
FOXP3 Isoforms Expression in Cervical Cancer: Evidence about the Cancer-Related Properties of FOXP3Δ2Δ7 in Keratinocytes
 and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. FOXP3Δ2Δ7 Open Reading Frame Cloning
2.3. Production of Lentiviral Particles
2.4. HaCaT Cell Transduction
2.5. Quantitative PCR
2.6. Proliferation Assay (xCELLigence)
2.7. Cell Division Tracking
2.8. Cell Migration Assay
2.9. RNAseq
2.10. Gene Set Enrichment Analysis
2.11. Statistics
3. Results
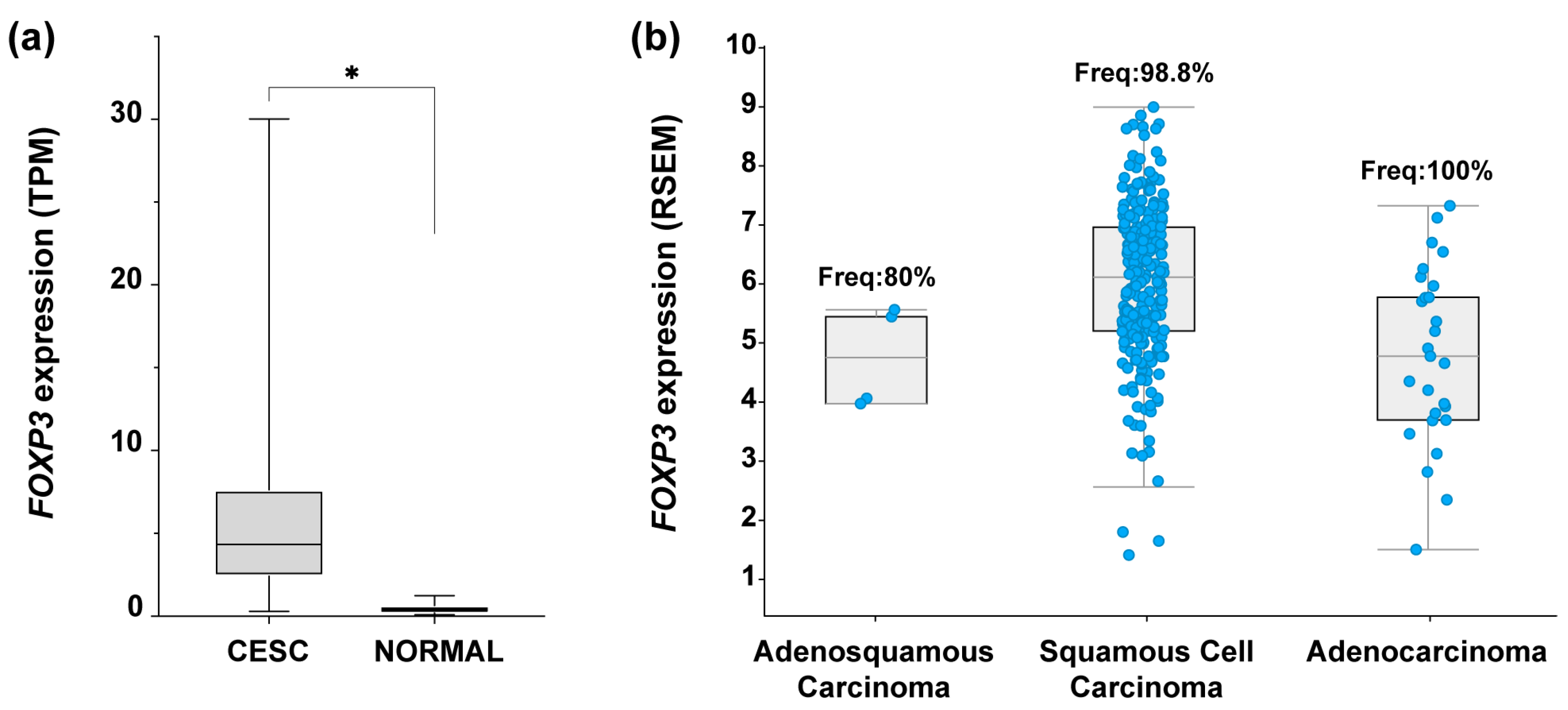
3.1. Augmented FOXP3 Expression in Cervical Cancer
3.2. FOXP3Δ2 Is the Most Prevalent Variant in Cervical Cancer Samples, and Its Expression Is Highly Differential in Metastatic Stages
3.3. HPV16 Infection Could Increase FOXP3 Expression Levels in Cervical Cancer
3.4. Exogenous Expression of FOXP3Δ2Δ7 Promotes Cell Proliferation, Division, and Migration
3.5. Exogenous Expression of FOXP3Δ2Δ7 Induces the Transcription of Protumoral Genes and the Enrichment of Immunological and Oncogenic Pathways
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buskwofie, A.; David-West, G.; Clare, C.A. A Review of Cervical Cancer: Incidence and Disparities. J. Natl. Med. Assoc. 2020, 112, 229–232. [Google Scholar] [CrossRef] [PubMed]
- Arbyn, M.; Weiderpass, E.; Bruni, L.; de Sanjosé, S.; Saraiya, M.; Ferlay, J.; Bray, F. Estimates of incidence and mortality of cervical cancer in 2018: A worldwide analysis. Lancet Glob. Health 2020, 8, e191–e203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prendiville, W.S.R. Colposcopy and Treatment of Cervical Precancer; World Health Organization: Geneva, Switzerland, 2017; Volume 45, p. 178. [Google Scholar]
- Zhou, J.; Wu, S.G.; Sun, J.Y.; Li, F.Y.; Lin, H.X.; Chen, Q.H.; He, Z.Y. Comparison of clinical outcomes of squamous cell carcinoma, adenocarcinoma, and adenosquamous carcinoma of the uterine cervix after definitive radiotherapy: A population-based analysis. J. Cancer Res. Clin. Oncol. 2017, 143, 115–122. [Google Scholar] [CrossRef]
- Lu, X.; Jiang, L.; Zhang, L.; Zhu, Y.; Hu, W.; Wang, J.; Ruan, X.; Xu, Z.; Meng, X.; Gao, J.; et al. Immune Signature-Based Subtypes of Cervical Squamous Cell Carcinoma Tightly Associated with Human Papillomavirus Type 16 Expression, Molecular Features, and Clinical Outcome. Neoplasia 2019, 21, 591–601. [Google Scholar] [CrossRef]
- Manini, I.; Montomoli, E. Epidemiology and prevention of Human Papillomavirus. Ann. Di Ig. Med. Prev. E Di Comunita 2018, 30, 28–32. [Google Scholar] [CrossRef]
- Chan, C.K.; Aimagambetova, G.; Ukybassova, T.; Kongrtay, K.; Azizan, A. Human Papillomavirus Infection and Cervical Cancer: Epidemiology, Screening, and Vaccination-Review of Current Perspectives. J. Oncol. 2019, 2019, 3257939. [Google Scholar] [CrossRef]
- Kaliff, M.; Sorbe, B.; Bohr Mordhorst, L.; Helenius, G.; Karlsson, M.G.; Lillsunde-Larsson, G. Findings of multiple HPV genotypes in cervical carcinoma are associated with poor cancer-specific survival in a Swedish cohort of cervical cancer primarily treated with radiotherapy. Oncotarget 2018, 9, 18786. [Google Scholar] [CrossRef] [Green Version]
- Vats, A.; Trejo-Cerro, O.; Thomas, M.; Banks, L. Human papillomavirus E6 and E7: What remains? Tumour Virus Res. 2021, 11, 200213. [Google Scholar] [CrossRef]
- Gutiérrez-Hoya, A.; Soto-Cruz, I. Role of the JAK/STAT Pathway in Cervical Cancer: Its Relationship with HPV E6/E7 Oncoproteins. Cells 2020, 9, 2297. [Google Scholar] [CrossRef]
- Cosper, P.F.; McNair, C.; González, I.; Wong, N.; Knudsen, K.E.; Chen, J.J.; Markovina, S.; Schwarz, J.K.; Grigsby, P.W.; Wang, X. Decreased local immune response and retained HPV gene expression during chemoradiotherapy are associated with treatment resistance and death from cervical cancer. Int. J. Cancer 2020, 146, 2047–2058. [Google Scholar] [CrossRef] [PubMed]
- Yeo-Teh, N.S.L.; Ito, Y.; Jha, S. High-Risk Human Papillomaviral Oncogenes E6 and E7 Target Key Cellular Pathways to Achieve Oncogenesis. Int. J. Mol. Sci. 2018, 19, 1706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mailer, R.K.W. Alternative Splicing of FOXP3-Virtue and Vice. Front. Immunol. 2018, 9, 530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, G.; Goodall, J.C.; Jarvis, L.B.; Hill Gaston, J.S. Characterisation of Foxp3 splice variants in human CD4+ and CD8+ T cells--identification of Foxp3Δ7 in human regulatory T cells. Mol. Immunol. 2010, 48, 321–332. [Google Scholar] [CrossRef]
- Williams, L.M.; Rudensky, A.Y. Maintenance of the Foxp3-dependent developmental program in mature regulatory T cells requires continued expression of Foxp3. Nat. Immunol. 2007, 8, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Deng, G.; Song, X.; Fujimoto, S.; Piccirillo, C.A.; Nagai, Y.; Greene, M.I. Foxp3 Post-translational Modifications and Treg Suppressive Activity. Front. Immunol. 2019, 10, 2486. [Google Scholar] [CrossRef] [Green Version]
- Fleskens, V.; Minutti, C.M.; Wu, X.; Wei, P.; Pals, C.E.G.M.; McCrae, J.; Hemmers, S.; Groenewold, V.; Vos, H.-J.; Rudensky, A.; et al. Nemo-like Kinase Drives Foxp3 Stability and Is Critical for Maintenance of Immune Tolerance by Regulatory T Cells. Cell Rep. 2019, 26, 3600–3612.e3606. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Huang, C.; Zhou, B.; Ziegler, S.F. Isoform-specific inhibition of ROR alpha-mediated transcriptional activation by human FOXP3. J. Immunol. 2008, 180, 4785–4792. [Google Scholar] [CrossRef] [Green Version]
- Ichiyama, K.; Yoshida, H.; Wakabayashi, Y.; Chinen, T.; Saeki, K.; Nakaya, M.; Takaesu, G.; Hori, S.; Yoshimura, A.; Kobayashi, T. Foxp3 inhibits RORgammat-mediated IL-17A mRNA transcription through direct interaction with RORgammat. J. Biol. Chem. 2008, 283, 17003–17008. [Google Scholar] [CrossRef] [Green Version]
- Sambucci, M.; Gargano, F.; De Rosa, V.; De Bardi, M.; Picozza, M.; Placido, R.; Ruggieri, S.; Capone, A.; Gasperini, C.; Matarese, G.; et al. FoxP3 isoforms and PD-1 expression by T regulatory cells in multiple sclerosis. Sci. Rep. 2018, 8, 3674. [Google Scholar] [CrossRef]
- Li, L.; Xu, X.T.; Wang, L.L.; Qin, S.B.; Zhou, J.Y. Expression and clinicopathological significance of Foxp3 and VISTA in cervical cancer. Am. J. Transl Res. 2021, 13, 10428–10438. [Google Scholar] [PubMed]
- Litwin, T.R.; Irvin, S.R.; Chornock, R.L.; Sahasrabuddhe, V.V.; Stanley, M.; Wentzensen, N. Infiltrating T-cell markers in cervical carcinogenesis: A systematic review and meta-analysis. Br. J. Cancer 2021, 124, 831–841. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.Y.; Chen, C.; Wang, L.; Chang, X.; Zheng, P.; Liu, Y. Cutting edge: Broad expression of the FoxP3 locus in epithelial cells: A caution against early interpretation of fatal inflammatory diseases following in vivo depletion of FoxP3-expressing cells. J. Immunol. 2008, 180, 5163–5166. [Google Scholar] [CrossRef] [Green Version]
- Alfaar, A.S.; Stürzbecher, L.; Diedrichs-Möhring, M.; Lam, M.; Roubeix, C.; Ritter, J.; Schumann, K.; Annamalai, B.; Pompös, I.-M.; Rohrer, B.; et al. FoxP3 expression by retinal pigment epithelial cells: Transcription factor with potential relevance for the pathology of age-related macular degeneration. J. Neuroinflammation 2022, 19, 260. [Google Scholar] [CrossRef]
- Karanikas, V.; Speletas, M.; Zamanakou, M.; Kalala, F.; Loules, G.; Kerenidi, T.; Barda, A.K.; Gourgoulianis, K.I.; Germenis, A.E. Foxp3 expression in human cancer cells. J. Transl. Med. 2008, 6, 19. [Google Scholar] [CrossRef] [Green Version]
- Jia, H.; Qi, H.; Gong, Z.; Yang, S.; Ren, J.; Liu, Y.; Li, M.-Y.; Chen, G.G. The expression of FOXP3 and its role in human cancers. Biochim. Et Biophys. Acta (BBA) Rev. Cancer 2019, 1871, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Zuo, T.; Liu, R.; Zhang, H.; Chang, X.; Liu, Y.; Wang, L.; Zheng, P.; Liu, Y. FOXP3 is a novel transcriptional repressor for the breast cancer oncogene SKP2. J. Clin. Investig. 2007, 117, 3765–3773. [Google Scholar] [CrossRef] [Green Version]
- Zuo, T.; Wang, L.; Morrison, C.; Chang, X.; Zhang, H.; Li, W.; Liu, Y.; Wang, Y.; Liu, X.; Chan, M.W.Y.; et al. FOXP3 is an X-linked breast cancer suppressor gene and an important repressor of the HER-2/ErbB2 oncogene. Cell 2007, 129, 1275–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douglass, S.; Ali, S.; Meeson, A.P.; Browell, D.; Kirby, J.A. The role of FOXP3 in the development and metastatic spread of breast cancer. Cancer Metastasis Rev. 2012, 31, 843–854. [Google Scholar] [CrossRef]
- Wang, L.; Liu, R.; Li, W.; Chen, C.; Katoh, H.; Chen, G.-Y.; McNally, B.; Lin, L.; Zhou, P.; Zuo, T.; et al. Somatic single hits inactivate the X-linked tumor suppressor FOXP3 in the prostate. Cancer Cell 2009, 16, 336–346. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Sun, H. Up-regulation of Foxp3 inhibits cell proliferation, migration and invasion in epithelial ovarian cancer. Cancer Lett. 2010, 287, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Chu, R.; Liu, S.Y.W.; Vlantis, A.C.; van Hasselt, C.A.; Ng, E.K.W.; Fan, M.D.; Ng, S.K.; Chan, A.B.W.; Du, J.; Wei, W.; et al. Inhibition of Foxp3 in cancer cells induces apoptosis of thyroid cancer cells. Mol. Cell. Endocrinol. 2015, 399, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Hinz, S.; Pagerols-Raluy, L.; Oberg, H.H.; Ammerpohl, O.; Grussel, S.; Sipos, B.; Grutzmann, R.; Pilarsky, C.; Ungefroren, H.; Saeger, H.D.; et al. Foxp3 expression in pancreatic carcinoma cells as a novel mechanism of immune evasion in cancer. Cancer Res. 2007, 67, 8344–8350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.; Grimmig, T.; Grimm, M.; Lazariotou, M.; Meier, E.; Rosenwald, A.; Tsaur, I.; Blaheta, R.; Heemann, U.; Germer, C.T.; et al. Expression of Foxp3 in colorectal cancer but not in Treg cells correlates with disease progression in patients with colorectal cancer. PLoS ONE 2013, 8, e53630. [Google Scholar] [CrossRef] [Green Version]
- Jou, Y.C.; Tsai, Y.S.; Lin, C.T.; Tung, C.L.; Shen, C.H.; Tsai, H.T.; Yang, W.H.; Chang, H.I.; Chen, S.Y.; Tzai, T.S. Foxp3 enhances HIF-1α target gene expression in human bladder cancer through decreasing its ubiquitin-proteasomal degradation. Oncotarget 2016, 7, 65403–65417. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Xu, J.; Zhang, X.; Zhang, Y.; Wang, L.; Huang, X.; Xu, Z. The Role of Tumoral FOXP3 on Cell Proliferation, Migration, and Invasion in Gastric Cancer. Cell. Physiol. Biochem. 2017, 42, 1739–1754. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Qi, H.; Peng, J.; Li, M.; Chen, G.G. FOXP3Δ3, the major isoform of FOXP3, promotes proliferation, migration, and invasion in non-small cell lung cancer. Ann. Oncol. 2019, 30, ii5–ii6. [Google Scholar] [CrossRef]
- Zeng, C.; Yao, Y.; Jie, W.; Zhang, M.; Hu, X.; Zhao, Y.; Wang, S.; Yin, J.; Song, Y. Up-regulation of Foxp3 participates in progression of cervical cancer. Cancer Immunol. Immunother. CII 2012, 62, 481–487. [Google Scholar] [CrossRef]
- Luo, Q.; Zhang, S.; Wei, H.; Pang, X.; Zhang, H. Roles of Foxp3 in the occurrence and development of cervical cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 8717–8730. [Google Scholar]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [Green Version]
- Okonechnikov, K.; Golosova, O.; Fursov, M. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef] [PubMed]
- Team, R. RStudio: Integrated Development for R; 1.4.1717; RStudio, Inc.: Boston, MA, USA, 2015. [Google Scholar]
- Afgan, E.; Baker, D.; Batut, B.; van den Beek, M.; Bouvier, D.; Čech, M.; Chilton, J.; Clements, D.; Coraor, N.; Grüning, B.A.; et al. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2018 update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, S. FastQC A Quality Control tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 3 February 2022).
- Liao, Y.; Smyth, G.K.; Shi, W. The R package Rsubread is easier, faster, cheaper and better for alignment and quantification of RNA sequencing reads. Nucleic Acids Res. 2019, 47, e47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2013, 30, 923–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Tang, G.; Cho, M.; Wang, X. OncoDB: An interactive online database for analysis of gene expression and viral infection in cancer. Nucleic Acids Res. 2022, 50, D1334–D1339. [Google Scholar] [CrossRef]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal 2013, 6, pl1. [Google Scholar] [CrossRef] [Green Version]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio Cancer Genomics Portal: An Open Platform for Exploring Multidimensional Cancer Genomics Data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [Green Version]
- Yang, I.S.; Son, H.; Kim, S.; Kim, S. ISOexpresso: A web-based platform for isoform-level expression analysis in human cancer. BMC Genom. 2016, 17, 631. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.; Duan, T.; Ye, P.; Chen, K.; Zhang, G.; Lai, M.; Zhang, H. TSVdb: A web-tool for TCGA splicing variants analysis. BMC Genom. 2018, 19, 405. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Josefowicz, S.Z.; Kas, A.; Chu, T.T.; Gavin, M.A.; Rudensky, A.Y. Genome-wide analysis of Foxp3 target genes in developing and mature regulatory T cells. Nature 2007, 445, 936–940. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Yi, B.; Wei, S.; Rao, D.; He, Y.; Naik, G.; Bae, S.; Liu, X.M.; Yang, W.H.; Sonpavde, G.; et al. Loss of FOXP3 and TSC1 Accelerates Prostate Cancer Progression through Synergistic Transcriptional and Posttranslational Regulation of c-MYC. Cancer Res. 2019, 79, 1413–1425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Prado, K.; Zhang, K.X.; Peek, E.M.; Lee, J.; Wang, X.; Huang, J.; Li, G.; Pellegrini, M.; Chin, A.I. Biased Expression of the FOXP3Δ3 Isoform in Aggressive Bladder Cancer Mediates Differentiation and Cisplatin Chemotherapy Resistance. Clin Cancer Res. 2016, 22, 5349–5361. [Google Scholar] [CrossRef] [Green Version]
- Lopes, J.E.; Torgerson, T.R.; Schubert, L.A.; Anover, S.D.; Ocheltree, E.L.; Ochs, H.D.; Ziegler, S.F. Analysis of FOXP3 Reveals Multiple Domains Required for Its Function as a Transcriptional Repressor. J. Immunol. 2006, 177, 3133–3142. [Google Scholar] [CrossRef] [Green Version]
- Chae, W.J.; Henegariu, O.; Lee, S.K.; Bothwell, A.L. The mutant leucine-zipper domain impairs both dimerization and suppressive function of Foxp3 in T cells. Proc. Natl. Acad. Sci. USA 2006, 103, 9631–9636. [Google Scholar] [CrossRef] [Green Version]
- van der Vliet, H.J.; Nieuwenhuis, E.E. IPEX as a result of mutations in FOXP3. Clin. Dev. Immunol. 2007, 2007, 89017. [Google Scholar] [CrossRef] [Green Version]
- Magg, T.; Wiebking, V.; Conca, R.; Krebs, S.; Arens, S.; Schmid, I.; Klein, C.; Albert, M.H.; Hauck, F. IPEX due to an exon 7 skipping FOXP3 mutation with autoimmune diabetes mellitus cured by selective TReg cell engraftment. Clin. Immunol. 2018, 191, 52–58. [Google Scholar] [CrossRef]
- Mailer, R.K.; Falk, K.; Rotzschke, O. Absence of leucine zipper in the natural FOXP3Delta2Delta7 isoform does not affect dimerization but abrogates suppressive capacity. PLoS ONE 2009, 4, e6104. [Google Scholar] [CrossRef] [Green Version]
- Lv, J.H.; Wang, F.; Shen, M.H.; Wang, X.; Zhou, X.J. SATB1 expression is correlated with β-catenin associated epithelial-mesenchymal transition in colorectal cancer. Cancer Biol. Ther. 2016, 17, 254–261. [Google Scholar] [CrossRef] [Green Version]
- Mao, L.J.; Yang, C.H.; Fan, L.; Gao, P.; Yang, D.R.; Xue, B.X.; Zheng, J.N.; Shan, Y.X. SATB1 promotes prostate cancer metastasis by the regulation of epithelial-mesenchymal transition. Biomed Pharm. 2016, 79, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Glatzel-Plucinska, N.; Piotrowska, A.; Rzechonek, A.; Podhorska-Okolow, M.; Dziegiel, P. SATB1 protein is associated with the epithelial-mesenchymal transition process in non-small cell lung cancers. Oncol. Rep. 2021, 45, 118. [Google Scholar] [CrossRef] [PubMed]
- Huo, F.-C.; Pan, Y.-J.; Li, T.-T.; Mou, J.; Pei, D.-S. PAK5 promotes the migration and invasion of cervical cancer cells by phosphorylating SATB1. Cell Death Differ. 2019, 26, 994–1006. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zeng, J.; Xiao, R.; Xu, G.; Liu, G.; Xiong, D.; Ye, Y.; Chen, B.; Wang, H.; Luo, Q.; et al. Poor prognosis and SATB1 overexpression in solid tumors: A meta-analysis. Cancer Manag. Res. 2018, 10, 1471–1478. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Zheng, Y.; Ji, Y.; Zhang, X. The expression of special AT-rich binding protein 1 in cervical cancer and its clinical significance. Onco. Targets Ther. 2019, 12, 945–951. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Bai, C.B.; Joyner, A.L.; Wang, B. Sonic hedgehog signaling regulates Gli2 transcriptional activity by suppressing its processing and degradation. Mol. Cell Biol. 2006, 26, 3365–3377. [Google Scholar] [CrossRef] [Green Version]
- Ichimiya, S.; Onishi, H.; Nagao, S.; Koga, S.; Sakihama, K.; Nakayama, K.; Fujimura, A.; Oyama, Y.; Imaizumi, A.; Oda, Y.; et al. GLI2 but not GLI1/GLI3 plays a central role in the induction of malignant phenotype of gallbladder cancer. Oncol Rep. 2021, 45, 997–1010. [Google Scholar] [CrossRef]
- Lu, Y.; Zhang, B.; Wang, B.; Wu, D.; Wang, C.; Gao, Y.; Liang, W.; Xi, H.; Wang, X.; Chen, L. MiR-144-3p inhibits gastric cancer progression and stemness via directly targeting GLI2 involved in hedgehog pathway. J. Transl. Med. 2021, 19, 432. [Google Scholar] [CrossRef]
- Zhu, H.; Xia, L.; Shen, Q.; Zhao, M.; Gu, X.; Bouamar, H.; Wang, B.; Sun, L.Z.; Zhu, X. Differential effects of GLI2 and GLI3 in regulating cervical cancer malignancy in vitro and in vivo. Lab. Investig. 2018, 98, 1384–1396. [Google Scholar] [CrossRef] [Green Version]
- Riihilä, P.; Viiklepp, K.; Nissinen, L.; Farshchian, M.; Kallajoki, M.; Kivisaari, A.; Meri, S.; Peltonen, J.; Peltonen, S.; Kähäri, V.M. Tumour-cell-derived complement components C1r and C1s promote growth of cutaneous squamous cell carcinoma. Br. J. Dermatol. 2020, 182, 658–670. [Google Scholar] [CrossRef] [Green Version]
- Viiklepp, K.; Nissinen, L.; Ojalill, M.; Riihilä, P.; Kallajoki, M.; Meri, S.; Heino, J.; Kähäri, V.M. C1r Upregulates Production of Matrix Metalloproteinase-13 and Promotes Invasion of Cutaneous Squamous Cell Carcinoma. J. Invest. Dermatol. 2022, 142, 1478–1488.e1479. [Google Scholar] [CrossRef] [PubMed]
- Gürz, S.; Çelik, B.; Menteşe, A.; Us Altay, D. Diagnostic value of signal peptide-Complement C1r/C1s, Uegf, and Bmp1-epidermal growth factor domain-containing protein 1 on serum and tissue samples in non-small cell lung cancer. Turk. Gogus Kalp. Damar. Cerrahisi Derg. 2018, 26, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Kanao, H.; Enomoto, T.; Kimura, T.; Fujita, M.; Nakashima, R.; Ueda, Y.; Ueno, Y.; Miyatake, T.; Yoshizaki, T.; Buzard, G.S.; et al. Overexpression of LAMP3/TSC403/DC-LAMP promotes metastasis in uterine cervical cancer. Cancer Res. 2005, 65, 8640–8645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bettelli, E.; Dastrange, M.; Oukka, M. Foxp3 interacts with nuclear factor of activated T cells and NF-kappa B to repress cytokine gene expression and effector functions of T helper cells. Proc. Natl. Acad. Sci. USA 2005, 102, 5138–5143. [Google Scholar] [CrossRef] [Green Version]
- Niu, J.; Jiang, C.; Li, C.; Liu, L.; Li, K.; Jian, Z.; Gao, T. Foxp3 expression in melanoma cells as a possible mechanism of resistance to immune destruction. Cancer Immunol. Immunother. 2011, 60, 1109–1118. [Google Scholar] [CrossRef]
- Downward, J. Role of receptor tyrosine kinases in G-protein-coupled receptor regulation of Ras: Transactivation or parallel pathways? Biochem. J. 2003, 376, e9–e10. [Google Scholar] [CrossRef] [Green Version]
- Perera, D.; Venkitaraman, A.R. Oncogenic KRAS triggers MAPK-dependent errors in mitosis and MYC-dependent sensitivity to anti-mitotic agents. Sci. Rep. 2016, 6, 29741. [Google Scholar] [CrossRef] [Green Version]
- Hamarsheh, S.a.; Groß, O.; Brummer, T.; Zeiser, R. Immune modulatory effects of oncogenic KRAS in cancer. Nat. Commun. 2020, 11, 5439. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, W.; Na, F.; Li, M.; Tong, S. LINC01354/microRNA-216b/KRAS Axis Promotes the Occurrence and Metastasis of Endometrial Cancer. Nanoscale Res. Lett. 2022, 17, 21. [Google Scholar] [CrossRef]
- Liao, W.; Overman, M.J.; Boutin, A.T.; Shang, X.; Zhao, D.; Dey, P.; Li, J.; Wang, G.; Lan, Z.; Li, J.; et al. KRAS-IRF2 Axis Drives Immune Suppression and Immune Therapy Resistance in Colorectal Cancer. Cancer Cell 2019, 35, 559–572.e557. [Google Scholar] [CrossRef] [Green Version]
- Tsujino, T.; Sugito, N.; Taniguchi, K.; Honda, R.; Komura, K.; Yoshikawa, Y.; Takai, T.; Minami, K.; Kuranaga, Y.; Shinohara, H.; et al. MicroRNA-143/Musashi-2/KRAS cascade contributes positively to carcinogenesis in human bladder cancer. Cancer Sci. 2019, 110, 2189–2199. [Google Scholar] [CrossRef] [PubMed]
- Tang, A.H.; Hoefer, R.A.; Guye, M.L.; Bear, H.D. Persistent EGFR/K-RAS/SIAH pathway activation drives chemo-resistance and early tumor relapse in triple-negative breast cancer. Cancer Drug Resist. 2022, 5, 691–702. [Google Scholar] [CrossRef] [PubMed]
- Wegman, P.; Ahlin, C.; Sorbe, B. Genetic Alterations in the K-Ras Gene Influence the Prognosis in Patients With Cervical Cancer Treated by Radiotherapy. Int. J. Gynecol. Cancer 2011, 21, 86. [Google Scholar] [CrossRef]
- Ramos-Solano, M.; Meza-Canales, I.D.; Torres-Reyes, L.A.; Alvarez-Zavala, M.; Alvarado-Ruíz, L.; Rincon-Orozco, B.; Garcia-Chagollan, M.; Ochoa-Hernández, A.B.; Ortiz-Lazareno, P.C.; Rösl, F.; et al. Expression of WNT genes in cervical cancer-derived cells: Implication of WNT7A in cell proliferation and migration. Exp. Cell Res. 2015, 335, 39–50. [Google Scholar] [CrossRef]
- Mir, R.; Pradhan, S.J.; Patil, P.; Mulherkar, R.; Galande, S. Wnt/β-catenin signaling regulated SATB1 promotes colorectal cancer tumorigenesis and progression. Oncogene 2016, 35, 1679–1691. [Google Scholar] [CrossRef]
- Ruan, K.; Song, G.; Ouyang, G. Role of hypoxia in the hallmarks of human cancer. J. Cell Biochem. 2009, 107, 1053–1062. [Google Scholar] [CrossRef] [PubMed]
- Jing, X.; Yang, F.; Shao, C.; Wei, K.; Xie, M.; Shen, H.; Shu, Y. Role of hypoxia in cancer therapy by regulating the tumor microenvironment. Mol. Cancer 2019, 18, 157. [Google Scholar] [CrossRef] [Green Version]
- Emami Nejad, A.; Najafgholian, S.; Rostami, A.; Sistani, A.; Shojaeifar, S.; Esparvarinha, M.; Nedaeinia, R.; Haghjooy Javanmard, S.; Taherian, M.; Ahmadlou, M.; et al. The role of hypoxia in the tumor microenvironment and development of cancer stem cell: A novel approach to developing treatment. Cancer Cell Int. 2021, 21, 62. [Google Scholar] [CrossRef]
- Yan, B.; Ma, Q.F.; Tan, W.F.; Cai, H.N.; Li, Y.L.; Zhou, Z.G.; Dai, X.; Zhu, F.X.; Xiong, Y.J.; Xu, M.; et al. Expression of HIF-1α is a predictive marker of the efficacy of neoadjuvant chemotherapy for locally advanced cervical cancer. Oncol. Lett. 2020, 20, 841–849. [Google Scholar] [CrossRef]
- Datta, A.; West, C.; O’Connor, J.P.B.; Choudhury, A.; Hoskin, P. Impact of hypoxia on cervical cancer outcomes. Int. J. Gynecol. Cancer 2021, 31, 1459–1470. [Google Scholar] [CrossRef]
- Dehdashti, F.; Grigsby, P.W.; Lewis, J.S.; Laforest, R.; Siegel, B.A.; Welch, M.J. Assessing tumor hypoxia in cervical cancer by PET with 60Cu-labeled diacetyl-bis(N4-methylthiosemicarbazone). J. Nucl. Med. 2008, 49, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Mujcic, H.; Nagelkerke, A.; Rouschop, K.M.; Chung, S.; Chaudary, N.; Span, P.N.; Clarke, B.; Milosevic, M.; Sykes, J.; Hill, R.P.; et al. Hypoxic activation of the PERK/eIF2α arm of the unfolded protein response promotes metastasis through induction of LAMP3. Clin. Cancer Res. 2013, 19, 6126–6137. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse | Amplicon Size |
|---|---|---|---|
| FOXP31 | 5′ ACA AGC CAG GCT GAT CCT T 3′ | 5′ CAC ATC CAG GGC CTA TCA TC 3′ | 1347 bp |
| FOXP32 | 5′ CAA GTT CCA CAA CAT GCG ACC 3′ | 5′ GCT CTC CAC CCG CAC AAA 3′ | 208 bp |
| SATB12 | 5′ CCT CAG CCA GAA CGT GAT GC 3′ | 5′ GAC TCT GCT GGA GAG GCC A 3′ | 236 bp |
| C1R2 | 5′ AAG ATT CCT CGG TGC TTG CC 3′ | 5′ GTT GCT TTG CGC TTC GTG TT 3′ | 216 bp |
| GLI22 | 5′ CAA CAA TGA CAG TGG CGT GG 3′ | 5′ CTG CCA CTG AAG TTT TCC AGG 3′ | 297 bp |
| LAMP32 | 5′ ACA TGC GGT GGT GAT GTT CC 3′ | 5′ AGG CAG AGA CCA ACC ACG AT 3′ | 219 bp |
| NSG12 | 5′ TTC CTC ACC TGC GTC GTC TT 3′ | 5′ AAC TTG CCC ATC CCG CTA AG 3′ | 297 bp |
| HSPB82 | 5′ GGT GGC ATT GTT TCT AAG A 3′ | 5′ TAC TGG CAT CTC AGG TAC AG 3′ | 208 bp |
| RPLP02 | 5′ CCT CAT ATC CGG GGG AAT GTG 3′ | 5′ GCA GCA GCT GGC ACC TTA TTG 3′ | 95 pb |
| RPL322 | 5′ GCA TTG ACA ACA GGG TTC GTA G 3′ | 5′ ATT TAA ACA GAA AAC GTG CAC A 3′ | 320 pb |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia-Becerra, N.; Aguila-Estrada, M.U.; Palafox-Mariscal, L.A.; Hernandez-Flores, G.; Aguilar-Lemarroy, A.; Jave-Suarez, L.F. FOXP3 Isoforms Expression in Cervical Cancer: Evidence about the Cancer-Related Properties of FOXP3Δ2Δ7 in Keratinocytes. Cancers 2023, 15, 347. https://doi.org/10.3390/cancers15020347
Garcia-Becerra N, Aguila-Estrada MU, Palafox-Mariscal LA, Hernandez-Flores G, Aguilar-Lemarroy A, Jave-Suarez LF. FOXP3 Isoforms Expression in Cervical Cancer: Evidence about the Cancer-Related Properties of FOXP3Δ2Δ7 in Keratinocytes. Cancers. 2023; 15(2):347. https://doi.org/10.3390/cancers15020347
Chicago/Turabian StyleGarcia-Becerra, Natalia, Marco Ulises Aguila-Estrada, Luis Arturo Palafox-Mariscal, Georgina Hernandez-Flores, Adriana Aguilar-Lemarroy, and Luis Felipe Jave-Suarez. 2023. "FOXP3 Isoforms Expression in Cervical Cancer: Evidence about the Cancer-Related Properties of FOXP3Δ2Δ7 in Keratinocytes" Cancers 15, no. 2: 347. https://doi.org/10.3390/cancers15020347