
Malignancy Associated with Low-Risk HPV6 and HPV11: A Systematic Review and Implications for Cancer Prevention
 , , , , , ,
, , , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
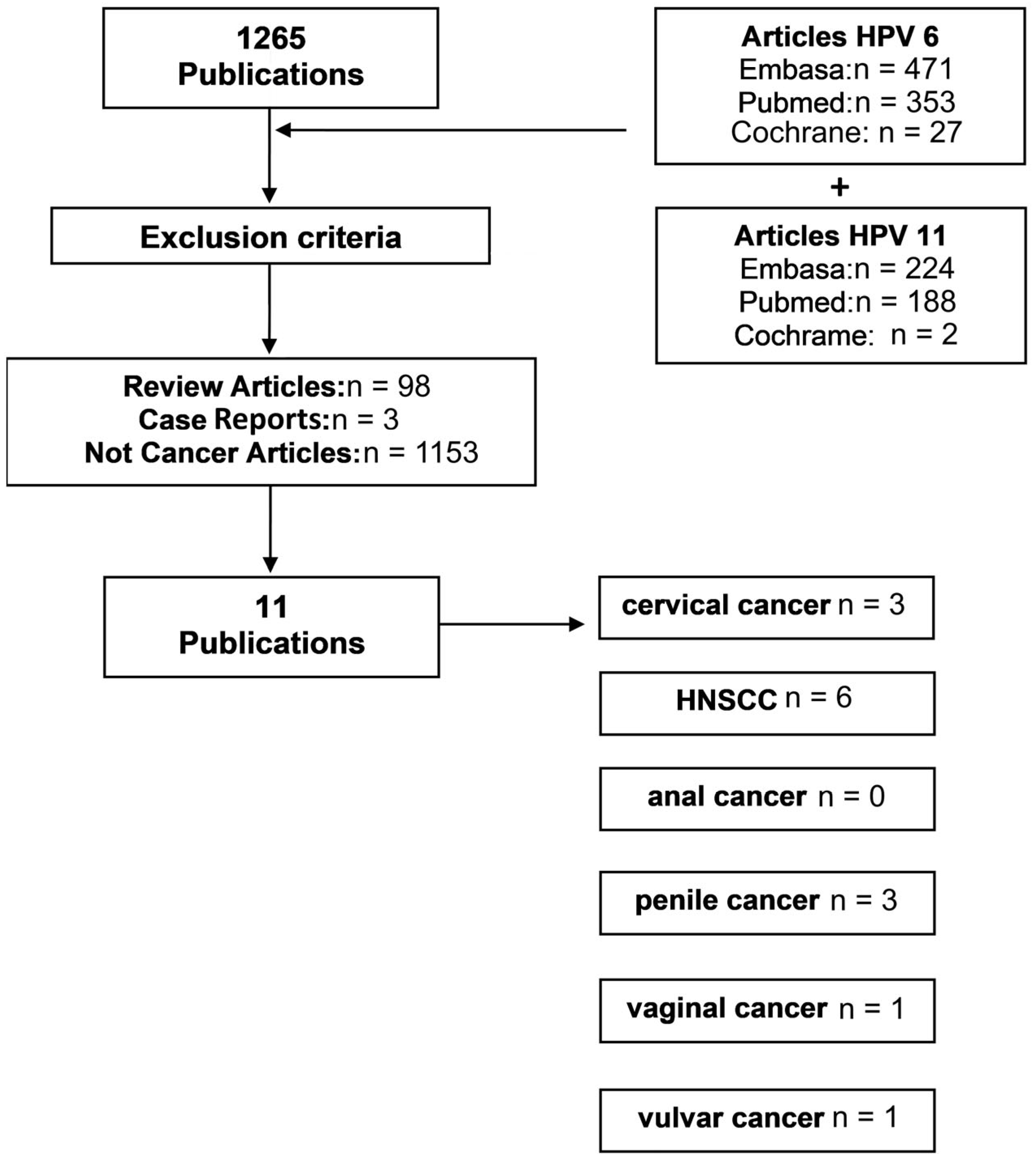
2.1. Systematic Review of HPV6 and HPV11 in Cancer
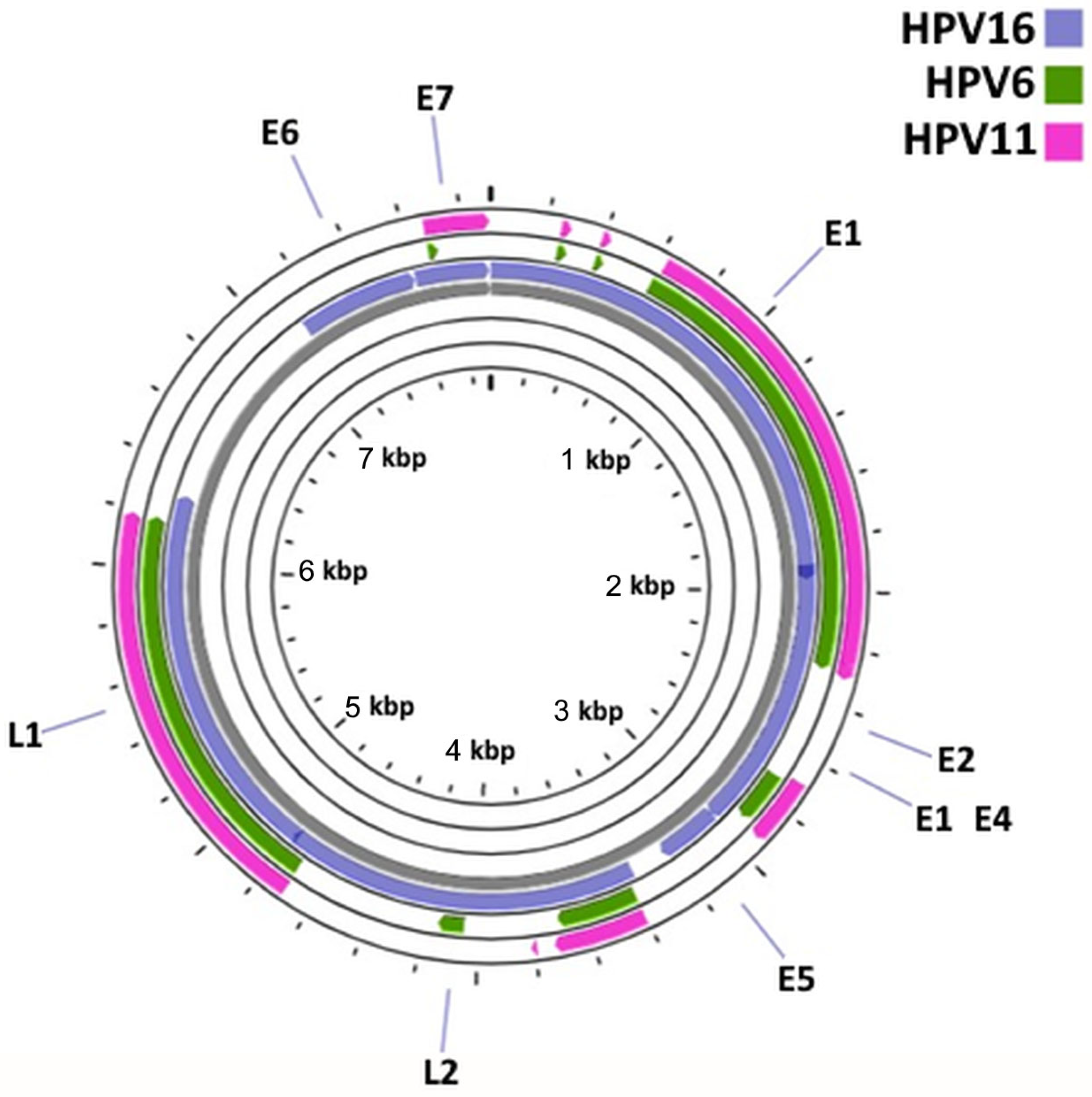
2.2. Comparative Genomic Analysis of HPV6, HPV11 and HPV16
3. Results
3.1. Systematic Review of HPV6 and HPV11 in Cancer
3.2. HPV6/11/16 Comparative Genomic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tommasino, M. The human papillomavirus family and its role in carcinogenesis. Semin. Cancer Biol. 2014, 26, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Vashisht, S.; Mishra, H.; Mishra, P.K.; Ekielski, A.; Talegaonkar, S. Structure, genome, infection cycle and clinical manifestations associated with human papillomavirus. Curr. Pharm. Biotechnol. 2019, 20, 1260–1280. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Baleja, J. Structure and function of the papillomavirus E6 protein and its interacting proteins. Front. Biosci. 2008, 1, 121–134. [Google Scholar] [CrossRef] [Green Version]
- De Sanjose, S.; Brotons, M.; Pavon, M.A. The natural history of human papillomavirus infection. Best Pract. Res. Clin. Obstet. Gynaecol. 2018, 47, 2–13. [Google Scholar] [CrossRef]
- Estêvão, D.; Costa, N.R.; da Costa RM, G.; Medeiros, R. Hallmarks of HPV carcinogenesis: The role of E6, E7 and E5 oncoproteins in cellular malignancy. Biochim. Biophys. Acta–Gene Regul. Mech. 2019, 1862, 153–162. [Google Scholar] [CrossRef]
- Zur Hausen, H. Papillomaviruses and cancer: From basic studies to clinical application. Nat. Rev. Cancer 2002, 2, 342–350. [Google Scholar] [CrossRef]
- Zur Hausen, H. Papillomavirus infections: A major cause of human cancers. In Infections Causing Human Cancer; Zur Hausen, H., Ed.; Wiley–VCH: Weinheim, Germany, 2006; pp. 145–243. [Google Scholar]
- Bernard, H.U.; Burk, R.D.; Chen, Z.; van Doorslaer, K.; Zur Hausen, H.; de Villiers, E.M. Classification of papillomaviruses (PVs) based on 189 PV types and proposal of taxonomic amendments. Virology 2010, 401, 70–79. [Google Scholar] [CrossRef] [Green Version]
- De Villiers, E.M.; Fauquet, C.; Broker, T.R.; Bernard, H.U.; Zur Hausen, H. Classification of papillomaviruses. Virology 2004, 324, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, N.; Bosch, F.X.; De Sanjosé, S.; Herrero, R.; Castellsagué, X.; Shah, K.V.; Snijders, P.J.F.; Meijer, C.J.L.M. Epidemiologic classifcation of human papillomavirus types associated with cervical cancer. N. Engl. J. Med. 2003, 348, 518–527. [Google Scholar] [CrossRef] [Green Version]
- Salgado, A.H.; Martín-Gámez, D.C.; Moreno, P.; Murillo, R.; Bravo, M.M.; Villa, L.; Molano, M. E6 molecular variants of human papillomavirus (HPV) type 16: An updated and unified criterion for clustering and nomenclature. Virology 2011, 410, 201–215. [Google Scholar] [CrossRef] [Green Version]
- Sichero, L.; Sobrinho, J.S.; Villa, L.L. Oncogenic potential diverge among human papillomavirus type 16 natural variants. Virology 2012, 432, 127–132. [Google Scholar] [CrossRef]
- Cochicho, D.; da Costa, R.G.; Felix, A. Exploring the roles of HPV16 variants in head and neck squamous cell carcinoma: Current challenges and opportunities. Virol. J. 2021, 18, 217. [Google Scholar] [CrossRef]
- Leto, M.D.G.P.; Santos Júnior, G.F.D.; Porro, A.M.; Tomimori, J. Human papillomavirus infection: Etiopathogenesis, molecular biology and clinical manifestations. An. Bras. Dermatol. 2011, 86, 306–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kroupis, C.; Vourlidis, N. Human papilloma virus (HPV) molecular diagnostics. Clin. Chem. Lab. Med. 2011, 49, 1783–1799. [Google Scholar] [CrossRef] [PubMed]
- Hampson, I.N. Effects of the prophylactic HPV vaccines on HPV type prevalence and cervical pathology. Viruses 2022, 14, 757. [Google Scholar] [CrossRef] [PubMed]
- Handisurya, A.; Schellenbacher, C.; Kirnbauer, R. Diseases caused by human papillomaviruses (HPV). J. Dtsch. Dermatol. Ges. 2009, 7, 453–666, quiz 466, 467. [Google Scholar] [CrossRef] [PubMed]
- Forman, D.; de Martel, C.; Lacey, C.J.; Soerjomataram, I.; Lortet-Tieulent, J.; Bruni, L.; Vignat, J.; Ferlay, J.; Bray, F.; Plummer, M.; et al. Global burden of human papillomavirus and related diseases. Vaccine 2012, 30, F12–F23. [Google Scholar] [CrossRef] [Green Version]
- Yuan, H.; Zhou, D.; Wang, J.; Schlegel, R. Divergent human papillomavirus associated with recurrent respiratory papillomatosis with lung involvement. Genome Announc. 2013, 1, 10. [Google Scholar] [CrossRef] [Green Version]
- Donne, A.; Hampson, L.; Homer, J.; Hampson, I. The role of HPV type in recurrent respiratory papillomatosis. Int. J. Pediatr. Otorhinolaryngol. 2010, 74, 7–14. [Google Scholar] [CrossRef]
- Faber, M.T.; Sand, F.L.; Albieri, V.; Norrild, B.; Kjær, S.K.; Verdoodt, F. Prevalence and type distribution of human papillomavirus in squamous cell carcinoma and intraepithelial neoplasia of the vulva. Int. J. Cancer 2017, 141, 1161–1169. [Google Scholar] [CrossRef] [Green Version]
- Olesen, T.B.; Sand, F.L.; Rasmussen, C.L.; Albieri, V.; Toft, B.G.; Norrild, B.; Munk, C.; Kjær, S.K. Prevalence of human papillomavirus DNA and p16INK4a in penile cancer and penile intraepithelial neoplasia: A systematic review and meta-analysis. Lancet Oncol. 2019, 20, 145–158. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. Int. J. Surg. 2021, 88, 105906. [Google Scholar] [CrossRef]
- Grant, J.R.; Stothard, P. The CGView server: A comparative genomics tool for circular genomes. Nucleic Acids Res. 2008, 36, W181–W184. [Google Scholar] [CrossRef] [PubMed]
- Tao, G.; Yaling, G.; Zhan, G.; Pu, L.; Miao, H. Human papillomavirus genotype distribution among HPV-positive women in Sichuan province, Southwest China. Arch. Virol. 2017, 163, 65–72. [Google Scholar] [CrossRef]
- Das, D.; Rai, A.K.; Kataki, A.C.; Barmon, D.; Deka, P.; Sharma, J.D.; Sarma, A.; Shrivastava, S.; Bhattacharyya, M.; Kalita, A.K.; et al. Nested multiplex PCR based detection of human papillomavirus in cervical carcinoma patients of North-East India. Asian Pac. J. Cancer Prev. 2013, 14, 785–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aldersley, J.; Lorenz, D.R.; Mouw, K.W.; D’Andrea, A.D.; Gabuzda, D. Genomic landscape of primary and recurrent anal squamous cell carcinomas in relation to HPV integration, copy-number variation, and DNA damage response genes. Mol. Cancer Res. 2021, 19, 1308–1321. [Google Scholar] [CrossRef]
- Magaña-León, C.; Oros, C.; López-Revilla, R. Human papillomavirus types in non-cervical high-grade intraepithelial neoplasias and invasive carcinomas from San Luis Potosí, Mexico: A retrospective cross-sectional study. Infect. Agents Cancer 2015, 10, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taberna, M.; Resteghini, C.; Swanson, B.; Pickard, R.K.; Jiang, B.; Xiao, W.; Mena, M.; Kreinbrink, P.; Chio, E.; Gillison, M.L. Low etiologic fraction for human papillomavirus in larynx squamous cell carcinoma. Oral. Oncol. 2016, 61, 55–61. [Google Scholar] [CrossRef]
- Lam, E.W.H.; Chan, M.M.H.; Wai, C.K.C.; Ngai, C.M.; Chen, Z.; Wong, M.C.S.; Yeung, A.C.M.; Tong, J.H.M.; Chan, A.B.W.; To, K.F.; et al. The role of human papillomavirus in laryngeal cancer in Southern China. J. Med. Virol. 2018, 90, 1150–1159. [Google Scholar] [CrossRef] [PubMed]
- Weiss, D.; Heinkele, T.; Rudack, C. Reliable detection of human papillomavirus in recurrent laryngeal papillomatosis and associated carcinoma of archival tissue. J. Med. Virol. 2015, 87, 860–870. [Google Scholar] [CrossRef]
- Sun, J.; Xiong, J.; Zhen, Y.; Chen, Z.L.; Zhang, H. P53 and PCNA is positively correlated with HPV infection in laryngeal epitheliopapillomatous lesions in patiets with different ethnic backgrounds in Xinjiang. Asian Pac. J. Cancer Prev. 2012, 13, 5439–5444. [Google Scholar] [CrossRef]
- Vietía, D.; Liuzzi, J.; Avila, M.; De Guglielmo, Z.; Prado, Y.; Correnti, M. Human papillomavirus detection in head and neck squamous cell carcinoma. Ecancermedicalscience 2014, 8, 475. [Google Scholar] [CrossRef] [Green Version]
- Barzon, L.; Cappellesso, R.; Peta, E.; Militello, V.; Sinigaglia, A.; Fassan, M.; Simonato, F.; Guzzardo, V.; Ventura, L.; Blandamura, S.; et al. Profiling of expression of human papillomavirus-related cancer miRNAs in penile squamous cell carcinomas. Am. J. Pathol. 2014, 184, 3376–3383. [Google Scholar] [CrossRef]
- Alemany, L.; Cubilla, A.; Halec, G.; Kasamatsu, E.; Quirós, B.; Masferrer, E.; Tous, S.; Lloveras, B.; Hernández-Suarez, G.; Lonsdale, R.; et al. Role of human papillomavirus in penile carcinomas worldwide. Eur. Urol. 2016, 69, 953–961. [Google Scholar] [CrossRef]
- Lechner, M.; Liu, J.; Masterson, L.; Fenton, T.R. HPV-associated oropharyngeal cancer: Epidemiology, molecular biology and clinical mangement. Nat. Rev. Clin. Oncol. 2022, 19, 306–327. [Google Scholar] [CrossRef]
- Serrano, B.; Brotons, M.; Bosch, F.X.; Bruni, L. Epidemiology and burden of HPV-related disease. Clin. Obstet. Gynaecol. 2018, 47, 14–26. [Google Scholar] [CrossRef]
- Mestre, V.F.; Medeiros-Fonseca, B.; Estêvão, D.; Casaca, F.; Silva, S.; Félix, A.; Silva, F.; Colaço, B.; Seixas, F.; Bastos, M.M.; et al. HPV16 is sufficient to induce squamous cell carcinoma specifically in the tongue base in transgenic mice. J. Pathol. 2020, 251, 4–11. [Google Scholar] [CrossRef]
- Medeiros-Fonseca, B.; Mestre, V.F.; Estêvão, D.; Sánchez, D.F.; Cañete-Portillo, S.; Fernández-Nestosa, M.J.; Casaca, F.; Silva, S.; Brito, H.; Félix, A.; et al. HPV16 induces penile intraepithelial neoplasia and squamous cell carcinoma in transgenic mice: First mpouse model for HPV-related penile cancer. J. Pathol. 2020, 251, 411–419. [Google Scholar] [CrossRef]
- Stelzer, M.K.; Pitot, H.C.; Liem, A.; Schweizer, J.; Mahoney, C.; Lambert, P.F. A mouse model for human anal cancer. Cancer Prev. Res. 2010, 3, 1534–1541. [Google Scholar] [CrossRef] [Green Version]
- Cochicho, D.; Nunes, A.; Gomes, J.P.; Martins, L.; Cunha, M.; Medeiros-Fonseca, B.; Oliveira, P.; Bastos, M.M.S.M.; Medeiros, R.; Mendonça, J.; et al. Characterization of the human papillomavirus 16 oncogenes in K14HPV16 mice: Sublineage A1 drives multi-organ carcinogenesis. Int. J. Mol. Sci. 2022, 23, 12371. [Google Scholar] [CrossRef]
- Trottier, H.; Franco, E.L. The epidemiology of genital human papillomavirus infection. Vaccine 2006, 24, S4–S15. [Google Scholar] [CrossRef]
- Garbuglia, A.R.; Gentile, M.; Del Nonno, F.; Lorenzini, P.; Lapa, D.; Lupi, F.; Pinnetti, C.; Baiocchini, A.; Libertone, R.; Cicalini, S.; et al. An anal cancer screening program for MSM in Italy: Prevalence of multiple HPV types and vaccine-targeted infections. J. Clin. Virol. 2015, 72, 49–54. [Google Scholar] [CrossRef]
- Jain, M.A.; Limaiem, F. Cervical Intraepithelial Squamous Cell Lesion; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Cohen, P.A.; Jhingran, A.; Oaknin, A.; Denny, L. Cervical cancer. Lancet 2019, 393, 169–182. [Google Scholar] [CrossRef]
- Li, N.; Franceschi, S.; Howell-Jones, R.; Snijders, P.J.; Clifford, G.M. Human papillomavirus type distribution in 30,848 invasive cervical cancers worldwide: Variation by geographical region, histological type and year of publication. Int. J. Cancer 2011, 128, 927–935. [Google Scholar] [CrossRef]
- Manyere, N.R.; Dube Mandishora, R.S.; Magwali, T.; Mtisi, F.; Mataruka, K.; Mtede, B.; Palefsky, J.M.; Chirenje, Z.M. Human papillomavirus genotype distribution in genital warts among women in Harare-Zimbabwe. J. Obstet. Gynaecol. 2020, 40, 830–836. [Google Scholar] [CrossRef]
- Viau, M.; Renaud, M.C.; Grégoire, J.; Sebastianelli, A.; Plante, M. Paraneoplastic syndromes associated with gynecological cancers: A systematic review. Gynecol. Oncol. 2017, 146, P661–P671. [Google Scholar] [CrossRef]
- Pang, S.S.; Murphy, M.; Markham, M.J. Current management of locally advanced and metastatic cervical cancer in the United States. JCO Oncol. Pract. 2022, 18, 417–422. [Google Scholar] [CrossRef]
- Peixoto da Silva, S.; Santos, J.M.; Costa e Silva, M.P.; Gil da Costa, R.M.; Medeiros, R. Cancer cachexia and its pathophysiology: Links with sarcopenia, anorexia and asthenia. J. Cachexia Sarcopenia Muscle 2020, 11, 619–635. [Google Scholar] [CrossRef]
- Monti, M.; D’Aniello, D.; Scopelliti, A.; Tibaldi, V.; Santangelo, G.; Colagiovanni, V.; Giannini, A.; DI Donato, V.; Palaia, I.; Perniola, G.; et al. Relationship between cervical excisional treatment for cervical intraepithelial neoplasia and obstectrical outcome. Minerva Obstet. Gynecol. 2020, 73, 233–246. [Google Scholar]
- Giannini, A.; Di Donato, V.; Sopracordevole, F.; Ciavattini, A.; Ghelardi, A.; Vizza, E.; D’Oria, O.; Simoncini, T.; Plotti, F.; Casarin, J.; et al. Outcomes of high-grade cervical dysplasia with positive margins anf HPV persistence after cervical conization. Vaccines 2022, 11, 698. [Google Scholar] [CrossRef]
- Cochicho, D.; Esteves, S.; Rito, M.; Silva, F.; Martins, L.; Montalvão, P.; Cunha, M.; Magalhães, M.; Gil da Costa, R.M.; Felix, A. PIK3CA gene mutations in HNSCC: Systematic review and correlations with HPV status and patient survival. Cancers 2022, 14, 1286. [Google Scholar] [CrossRef]
- Rabah, R.; Lancaster, W.D.; Thomas, R.; Gregoire, L. Human papillomavirus-11-associated recurrent respiratory papillomatosis is more aggressive than human papillomavirus-6-associated disease. Pediatr. Dev. Pathol. 2001, 4, 68–72. [Google Scholar] [CrossRef]
- Lee, L.-A.; Cheng, A.-J.; Fang, T.-J.; Huang, C.-G.; Liao, C.-T.; Chang, J.T.-C.; Li, H.-Y. High incidence of malignant transformation of laryngeal papilloma in Taiwan. Laryngoscope 2008, 118, 50–55. [Google Scholar] [CrossRef]
- Wieland, U.; Kreuter, A. HPV-induced anal lesions. Hautarzt 2015, 66, 439–445. [Google Scholar] [CrossRef]
- Zaouak, A.; Ebdelli, W.; Bacha, T.; Koubaa, W.; Hammami, H.; Fenniche, S. Verrucous carcinoma arising in an extended giant condyloma acuminatum. Skinmed 2023, 21, 53–54. [Google Scholar]
- De Sousa, I.D.B.; Vidal, F.C.B.; Vidal, J.P.C.B.; de Mello, G.C.F.; Nascimento, M.D.D.S.B.; Brito, L.M.O. Prevalence of human papillomavirus in penile malignant tumors: Viral genotyping and clinical aspects. BMC Urol. 2015, 15, 13. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Nestosa, M.J.; Guimerà, N.; Sanchez, D.F.; Cañete-Portillo, S.; Velazquez, E.F.; Jenkins, D.; Quint, W.; Cubilla, A.L. Human papillomavirus (HPV) genotypes in condylomas, intraepithelial neoplasia, and invasive carcinoma of the penis using laser capture microdissection (LCM)-PCR: A study of 191 lesions in 43 patients. Am. J. Surg. Pathol. 2017, 41, 820–832. [Google Scholar] [CrossRef]
- Srodon, M.; Stoler, M.H.; Baber, G.B.; Kurman, R.J. The distribution of low and high-risk HPV types in vulvar and vaginal intraepithelial neoplasia (VIN and VaIN). Am. J. Surg. Pathol. 2006, 30, 1513–1518. [Google Scholar] [CrossRef]
- Facio, F.N., Jr.; Facio, M.F.W.; Spessoto, A.C.N.; Godoy, M.; Tessaro, H.; Campos, R.; Zanatto, D.; Calmon, M.; Rahal, P.; Fava, L.C.; et al. Clinical and molecular profile of patients with condyloma acuminatum treated in the Brazilian public healthcare system. Cureus 2022, 14, e21961. [Google Scholar] [CrossRef]
- Horn, L.C.; Klostermann, K.; Hautmann, S.; Höhn, A.K.; Beckmann, M.W.; Mehlhorn, G. HPV-associated alterations of the vulva and vagina. Morphology and molecular pathology. Pathologe 2011, 32, 467–475. [Google Scholar] [CrossRef]
- Cornall, A.M.; Roberts, J.M.; Garland, S.M.; Hillman, R.J.; Grulich, A.E.; Tabrizi, S.N. Anal and perianal squamous carcinomas and high-grade intraepithelial lesions exclusively associated with “low-risk” HPV genotypes 6 and 11. Int. J. Cancer 2013, 133, 2253–2258. [Google Scholar] [CrossRef]
- De Pokomandy, A.; Rouleau, D.; Ghattas, G.; Vézina, S.; Coté, P.; Macleod, J.; Allaire, G.; Franco, E.L.; HIPVIRG Study Group. Prevalence, clearance, and incidence of anal human papillomavirus infection in HIV-infected men: The HIPVIRG Cohort Study. J. Infect. Dis. 2009, 199, 965–973. [Google Scholar] [CrossRef]
- Goldstone, S.; Palefsky, J.M.; Giuliano, A.R.; Moreira, E.D.; Aranda, C.; Jessen, H.; Hillman, R.J.; Ferris, D.G.; Coutlee, F.; Liaw, K.-L.; et al. Prevalence of and risk factors for human papillomavirus (HPV) infection among HIV-seronegative men who have sex with men. J. Infect. Dis. 2011, 203, 66–74. [Google Scholar] [CrossRef]
- Alexandrou, A.; Dimitriou, N.; Levidou, G.; Griniatsos, J.; Sougioultzis, S.; Korkolopoulou, P.; Felekouras, E.; Pikoulis, E.; Diamantis, T.; Tsigris, C.; et al. The incidence of HPV infection in anal cancer patients in Greece. Acta Gastroenterol. Belg. 2014, 77, 213–216. [Google Scholar] [PubMed]
- Egawa, N.; Nakahara, T.; Ohno, S.; Narisawa-Saito, M.; Yugawa, T.; Fujita, M.; Yamato, K.; Natori, Y.; Kiyono, T. The E1 protein of human papillomavirus type 16 is dispensable for maintenance replication of the viral genome. J. Virol. 2012, 86, 3276–3283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, F.J.; Romanos, M.A. E1 protein of human papillomavirus is a DNA helicase/ATPase. Nucleic Acids Res. 1993, 21, 5817–5823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doorbar, J.; Egawa, N.; Griffin, H.; Kranjec, C.; Murakami, I. Human papillomavirus molecular biology and disease association. Rev. Med. Virol. 2015, 25 (Suppl. S1), 2–23. [Google Scholar] [CrossRef] [Green Version]
- Sanders, C.M.; Stenlund, A. Transcription factor-dependent loading of the E1 initiator reveals modular assembly of the papillomavirus origin melting complex. J. Biol. Chem. 2000, 275, 3522–3534. [Google Scholar] [CrossRef] [Green Version]
- Võsa, L.; Sudakov, A.; Remm, M.; Ustav, M.; Kurg, R. Identification and analysis of papillomavirus E2 protein binding sites in the human genome. J. Virol. 2012, 86, 348–357. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Meyers, C.; Wang, H.-K.; Chow, L.T.; Zheng, Z.-M. Construction of a full transcription map of human papillomavirus type 18 during productive viral infection. J. Virol. 2011, 85, 8080–8092. [Google Scholar] [CrossRef] [Green Version]
- Müller, M.; Prescott, E.L.; Wasson, C.W.; Macdonald, A. Human papillomavirus E5 oncoprotein: Function and potential target for antiviral therapeutics. Future Virol. 2015, 10, 27–39. [Google Scholar] [CrossRef]
- DiMaio, D.; Petti, L.M. The E5 proteins. Virology 2013, 445, 99–114. [Google Scholar] [CrossRef] [PubMed]
- Filippova, M.; Johnson, M.M.; Bautista, M.; Filippov, V.; Fodor, N.; Tungteakkhun, S.S.; Williams, K.; Duerksen-Hughes, P.J. The large and small isoforms of human papillomavirus type 16 E6 bind to and differentially affect procaspase 8 stability and activity. J. Virol. 2007, 81, 4116–4129. [Google Scholar] [CrossRef] [Green Version]
- Genther Williams, S.M.; Disbrow, G.L.; Schlegel, R.; Lee, D.; Threadgill, D.W.; Lambert, P.F. Requirement of epidermal growth factor receptor for hyperplasia induced by E5, a high-risk human papillomavirus oncogene. Cancer Res. 2005, 65, 6534–6542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roman, A.; Munger, K. The papillomavirus E7 proteins. Virology 2013, 445, 138–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLaughlin-Drubin, M.E.; Bromberg-White, J.L.; Meyers, C. The role of the human papillomavirus type 18 E7 oncoprotein during the complete viral life cycle. Virology 2005, 338, 61–68. [Google Scholar] [CrossRef] [Green Version]
- Egawa, N.; Doorbar, J. The low-risk papillomaviruses. Virus Res. 2017, 231, 119–127. [Google Scholar] [CrossRef]
- Barrow-Laing, L.; Chen, W.; Romão, A. Low- and high-risk human papillomavirusE7 proteins regulate p130 differently. Virology 2010, 400, 233–239. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Chen, W.; Roman, A. The E7 proteins of low- and high-risk human papillomaviruses share the ability to target the pRB family member p130 for degradation. Proc. Natl. Acad. Sci. USA 2006, 103, 437–442. [Google Scholar] [CrossRef]
- Genovese, N.J.; Broker, T.R.; Chow, L.T. Nonconserved lysine residues attenuate the biological function of the low-risk human papillomavirus E7 protein. J. Virol. 2011, 85, 5546–5554. [Google Scholar] [CrossRef] [Green Version]
- Singhi, A.D.; Westra, W.H. Comparison of human papillomavirus in situ hybridization and p16 immunohistochemistry in the detection of human papillomavirus-associated head and neck cancer based on a prospective clinical experience. Cancer 2010, 116, 2166–2173. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| References | Type of Sample | Detection Method | Genotyping Method | Total Sample and (HPV + Sample) |
|---|---|---|---|---|
| Tao et al., 2017 [25] | Cervical scrappings | PCR/Luminex 200 (Tellgen, Shanghai, China) | PCR/Luminex 200 (Tellgen, China) | 142 (142) |
| Das et al., 2013 [26] | Tissue biopsy in RNAlater (Qiagen, Hilden, Germany) | Nested PCR | Digene HPV Hybrid Capture II Test (Qiagen, Germany) | 107 (105) |
| Aldersley et al., 2021 [27] | Whole exome data from previous publications | SureSelect Exon Capture (Agilent, Santa Clara, CA, USA) and HiSeq sequencing (Illumina, San Diego, CA, USA) | SureSelect Exon Capture (Agilent) and HiSeq sequencing (Illumina, USA) | 72 (62) |
| Barzon et al., 2014 [34] | FFPE | PCR (Inno-LiPa, Tokyo, Japan) | Real-time PCR | 59 (18) |
| Alemany et al., 2016 [35] | FFPE | SPF-10/DEIA/LIPA25 (Laboratory Biomedical Products, Rijswijk, The Netherlands) | LIPA25 (Laboratory Biomedical Products, The Netherlands) and Sanger sequencing | 1010 (334) |
| Vietía et al., 2014 [33] | Fresh biopsies | PCR (Inno-LiPa, Japan) | PCR (Inno-LiPa, Japan) | 71 (48) |
| Taberna et al., 2016 [29] | FFPE | PCR (Inno-LiPa, Japan) | Real-Time PCR | 404 (54) |
| Lam et al., 2018 [30] | FFPE | Nested PCR | Sanger sequncing | 85 (1) |
| Weiss et al., 2015 [31] | FFPE | Real-Time PCR GP5+/6+ and In Situ Hybridization | Real-Time PCR | 8 (6) |
| Sun et al., 2012 [32] | FFPE | PCR for HPV 6/11 and HPV 16/18 | PCR for HPV 6/11 and HPV 16/18 | 83 (42) |
| Magaña-León et al., 2015 [28] | FFPE | SPF-10/DEIA/LIPA25 (Laboratory Biomedical Products, The Netherlands) | PCR (Inno-LiPa, Japan) | 35 (10) |
| Anatomic Location | HPV6 % (n/N) | HPV11 % (n/N) | Multiple Low-Risk% (n/N) | Geographical Location | References | |
|---|---|---|---|---|---|---|
| Uterine cervix | 0/142 | 1/142 | 1/142 | China | Tao et al., 2017 [25] | |
| 0/105 | 0/105 | 0/105 | India | Das et al., 2013 [26] | ||
| 1.4% (1/62) | 0/62 | 1/62 | Republic of Korea/United States/France | Aldersley et al., 2021 [27] | ||
| Penis | 0/18 | 5.5% (1/18) | 1/18 | Italy | Barzon et al., 2014 [34] | |
| 3.6% (12/334) | 1.2% (4/334) | 4.8% (16/334) | 25 Countries | Alemany et al., 2016 [35] | ||
| 0/8 | 0/8 | 0/8 | Mexico | Magaña-León et al., 2015 [28] | ||
| Head and neck | Oral cavity | 0/9 | 0/9 | 0/9 | Mexico | Magaña-León et al., 2015 [28] |
| Larynx | 3.7% (2/54) | 3.7% (2/54) | 4/54 | United States | Taberna et al., 2016 [29] | |
| 1.2% (1/85) | 0/85 | 1/85 | China | Lam et al., 2018 [30] | ||
| 75.0% (6/8) | 12.5% (1/8) | 7/8 | Germany | Weiss et al., 2015 [31] | ||
| Not tested | Not tested | 9.6% (8/42) | China | Sun et al., 2012 [32] | ||
| 0/9 | 0/9 | 0/9 | Mexico | Magaña-León et al., 2015 [28] | ||
| Mixed locations | 12.5% (6/48) | 0% (0/48) | 16.67% (8/48) | Venezuela | Vietía et al., 2014 [33] | |
| Vagina | 0/7 | 0/7 | 1/7 * | Mexico | Magaña-León et al., 2015 [28] | |
| Vulva | 0/1 | 0/1 | 0/1 | Mexico | Magaña-León et al., 2015 [28] | |
| HPV6 | |||||
|---|---|---|---|---|---|
| E6 (NP_040296.1) | E7 (NP_040297.1) | E5A (NP_040301.1) | E5B (NP_040302.1) | ||
| HPV16 | E6 (NP_041325.1) | Identities: 39%; Positives: 60%; Expect: 2 × 10−41 | |||
| E7 (NP_041326.1) | Identities: 57%; Positives: 69%; Expect: 3 × 10−34 | ||||
| E5 (NP_041330.2) | Identities: 24%; Positives: 58%; Expect: 0.018 | No significant similarity found. | |||
| HPV11 | |||||
|---|---|---|---|---|---|
| E6 (QXM18822.1) | E7 (QXM18823.1) | E5A (QXM18827.1) | E5B (QXM18828.1) | ||
| HPV16 | E6 (NP_041325.1) | Identities: 37%; Positives: 61%; Expect: 2 × 10−40 | |||
| E7 (NP_041326.1) | Identities: 55%; Positives: 70%; Expect: 1 × 10−33 | ||||
| E5 (NP_041330.2) | No significant similarity found. | No significant similarity found. | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, L.L.d.; Teles, A.M.; Santos, J.M.O.; Souza de Andrade, M.; Medeiros, R.; Faustino-Rocha, A.I.; Oliveira, P.A.; dos Santos, A.P.A.; Ferreira Lopes, F.; Braz, G.; et al. Malignancy Associated with Low-Risk HPV6 and HPV11: A Systematic Review and Implications for Cancer Prevention. Cancers 2023, 15, 4068. https://doi.org/10.3390/cancers15164068
Silva LLd, Teles AM, Santos JMO, Souza de Andrade M, Medeiros R, Faustino-Rocha AI, Oliveira PA, dos Santos APA, Ferreira Lopes F, Braz G, et al. Malignancy Associated with Low-Risk HPV6 and HPV11: A Systematic Review and Implications for Cancer Prevention. Cancers. 2023; 15(16):4068. https://doi.org/10.3390/cancers15164068
Chicago/Turabian StyleSilva, Leandro Lima da, Amanda Mara Teles, Joana M. O. Santos, Marcelo Souza de Andrade, Rui Medeiros, Ana I. Faustino-Rocha, Paula A. Oliveira, Ana Paula Azevedo dos Santos, Fernanda Ferreira Lopes, Geraldo Braz, and et al. 2023. "Malignancy Associated with Low-Risk HPV6 and HPV11: A Systematic Review and Implications for Cancer Prevention" Cancers 15, no. 16: 4068. https://doi.org/10.3390/cancers15164068