil, K.; Dahalan, F.A.; Shukor, M.Y. Keratinase production and biodegradation of polluted secondary chicken feather wastes by a newly isolated multi heavy metal tolerant bacterium-Alcaligenes sp. AQ05-001. J. Environ. Manag. 2016, 183, 182–195. [Google Scholar] [CrossRef]
- Covino, S.; D’Annibale, A.; Stazi, S.R.; Cajthaml, T.; Čvančarová, M.; Stella, T.; Petruccioli, M. Assessment of degradation potential of aliphatic hydrocarbons by autochthonous filamentous fungi from a historically polluted clay soil. Sci. Total Environ. 2015, 505, 545–554. [Google Scholar] [CrossRef]
- Riffel, A.; Lucas, F.; Heeb, P.; Brandelli, A. Characterization of a new keratinolytic bacterium that completely degrades native feather keratin. Arch. Microbiol. 2003, 179, 258–265. [Google Scholar] [CrossRef]
- Pillai, P.; Archana, G. Hide depilation and feather disintegration studies with keratinolytic serine protease from a novel Bacillus subtilis isolate. Appl. Microbiol. Biotechnol. 2008, 78, 643–650. [Google Scholar] [CrossRef]
- Balaji, S.; Kumar, M.S.; Karthikeyan, R.; Kumar, R.; Kirubanandan, S.; Sridhar, R.; Sehgal, P.K. Purification and characterization of an extracellular keratinase from a hornmeal-degrading Bacillus subtilis MTCC (9102). World J. Microbiol. Biotechnol. 2008, 24, 2741–2745. [Google Scholar] [CrossRef]
- Bansal, M.; Sudhakara Reddy, M.; Kumar, A. Optimization of cell growth and bacoside—A production in suspension cultures of Bacopa monnieri (L.) Wettst. using response surface methodology. In Vitro Cell. Dev. Biol. Plant 2017, 53, 527–537. [Google Scholar] [CrossRef]
- Whitcomb, P.J.; Anderson, M.J. RSM Simplified: Optimizing Processes Using Response Surface Methods for Design of Experiments; Productivity Press: New York, NY, USA, 2004; ISBN 978-1-56327-297-4. [Google Scholar]
- Manogaran, M.; Shukor, M.Y.; Yasid, N.A.; Johari, W.L.W.; Ahmad, S.A. Isolation and characterisation of glyphosate-degrading bacteria isolated from local soils in Malaysia. Rend. Lincei 2017, 28, 471–479. [Google Scholar] [CrossRef]
- Roslan, M.A.H.; Abdullah, N.; Mustafa, S. Removal of shells in palm kernel cake via static cling and electrostatic separation. J. Biochem. Microbiol. Biotechnol. 2015, 3, 1–6. [Google Scholar]
- Aziz, N.F.; Halmi, M.I.E.; Johari, W.L.W. Statistical optimization of hexavalent molybdenum reduction by Serratia sp. strain MIE2 using Central Composite Design (CCD). J. Biochem. Microbiol. Biotechnol. 2017, 5, 8–11. [Google Scholar]
- Richa, K.; Bose, H.; K, S.; Loganathan, K.; Kumar, G.; Rao, B. Response surface optimization for the production of marine eubacterial protease and its application. Res. J. Biotechnol. 2013, 8, 78–85. [Google Scholar]
- Ghosh, A.; Chakrabarti, K.; Chattopadhyay, D. Degradation of raw feather by a novel high molecular weight extracellular protease from newly isolated Bacillus cereus DCUW. J. Ind. Microbiol. Biotechnol. 2008, 35, 825–834. [Google Scholar] [CrossRef]
- Joshi, S.G.; Tejashwini, M.M.; Revati, N.; Sridevi, R.; Roma, D. Isolation, identification and characterization of feather degrading bacteria. Int. Journall Poult. Sci. 2007, 6, 689–693. [Google Scholar] [CrossRef] [Green Version]
- Dulbecco, R.; Vogt, M. Plaque formation and isolation of pure lines with poliomyelitis viruses. J. Exp. Med. 1954, 99, 167–182. [Google Scholar] [CrossRef] [Green Version]
- Gajbhiye, A.; Rai, A.R.; Meshram, S.U.; Dongre, A.B. Isolation, evaluation and characterization of Bacillus subtilis from cotton rhizospheric soil with biocontrol activity against Fusarium oxysporum. World J. Microbiol. Biotechnol. 2010, 26, 1187–1194. [Google Scholar] [CrossRef]
- Schaeffer, A.B.; Fulton, M.D. A simplified methode of staining endospores. Science 1933, 77, 1990. [Google Scholar] [CrossRef] [PubMed]
- Cappuccino, J.; Sherman, N. Microbiology: A Laboratory Manual, 10th ed.; Pearson: London, UK, 2010. [Google Scholar]
- Habib, S.; Ahmad, S.A.; Johari, W.L.W.; Shukor, M.Y.A.; Alias, S.A.; Khalil, K.A.; Yasid, N.A. Evaluation of conventional and response surface level optimisation of n-dodecane (n-C12) mineralisation by psychrotolerant strains isolated from pristine soil at Southern Victoria Island, Antarctica. Microb. Cell Factories 2018, 17, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saitou, N.; Nei, M. The Neighbor-joining Method: A New Method for Reconstructing Phylogenetic Trees’. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Jukes, T.H.; Cantor, C.R. Evolution of protein molecules. In Mammalian Protein Metabolism; Munro, N., Ed.; Academic Press: New York, NY, USA, 1969; Volume 3, pp. 21–132. [Google Scholar]
- Jones, B.N.; Gilligan, J.P. o-phthaldialdehyde precolumn derivatization and reversed-phase high-performance liquid chromatography of polypeptide hydrolysates and physiological fluids. J. Chromatogr. A 1983, 266, 471–482. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Morphology |
|---|---|
| UPM-AAG1 | Circular, White, Dry, Flat |
| UPM-AAG6 | Irregular, Dry, White, Flat |
| UPM-AAG14 | Irregular, Dry, White, Flat |
| UPM-AAG15 | Irregular, Dry, White, Flat |
| UPM-AAG16 | Irregular, Dry, White, Flat |
| Run | Factors | ||||
|---|---|---|---|---|---|
| Temperature (°C) | Inoculum (v/v) | pH | Feather Concentration (w/v) | Keratinase Activity (U/mL) | |
| A | B | C | D | ||
| 1 | 25.00 | 10.00 | 5.00 | 1.00 | 6.4 ± 0.25 |
| 2 | 25.00 | 10.00 | 8.00 | 5.00 | 8.7 ± 0.07 |
| 3 | 35.00 | 5.00 | 8.00 | 1.00 | 7.6 ± 0.13 |
| 4 | 35.00 | 10.00 | 5.00 | 5.00 | 9.7 ± 0.26 |
| 5 | 35.00 | 10.00 | 8.00 | 1.00 | 7.7 ± 0.01 |
| 6 | 35.00 | 10.00 | 5.00 | 5.00 | 9.3 ± 0.11 |
| 7 | 25.00 | 5.00 | 5.00 | 1.00 | 8.3 ± 0.14 |
| 8 | 25.00 | 5.00 | 5.00 | 5.00 | 12.5 ± 0.28 |
| 9 | 25.00 | 10.00 | 8.00 | 1.00 | 5.8 ± 0.1 |
| 10 | 35.00 | 5.00 | 5.00 | 1.00 | 12 ± 0.16 |
| 11 | 25.00 | 5.00 | 8.00 | 5.00 | 10.5 ± 0.01 |
| 12 | 35.00 | 5.00 | 8.00 | 5.00 | 7.8 ± 0.26 |
| Source | Factors | F-Value | p-Value | Remarks | |
| Model | 70.33 | 0.0025 | Significant | ||
| A | Temperature | 16.98 | 0.0259 | Significant | |
| B | Inoculum | 63.75 | 0.0041 | Significant | |
| C | pH | 179.38 | 0.0009 | Significant | |
| D | Feather | 72.24 | 0.0034 | Significant | |
| Value | |||||
| R2 | 0.9947 | ||||
| Adjusted R2 | 0.9806 | ||||
| Predicted R2 | 0.8955 | ||||
| Adeq Precision | 26.980 | ||||
| Run Order | X1: Temperature | X2: Inoculum | X3: pH | X4: Feather (w/v) | Keratinase Activity (U/mL) | |
|---|---|---|---|---|---|---|
| Experimental Value | Predicted Value | |||||
| 1 | 35.00 | 5.00 | 5.50 | 5.00 | 42 ± 1.87 | 42.42 |
| 2 | 25.00 | 5.00 | 5.50 | 5.00 | 42.3 ± 1.75 | 46.18 |
| 3 | 30.00 | 7.50 | 6.75 | 3.00 | 41.1 ± 1.74 | 38.85 |
| 4 | 30.00 | 7.50 | 6.75 | 3.00 | 48 ± 0.28 | 48 |
| 5 | 35.00 | 5.00 | 5.50 | 1.00 | 22.5 ± 1.62 | 22.56 |
| 6 | 35.00 | 10.00 | 8.00 | 1.00 | 31.9 ± 1.57 | 29.97 |
| 7 | 35.00 | 10.00 | 5.50 | 5.00 | 38.4 ± 0.76 | 40.99 |
| 8 | 30.00 | 7.50 | 4.25 | 3.00 | 55.7 ± 0.49 | 53.8 |
| 9 | 25.00 | 10.00 | 8.00 | 5.00 | 50.5 ± 1.81 | 54.68 |
| 10 | 35.00 | 5.00 | 8.00 | 1.00 | 56.6 ± 0.51 | 54.34 |
| 11 | 25.00 | 5.00 | 8.00 | 1.00 | 14.9 ± 0.76 | 17.15 |
| 12 | 35.00 | 10.00 | 8.00 | 5.00 | 20 ± 1.02 | 22.21 |
| 13 | 25.00 | 5.00 | 8.00 | 5.00 | 33.1 ± 0.43 | 33.42 |
| 14 | 20.00 | 7.50 | 6.75 | 3.00 | 32.2 ± 0.86 | 36.73 |
| 15 | 40.00 | 7.50 | 6.75 | 3.00 | 19.5 ± 0.54 | 17.9 |
| 16 | 35.00 | 5.00 | 8.00 | 5.00 | 26.7 ± 0.86 | 26.6 |
| 17 | 30.00 | 12.50 | 6.75 | 3.00 | 20.2 ± 1.91 | 18.52 |
| 18 | 25.00 | 10.00 | 5.50 | 5.00 | 31.9 ± 0.11 | 30.98 |
| 19 | 30.00 | 7.50 | 6.75 | -1.00 | 60.1 ± 1 | 56.8 |
| 20 | 25.00 | 10.00 | 8.00 | 1.00 | 42.4 ± 0.61 | 43.1 |
| 21 | 25.00 | 5.00 | 5.50 | 1.00 | 47.4 ± 0.84 | 44.48 |
| 22 | 30.00 | 2.50 | 6.75 | 3.00 | 28.7 ± 0.54 | 29.02 |
| 23 | 30.00 | 7.50 | 6.75 | 3.00 | 41.6 ± 1.12 | 42.47 |
| 24 | 35.00 | 10.00 | 5.50 | 1.00 | 31 ± 1.11 | 27.53 |
| 25 | 30.00 | 7.50 | 6.75 | 3.00 | 45.1 ± 0.75 | 49.2 |
| 26 | 30.00 | 7.50 | 6.75 | 3.00 | 44 ± 1.87 | 49.2 |
| 27 | 30.00 | 7.50 | 6.75 | 3.00 | 50.6 ± 0.8 | 49.2 |
| 28 | 30.00 | 7.50 | 9.25 | 3.00 | 52.4 ± 1.12 | 49.2 |
| 29 | 30.00 | 7.50 | 6.75 | 7.00 | 51.1 ± 1.69 | 49.2 |
| 30 | 25.00 | 10.00 | 5.50 | 1.00 | 52 ± 1.67 | 49.2 |
| Source | Sum of Squares | DF | Mean Square | F Value | Prob > F | |
|---|---|---|---|---|---|---|
| Model | 4308.841 | 14 | 307.7743 | 23.79177 | <0.0001 | significant |
| A | 191.1947 | 1 | 191.1947 | 14.77986 | 0.0016 | |
| B | 21.89327 | 1 | 21.89327 | 1.692408 | 0.2129 | |
| C | 235.7161 | 1 | 235.7161 | 18.22148 | 0.0007 | |
| D | 1089.714 | 1 | 1089.714 | 84.23776 | <0.0001 | |
| A2 | 1024.804 | 1 | 1024.804 | 79.22009 | <0.0001 | |
| B2 | 265.7186 | 1 | 265.7186 | 20.54075 | 0.0004 | |
| C2 | 0.964286 | 1 | 0.964286 | 0.074542 | 0.7886 | |
| D2 | 345.6686 | 1 | 345.6686 | 26.7211 | 0.0001 | |
| AB | 13.3225 | 1 | 13.3225 | 1.029865 | 0.3263 | |
| AC | 29.16 | 1 | 29.16 | 2.254145 | 0.1540 | |
| AD | 16.81 | 1 | 16.81 | 1.299458 | 0.2722 | |
| BC | 484 | 1 | 484 | 37.41448 | <0.0001 | |
| BD | 1.96 | 1 | 1.96 | 0.151513 | 0.7026 | |
| CD | 1152.603 | 1 | 1152.603 | 89.09923 | <0.0001 | |
| Residual | 194.0425 | 15 | 12.93617 | |||
| Lack of Fit | 126.5425 | 10 | 12.65425 | 0.937352 | 0.5669 | not significant |
| Pure Error | 67.5 | 5 | 13.5 | |||
| Cor Total | 4502.883 | 29 | ||||
| Std. Dev. | 3.596688 | R-Squared | 0.956907 | |||
| Mean | 39.13 | Adj R-Squared | 0.916687 | |||
| C.V. | 9.191639 | Pred R-Squared | 0.816543 | |||
| PRESS | 826.0848 | Adeq Precision | 15.58869 |
| Lower | Upper | Lower | Upper | |||
|---|---|---|---|---|---|---|
| Name | Goal | Limit | Limit | Weight | Weight | Importance |
| Temperature | minimize | 25 | 35 | 1 | 1 | 3 |
| pH | is in range | 5.5 | 8 | 1 | 1 | 3 |
| Inoculum | minimize | 5 | 10 | 1 | 1 | 3 |
| Feather | maximize | 1 | 5 | 1 | 1 | 3 |
| Keratinase Activity | maximize | 14.9 | 60.1 | 1 | 1 | 5 |
| Number | Temperature | pH | Inoculum | Feather | Keratinase Activity | Desirability |
|---|---|---|---|---|---|---|
| 1 | 25.05 | 7.00 | 5.00 | 5.00 | 56.218 | 0.968 |
| Organism | Optimization Model | Optimum Temperature (°C) | Keratinase Activity (U/mL) | Substrate during Optimization | Carbon Sources during Optimization | Nitrogen Sources during Optimization | Time (h) | References |
|---|---|---|---|---|---|---|---|---|
| Bacillus pumilus A1 | OFAT Plackett-Burman CCD | 30 | 87.73 U/mL | heat-treated chicken feather meal | heat-treated chicken feather meal | peptone | 24 | [39] |
| Bacillus subtilis P13 | OFAT Plackett-Burman Box–Behnken | room | 2.07 U/mL | soybean meal | soybean meal | soybean meal | 24 | [17] |
| Bacillus sp. RKY3 | Plackett-Burman CCD | -ns- | 939 U/mL | corn starch, corn steep liquor | corn starch | corn steep liquor | 24 | [16] |
| Bacillus thuringiensis TS2 | OFAT | 50 | 90.78 U/mL | feather meal | starch | yeast extract | 96 | [22] |
| B. Subtilis KD-N2 | OFAT | 23 | 60.9 U/mL | feather | sucrose | feather | 30 | [40] |
| Bacillus subtilis DP1 | OFAT | 37 | 379.65 U/mL | feather coffee | feather coffee | feather coffee | 96 | [25] |
| Bacillus subtilis (MTCC9102) | OFAT | 37 | 15.972 U/mL | horn meal | dextrose | peptone | 48 | [41] |
| Bacillus licheniformis ALW1 | OFAT | 65 | 72.2 U/mL | native feather | galactose | corn steep liquor | 96 | [4] |
| Bacillus pumilus GRK | OFAT | 37 | 373 U/mL | feather | feather | feather | 24 | [26] |
| Bacillus subtilis AMR | OFAT | 50 | 163 U/mL | human hair | yeast extract | yeast extract | 192 | [42] |
| Bacillus cereus LAU 08 | OFAT | 50 | 51.7 U/mL | feather powder | feather powder | feather powder | 72 | [27] |
| Bacillus licheniformis | OFAT | 50 | 11 U/mL | feather meal | feather meal | feather meal | 32 | [20] |
| B. subtilis BLBC17 | CCD | 33 | 170 U/mL | soybean meal | soybean meal | yeast extract | 48 | [18] |
| Bacillus licheniformis ER-15: | OFAT PB CCD | 70 | 1962 U/mL | feather | glucose | soy flour | 48 | [43] |
| Bacillus subtilis S14 | OFAT CCD | 50 | 5.5 U/mL | feather meal | feather meal | feather meal | 24 | [23] |
| Bacillus cereus Wu2 | OFAT | 30 | 1750 U/mL | chicken feather powder | chicken feather powder | nh4cl | 96 | [19] |
| Bacillus weihenstephanensis | OFAT | 40 | 15.3 U/mL | chicken feather | cellulose | (NH4)2 SO4 | 168 | [21] |
| Bacillus pumilus FH9 | OFAT | 37 | 647 U/mL | chicken feather | chicken feather | NH4CL yeast extract | 48 | [15] |
| Bacillus sp. 5 MG-MASC-BT | BBD | 55 | 1075 U/mL | alkali-treated soluble keratin | alkali-treated soluble keratin | alkali-treated soluble keratin | 60 | [24] |
| Bacillus licheniformis RPk | OFAT | 60 | 37.35 U/mL | chicken feather | chicken feather | yeast extract | - | [44] |
| B. subtilis 1273 | - | - | 412 U/mL | feather meal | feather meal | feather meal | 168 | [45] |
| Bacillus sp. UPM-AAG1 | PB CCD | 30 | 60.1 U/mL | chicken feather | chicken feather | chicken feather | 24 | Current study |
| Factors | Independent Factor | Unit | Range Level | |
|---|---|---|---|---|
| Minimum (−1) | Maximum (+1) | |||
| X1 | Temperature | (°C) | 25 | 35 |
| X2 | Inoculum | % (v/v) | 1 | 5 |
| X3 | pH | - | 5 | 8 |
| X4 | Feather (w/v) | % (w/v) | 1 | 5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdul Gafar, A.; Khayat, M.E.; Ahmad, S.A.; Yasid, N.A.; Shukor, M.Y. Response Surface Methodology for the Optimization of Keratinase Production in Culture Medium Containing Feathers by Bacillus sp. UPM-AAG1. Catalysts 2020, 10, 848. https://doi.org/10.3390/catal10080848
Abdul Gafar A, Khayat ME, Ahmad SA, Yasid NA, Shukor MY. Response Surface Methodology for the Optimization of Keratinase Production in Culture Medium Containing Feathers by Bacillus sp. UPM-AAG1. Catalysts. 2020; 10(8):848. https://doi.org/10.3390/catal10080848
Chicago/Turabian StyleAbdul Gafar, Aa’ishah, Mohd Ezuan Khayat, Siti Aqlima Ahmad, Nur Adeela Yasid, and Mohd Yunus Shukor. 2020. "Response Surface Methodology for the Optimization of Keratinase Production in Culture Medium Containing Feathers by Bacillus sp. UPM-AAG1" Catalysts 10, no. 8: 848. https://doi.org/10.3390/catal10080848