
The Effects of Heat Stress on the Transcriptome of Human Cancer Cells: A Meta-Analysis
 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials & Methods
2.1. Data Collection and Curation
2.2. Acquisition and Processing of Unpublished Gene Expression Data
2.3. Meta-Analysis
2.4. Experimental Validation of Gene Expression Changes
3. Results
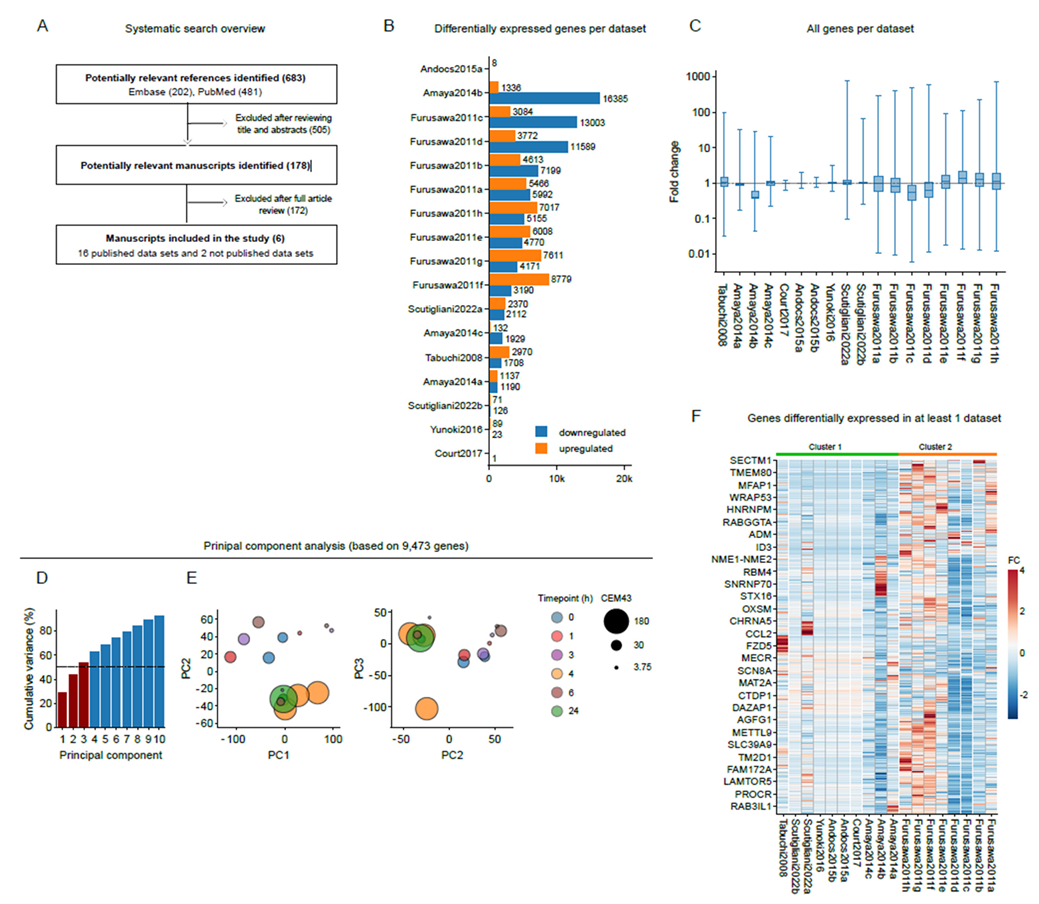
3.1. Quantification of Gene Expression after Heat Stress in Bladder and Cervix Cancer Cell Lines
3.2. Gene Expression Patterns Are Independent from Key Experimental Parameters
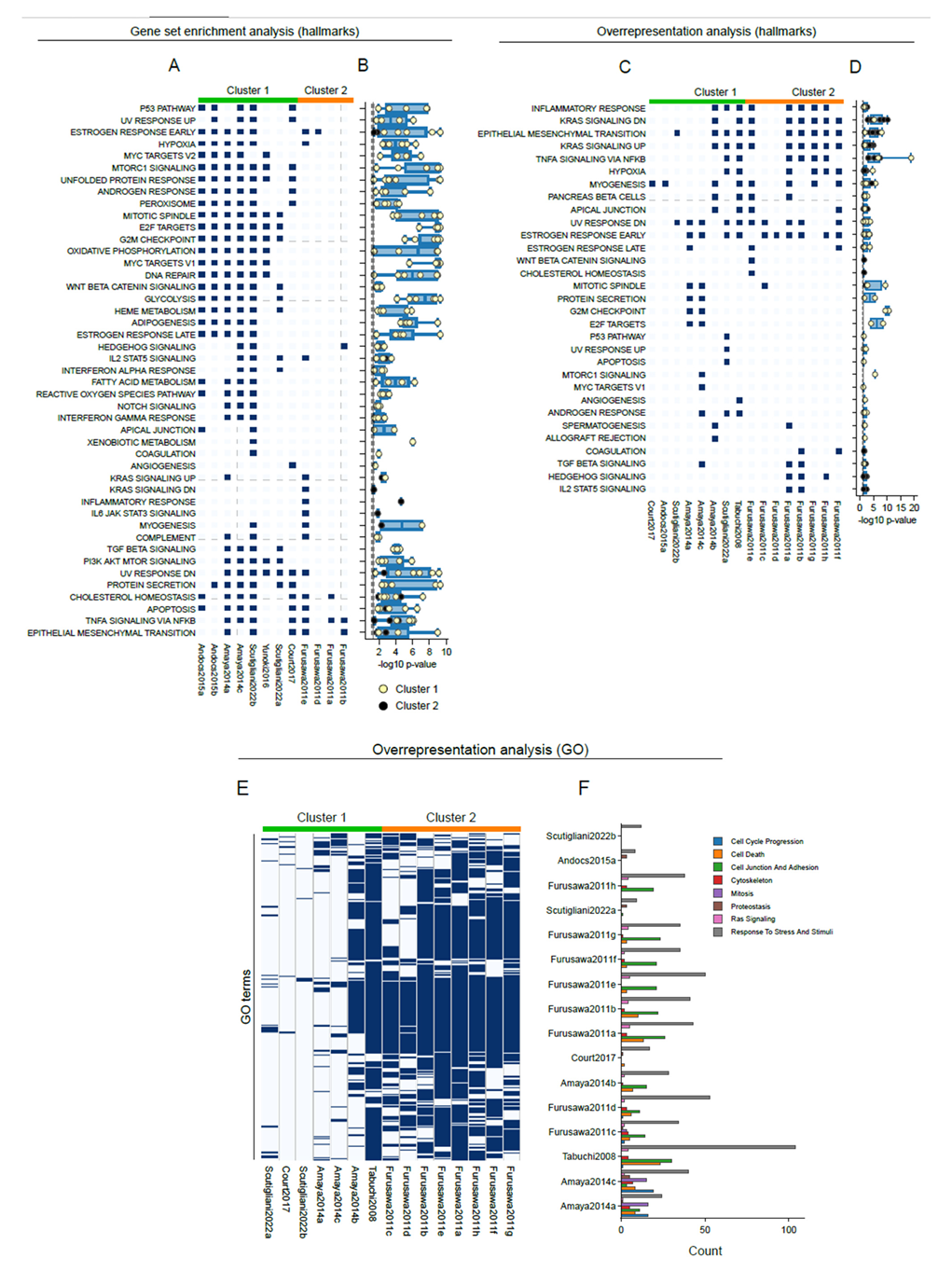
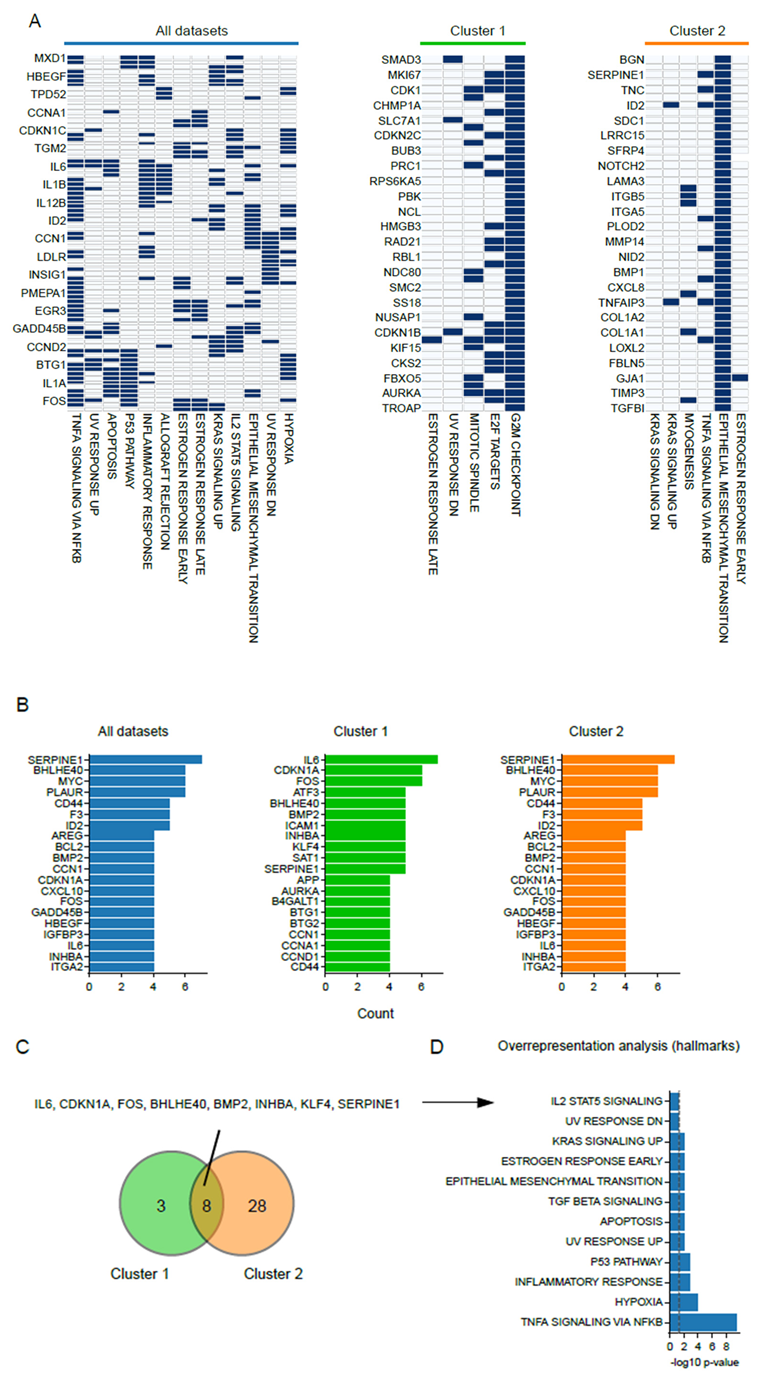
3.3. Common Patterns in Transcriptome Changes at a Pathway Level
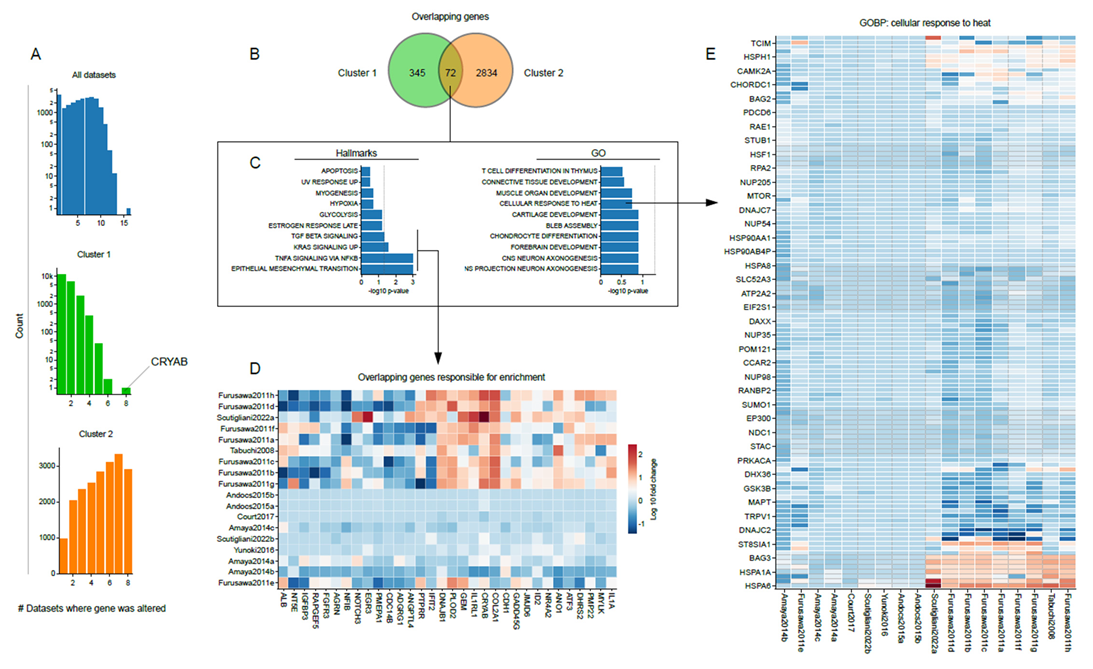
3.4. Shared Transcriptome Changes Are Driven by Highly Variable Gene Expression Patterns
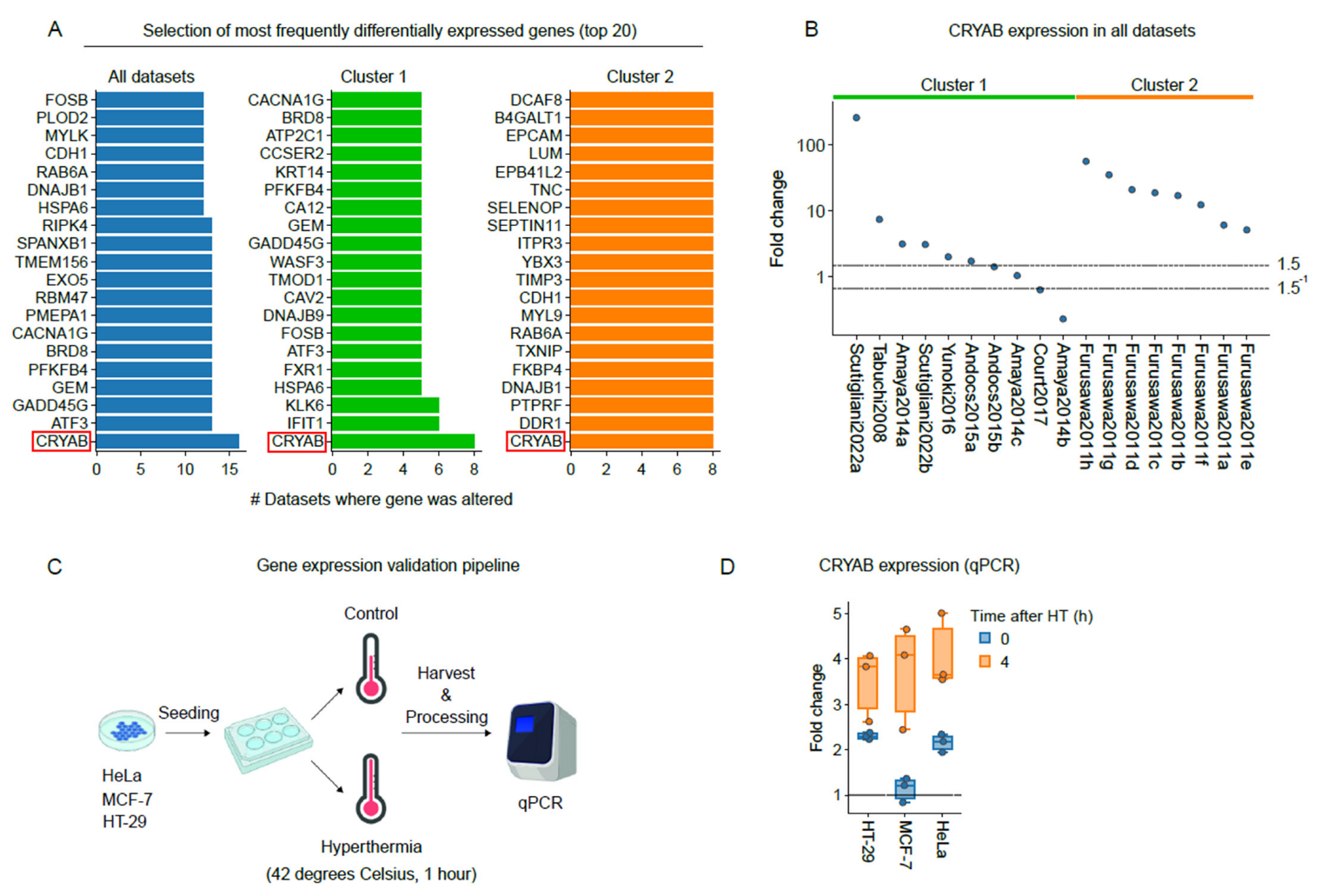
3.5. The Small Heat-Shock Protein CRYAB Is Most Commonly Overexpressed after Hyperthermia
4. Discussions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Van der Zee, J. Heating the Patient: A Promising Approach? Ann. Oncol. 2002, 13, 1173–1184. [Google Scholar] [CrossRef] [PubMed]
- Datta, N.R.; Ordóñez, S.G.; Gaipl, U.S.; Paulides, M.M.; Crezee, H.; Gellermann, J.; Marder, D.; Puric, E.; Bodis, S. Local Hyperthermia Combined with Radiotherapy and-/or Chemotherapy: Recent Advances and Promises for the Future. Cancer Treat. Rev. 2015, 41, 742–753. [Google Scholar] [CrossRef] [PubMed]
- Datta, N.R.; Puric, E.; Klingbiel, D.; Gomez, S.; Bodis, S. Hyperthermia and Radiation Therapy in Locoregional Recurrent Breast Cancers: A Systematic Review and Meta-Analysis. Int. J. Radiat. Oncol. Biol. Phys. 2016, 94, 1073–1087. [Google Scholar] [CrossRef] [PubMed]
- Datta, N.R.; Rogers, S.; Ordóñez, S.G.; Puric, E.; Bodis, S. Hyperthermia and Radiotherapy in the Management of Head and Neck Cancers: A Systematic Review and Meta-Analysis. Int. J. Hyperth. 2016, 32, 31–40. [Google Scholar] [CrossRef]
- Crezee, H.; Van Leeuwen, C.M.; Oei, A.L.; Stalpers, L.J.A.; Bel, A.; Franken, N.A.; Kok, H.P. Thermoradiotherapy Planning: Integration in Routine Clinical Practice. Int. J. Hyperth. 2016, 32, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Dewhirst, M.W.; Vujaskovic, Z.; Jones, E.; Thrall, D. Re-Setting the Biologic Rationale for Thermal Therapy. Int. J. Hyperth. 2005, 21, 779–790. [Google Scholar] [CrossRef]
- Van Den Tempel, N.; Horsman, M.R.; Kanaar, R. Improving Efficacy of Hyperthermia in Oncology by Exploiting Biological Mechanisms. Int. J. Hyperth. 2016, 32, 446–454. [Google Scholar] [CrossRef] [Green Version]
- Richter, K.; Haslbeck, M.; Buchner, J. The Heat Shock Response: Life on the Verge of Death. Mol. Cell 2010, 40, 253–266. [Google Scholar] [CrossRef]
- Scutigliani, E.M.; Liang, Y.; Crezee, H.; Kanaar, R.; Krawczyk, P.M. Modulating the Heat Stress Response to Improve Hyperthermia-Based Anticancer Treatments. Cancers 2021, 13, 1243. [Google Scholar] [CrossRef]
- van Rhoon, G.C.; Franckena, M.; ten Hagen, T.L.M. A Moderate Thermal Dose Is Sufficient for Effective Free and TSL Based Thermochemotherapy. Adv. Drug Deliv. Rev. 2020, 163–164, 145–156. [Google Scholar] [CrossRef]
- Jones, E.L.; Prosnitz, L.R.; Dewhirst, M.W.; Marcom, P.K.; Hardenbergh, P.H.; Marks, L.B.; Brizel, D.M.; Vujaskovic, Z. Thermochemoradiotherapy Improves Oxygenation in Locally Advanced Breast Cancer. Clin. Cancer Res. 2004, 10, 4287–4293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masunaga, S.; Ono, K.; Akaboshi, M.; Nishimura, Y.; Suzuki, M.; Kinashi, Y.; Takagaki, M.; Hiraoka, M.; Abe, M. Reduction of Hypoxic Cells in Solid Tumours Induced by Mild Hyperthermia: Special Reference to Differences in Changes in the Hypoxic Fraction between Total and Quiescent Cell Populations. Br. J. Cancer 1997, 76, 588–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oleson, J.R. Eugene Robertson Special Lecture Hyperthermia from the Clinic to the Laboratory: A Hypothesis. Int. J. Hyperth. 1995, 11, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Song, C.W.; Shakil, A.; Osborn, J.L.; Iwata, K. Tumour Oxygenation Is Increased by Hyperthermia at Mild Temperatures. Int. J. Hyperth. 1996, 12, 367–373. [Google Scholar] [CrossRef]
- Jain, R.K.; Grantham, F.H.; Gullino, P.M. Blood Flow and Heat Transfer in Walker 256 Mammary Carcinoma. J. Natl. Cancer Inst. 1979, 62, 927–933. [Google Scholar]
- Patterson, J.; Strang, R. The Role of Blood Flow in Hyperthermia. Int. J. Radiat. Oncol. Biol. Phys. 1979, 5, 235–241. [Google Scholar] [CrossRef]
- Vaupel, P.; Kallinowski, F. Physiological Effects of Hyperthermia. Recent Results Cancer Res. 1987, 104, 71–109. [Google Scholar]
- Wu, T.; Dai, Y. Tumor Microenvironment and Therapeutic Response. Cancer Lett. 2017, 387, 61–68. [Google Scholar] [CrossRef]
- Thews, O.; Riemann, A. Tumor pH and Metastasis: A Malignant Process beyond Hypoxia. Cancer Metastasis Rev. 2019, 38, 113–129. [Google Scholar] [CrossRef]
- Horsman, M.R.; Overgaard, J. The Impact of Hypoxia and Its Modification of the Outcome of Radiotherapy. J. Radiat. Res. 2016, 57, i90–i98. [Google Scholar] [CrossRef] [Green Version]
- Toraya-Brown, S.; Fiering, S. Local Tumour Hyperthermia as Immunotherapy for Metastatic Cancer. Int. J. Hyperth. 2014, 30, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Deng, J.; Sun, J.; Ma, Y. Hyperthermia Targeting the Tumor Microenvironment Facilitates Immune Checkpoint Inhibitors. Front. Immunol. 2020, 11, 595207. [Google Scholar] [CrossRef] [PubMed]
- Lindquist, S. The Heat-Shock Response. Annu. Rev. Biochem. 1986, 55, 1151–1191. [Google Scholar] [CrossRef] [PubMed]
- Gong, W.J.; Golic, K.G. Loss of Hsp70 in Drosophila Is Pleiotropic, with Effects on Thermotolerance, Recovery from Heat Shock and Neurodegeneration. Genetics 2006, 172, 275–286. [Google Scholar] [CrossRef] [Green Version]
- Harder, Y.; Amon, M.; Schramm, R.; Georgi, M.; Banic, A.; Erni, D.; Menger, M.D. Heat Shock Preconditioning Reduces Ischemic Tissue Necrosis by Heat Shock Protein (HSP)-32-Mediated Improvement of the Microcirculation rather than Induction of Ischemic Tolerance. Ann. Surg. 2005, 242, 869–878, discussion 878–879. [Google Scholar] [CrossRef]
- Watson, R.W.G.; Lebret, T.; Fitzpatrick, J.M. Heat Shock Proteins in the Genitourinary System. Curr. Urol. Rep. 2003, 4, 70–76. [Google Scholar] [CrossRef]
- Gasch, A.P.; Spellman, P.T.; Kao, C.M.; Carmel-Harel, O.; Eisen, M.B.; Storz, G.; Botstein, D.; Brown, P.O. Genomic Expression Programs in the Response of Yeast Cells to Environmental Changes. Mol. Biol. Cell 2000, 11, 4241–4257. [Google Scholar] [CrossRef]
- Sørensen, J.G.; Nielsen, M.M.; Kruhøffer, M.; Justesen, J.; Loeschcke, V. Full Genome Gene Expression Analysis of the Heat Stress Response in Drosophila Melanogaster. Cell Stress Chaperones 2005, 10, 312–328. [Google Scholar] [CrossRef]
- Brown, J.B.; Boley, N.; Eisman, R.; May, G.E.; Stoiber, M.H.; Duff, M.O.; Booth, B.W.; Wen, J.; Park, S.; Suzuki, A.M.; et al. Diversity and Dynamics of the Drosophila Transcriptome. Nature 2014, 512, 393–399. [Google Scholar] [CrossRef] [Green Version]
- Trinklein, N.D.; Murray, J.I.; Hartman, S.J.; Botstein, D.; Myers, R.M. The Role of Heat Shock Transcription Factor 1 in the Genome-Wide Regulation of the Mammalian Heat Shock Response. Mol. Biol. Cell 2004, 15, 1254–1261. [Google Scholar] [CrossRef]
- Vihervaara, A.; Duarte, F.M.; Lis, J.T. Molecular Mechanisms Driving Transcriptional Stress Responses. Nat. Rev. Genet. 2018, 19, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Vihervaara, A.; Mahat, D.B.; Guertin, M.J.; Chu, T.; Danko, C.G.; Lis, J.T.; Sistonen, L. Transcriptional Response to Stress Is Pre-Wired by Promoter and Enhancer Architecture. Nat. Commun. 2017, 8, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Mahat, D.B.; Salamanca, H.H.; Duarte, F.M.; Danko, C.G.; Lis, J.T. Mammalian Heat Shock Response and Mechanisms Underlying Its Genome-Wide Transcriptional Regulation. Mol. Cell 2016, 62, 63–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duarte, F.M.; Fuda, N.J.; Mahat, D.B.; Core, L.J.; Guertin, M.J.; Lis, J.T. Transcription Factors GAF and HSF Act at Distinct Regulatory Steps to Modulate Stress-Induced Gene Activation. Genes Dev. 2016, 30, 1731–1746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, J.; Munn, P.R.; Vihervaara, A.; Lewis, J.J.; Ozer, A.; Danko, C.G.; Lis, J.T. Chromatin Conformation Remains Stable upon Extensive Transcriptional Changes Driven by Heat Shock. Proc. Natl. Acad. Sci. USA 2019, 116, 19431–19439. [Google Scholar] [CrossRef] [Green Version]
- Guertin, M.J.; Lis, J.T. Chromatin Landscape Dictates HSF Binding to Target DNA Elements. PLoS Genet. 2010, 6, e1001114. [Google Scholar] [CrossRef] [Green Version]
- Mendillo, M.L.; Santagata, S.; Koeva, M.; Bell, G.W.; Hu, R.; Tamimi, R.M.; Fraenkel, E.; Ince, T.A.; Whitesell, L.; Lindquist, S. HSF1 Drives a Transcriptional Program Distinct from Heat Shock to Support Highly Malignant Human Cancers. Cell 2012, 150, 549–562. [Google Scholar] [CrossRef] [Green Version]
- Rohlin, L.; Trent, J.D.; Salmon, K.; Kim, U.; Gunsalus, R.P.; Liao, J.C. Heat Shock Response of Archaeoglobus Fulgidus. J. Bacteriol. 2005, 187, 6046–6057. [Google Scholar] [CrossRef] [Green Version]
- Richmond, C.S.; Glasner, J.D.; Mau, R.; Jin, H.; Blattner, F.R. Genome-Wide Expression Profiling in Escherichia Coli K-12. Nucleic Acids Res. 1999, 27, 3821–3835. [Google Scholar] [CrossRef]
- Eisen, M.B.; Spellman, P.T.; Brown, P.O.; Botstein, D. Cluster Analysis and Display of Genome-Wide Expression Patterns. Proc. Natl. Acad. Sci. USA 1998, 95, 14863–14868. [Google Scholar] [CrossRef] [Green Version]
- Matsuura, H.; Ishibashi, Y.; Shinmyo, A.; Kanaya, S.; Kato, K. Genome-Wide Analyses of Early Translational Responses to Elevated Temperature and High Salinity in Arabidopsis Thaliana. Plant Cell Physiol. 2010, 51, 448–462. [Google Scholar] [CrossRef] [PubMed]
- GuhaThakurta, D.; Palomar, L.; Stormo, G.D. Identification of a Novel Cis-Regulatory Element Involved in the Heat Shock Response in Caenorhabditis Elegans Using Microarray Gene Expression and Computational Methods. Genome Res. 2002, 12, 701–712. [Google Scholar] [PubMed]
- Tabuchi, Y.; Takasaki, I.; Wada, S.; Zhao, Q.-L.; Hori, T.; Nomura, T.; Ohtsuka, K.; Kondo, T. Genes and Genetic Networks Responsive to Mild Hyperthermia in Human Lymphoma U937 Cells. Int. J. Hyperth. 2008, 24, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Court, K.A.; Hatakeyama, H.; Wu, S.Y.; Lingegowda, M.S.; Rodríguez-Aguayo, C.; López-Berestein, G.; Ju-Seog, L.; Rinaldi, C.; Juan, E.J.; Sood, A.K.; et al. HSP70 Inhibition Synergistically Enhances the Effects of Magnetic Fluid Hyperthermia in Ovarian Cancer. Mol. Cancer Ther. 2017, 16, 966–976. [Google Scholar] [CrossRef] [Green Version]
- Amaya, C.; Kurisetty, V.; Stiles, J.; Nyakeriga, A.M.; Arumugam, A.; Lakshmanaswamy, R.; Botez, C.E.; Mitchell, D.C.; Bryan, B.A. A Genomics Approach to Identify Susceptibilities of Breast Cancer Cells to “fever-Range” Hyperthermia. BMC Cancer 2014, 14, 1–28. [Google Scholar] [CrossRef] [Green Version]
- Andocs, G.; Rehman, M.U.; Zhao, Q.-L.; Tabuchi, Y.; Kanamori, M.; Kondo, T. Comparison of Biological Effects of Modulated Electro-Hyperthermia and Conventional Heat Treatment in Human Lymphoma U937 Cells. Cell Death Discov. 2016, 2, 16039. [Google Scholar] [CrossRef] [Green Version]
- Yunoki, T.; Tabuchi, Y.; Hayashi, A.; Kondo, T. Network Analysis of Genes Involved in the Enhancement of Hyperthermia Sensitivity by the Knockdown of BAG3 in Human Oral Squamous Cell Carcinoma Cells. Int. J. Mol. Med. 2016, 38, 236–242. [Google Scholar] [CrossRef] [Green Version]
- Furusawa, Y.; Tabuchi, Y.; Wada, S.; Takasaki, I.; Ohtsuka, K.; Kondo, T. Identification of Biological Functions and Gene Networks Regulated by Heat Stress in U937 Human Lymphoma Cells. Int. J. Mol. Med. 2011, 28, 143–151. [Google Scholar]
- Andocs, G.; Meggyeshazi, N.; Balogh, L.; Spisak, S.; Maros, M.E.; Balla, P.; Kiszner, G.; Teleki, I.; Kovago, C.; Krenacs, T. Upregulation of Heat Shock Proteins and the Promotion of Damage-Associated Molecular Pattern Signals in a Colorectal Cancer Model by Modulated Electrohyperthermia. Cell Stress Chaperones 2015, 20, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene Set Enrichment Analysis: A Knowledge-Based Approach for Interpreting Genome-Wide Expression Profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liberzon, A.; Subramanian, A.; Pinchback, R.; Thorvaldsdóttir, H.; Tamayo, P.; Mesirov, J.P. Molecular Signatures Database (MSigDB) 3.0. Bioinformatics 2011, 27, 1739–1740. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. clusterProfiler 4.0: A Universal Enrichment Tool for Interpreting Omics Data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef] [PubMed]
- Whitney, J.; Carswell, W.; Rylander, N. Arrhenius Parameter Determination as a Function of Heating Method and Cellular Microenvironment Based on Spatial Cell Viability Analysis. Int. J. Hyperth. 2013, 29, 281–295. [Google Scholar] [CrossRef]
- Yang, K.-L.; Huang, C.-C.; Chi, M.-S.; Chiang, H.-C.; Wang, Y.-S.; Hsia, C.-C.; Andocs, G.; Wang, H.-E.; Chi, K.-H. In Vitro Comparison of Conventional Hyperthermia and Modulated Electro-Hyperthermia. Oncotarget 2016, 7, 84082–84092. [Google Scholar] [CrossRef] [Green Version]
- Sapareto, S.A.; Dewey, W.C. Thermal Dose Determination in Cancer Therapy. Int. J. Radiat. Oncol. Biol. Phys. 1984, 10, 787–800. [Google Scholar] [CrossRef]
- Franckena, M.; Fatehi, D.; de Bruijne, M.; Canters, R.A.M.; van Norden, Y.; Mens, J.W.; van Rhoon, G.C.; van der Zee, J. Hyperthermia Dose-Effect Relationship in 420 Patients with Cervical Cancer Treated with Combined Radiotherapy and Hyperthermia. Eur. J. Cancer 2009, 45, 1969–1978. [Google Scholar] [CrossRef] [PubMed]
- Maguire, P.D.; Samulski, T.V.; Prosnitz, L.R.; Jones, E.L.; Rosner, G.L.; Powers, B.; Layfield, L.W.; Brizel, D.M.; Scully, S.P.; Harrelson, J.M.; et al. A Phase II Trial Testing the Thermal Dose Parameter CEM43 Degrees T90 as a Predictor of Response in Soft Tissue Sarcomas Treated with Pre-Operative Thermoradiotherapy. Int. J. Hyperth. 2001, 17, 283–290. [Google Scholar] [CrossRef]
- de Bruijne, M.; van der Holt, B.; van Rhoon, G.C.; van der Zee, J. Evaluation of CEM43 Degrees CT90 Thermal Dose in Superficial Hyperthermia: A Retrospective Analysis. Strahlenther. Onkol. 2010, 186, 436–443. [Google Scholar] [CrossRef]
- Tang, S.; Yin, B.; Xu, J.; Bao, E. Rosemary Reduces Heat Stress by Inducing CRYAB and HSP70 Expression in Broiler Chickens. Oxid. Med. Cell. Longev. 2018, 2018, 7014126. [Google Scholar] [CrossRef] [Green Version]
- Murray, J.I.; Whitfield, M.L.; Trinklein, N.D.; Myers, R.M.; Brown, P.O.; Botstein, D. Diverse and Specific Gene Expression Responses to Stresses in Cultured Human Cells. Mol. Biol. Cell 2004, 15, 2361–2374. [Google Scholar] [CrossRef]
- Wang, Q.-L.; Liu, B.O.; Li, X.-J.; Hu, K.-P.; Zhao, K.; Ye, X.-M. Inhibition of mTOR Promotes Hyperthermia Sensitivity in SMMC-7721 Human Hepatocellular Carcinoma Cell Line. Exp. Ther. Med. 2016, 11, 961–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Duan, S.; Zheng, Y.; Yang, Y.; Wang, L.; Li, X.; Zhang, Q.; Thorne, R.F.; Li, W.; Yang, D. Hyperthermia Inhibits Growth of Nasopharyngeal Carcinoma through Degradation of c-Myc. Int. J. Hyperth. 2022, 39, 358–371. [Google Scholar] [CrossRef] [PubMed]
- Streffer, C. Aspects of Metabolic Change after Hyperthermia. Recent Results Cancer Res. 1988, 107, 7–16. [Google Scholar] [PubMed]
- Kanamori, T.; Miyazaki, N.; Aoki, S.; Ito, K.; Hisaka, A.; Hatakeyama, H. Investigation of Energy Metabolic Dynamism in Hyperthermia-Resistant Ovarian and Uterine Cancer Cells under Heat Stress. Sci. Rep. 2021, 11, 14726. [Google Scholar] [CrossRef] [PubMed]
- Bordonaro, M.; Shirasawa, S.; Lazarova, D.L. In Hyperthermia Increased ERK and WNT Signaling Suppress Colorectal Cancer Cell Growth. Cancers 2016, 8, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dellinger, T.H.; Han, E.S.; Raoof, M.; Lee, B.; Wu, X.; Cho, H.; He, T.-F.; Lee, P.; Razavi, M.; Liang, W.S.; et al. Hyperthermic Intraperitoneal Chemotherapy-Induced Molecular Changes in Humans Validate Preclinical Data in Ovarian Cancer. JCO Precis. Oncol. 2022, 6, e2100239. [Google Scholar] [CrossRef] [PubMed]
- Kokura, S.; Yoshida, N.; Ueda, M.; Imamoto, E.; Ishikawa, T.; Takagi, T.; Naito, Y.; Okanoue, T.; Yoshikawa, T. Hyperthermia Enhances Tumor Necrosis Factor Alpha-Induced Apoptosis of a Human Gastric Cancer Cell Line. Cancer Lett. 2003, 201, 89–96. [Google Scholar] [CrossRef]
- Nakabe, N.; Kokura, S.; Shimozawa, M.; Katada, K.; Sakamoto, N.; Ishikawa, T.; Handa, O.; Takagi, T.; Naito, Y.; Yoshida, N.; et al. Hyperthermia Attenuates TNF-Alpha-Induced up Regulation of Endothelial Cell Adhesion Molecules in Human Arterial Endothelial Cells. Int. J. Hyperth. 2007, 23, 217–224. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | GEO Accession | Aim | Highlighted Pathways and Functions | Highlighted Genes |
|---|---|---|---|---|
| Tabuchi et al., 2008 [43] | GSE10043 | Examine gene expression patterns in human myelomonocytic lymphoma U937 cells exposed to mild hyperthermia | Up- or downregulated:
| Unfolded protein response-related genes were upregulated: Hsp40 homologs (DNAJA1, DNAJB1), Hsp70 proteins (HSPA6, HSPA1A, HSPA1B, HSPA1L), HSPB1 (heat shock 27 kDa protein 1), HSPH1 (heat shock 105 kDa/110 kDa protein 1), PPP1R15A (protein phosphatase 1, regulatory (inhibitor) subunit 15 A), and SERPINH1 (serpin peptidase inhibitor, clade H (heat shock protein 47), member 1). |
| Furusawa et al., 2011 [48] | GSE23405 | Understand the molecular mechanisms underlying cellular responses to heat stress at temperatures higher and lower than the inflection point of hyperthermia | Upregulated:
| Peak expression of HSPs was observed 3 h after heat stress. The expression level of HSPs such as the Hsp70 (HSPA6, HSPA1A), Hsp40 (DNAJA1, DNAJB1) and Hsp27 (HSPB1) gene subfamilies was gradually elevated at 44 ˚C. |
| Amaya et al., 2014 [45] | GSE48398 | Identify the unique gene networks distinct between normal and cancer cell lines following hyperthermia | Upregulated:
| Mitotic regulatory genes were up-regulated: STAG2, NEK2, KPNA4, IPO5, TNPO1, CCNB1, CDK1, CDK6, NCAPG, NCAPG2, TOP2A, NUF2, CENPE, CENPF, ZWILCH, PDS5A, WEE1, KIF11, CHUK, and PPP1CB. |
| Court et al., 2017 [44] | GSE92990 | Investigate gene expression profiles after magnetic fluid hyperthermia in ovarian cancer cell lines to elucidate cellular response and select molecular targets to enhance its effect in vitro and in vivo | Upregulated:
| Top genes related to the aforementioned functions affected by magnetic fluid hyperthermia were HSPs, Hsp70 (HSPA6/HSPA7, HSPA1A, HSPA1B, HSPA1L, HSPA4L), Hsp60 (LOC643300), Hsp40 (DNBAJ family), Hsp20 (CRYAB) and Hsp27 (SERPINH1), and BAG3 (modulator of Hsp70). |
| Andocs et al. 2015 [49] | GSE58750 | Identify the gene expression alterations induced by heat treatment in human tumor HT29 colorectal cancer xenograft mouse model | Upregulated:
| Members of the heat shock protein 70 family including HSPA1A, HSPA1B, HSPA4, HSPA6, and HSPA8, and their co-chaperones Hsp40 (DNAJB1 and DNAJB4) and Bag3 became upregulated. Hsp90 alpha (HSP90AA1) and Hsp60 (HSPD1) gene transcripts were also elevated upon hyperthermia treatment. |
| Yunoki et al., 2016 [47] | GSE75127 | Identify gene networks involved in the enhancement of hyperthermia sensitivity by the knockdown of BAG3 in human oral squamous cell carcinoma cells | Upregulated:
| Genes associated with HSPs, such as DNAJB1, HSPA1A, HSPA5, HSPB1, HSPD1, and HSPH1, as well as BAG3 and clusterin (CLU), were up-regulated. |
| Experimental Parameters | Transcriptome Alterations | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Dataset | Cell Line and Origin | Technique/Platform | Heating Technique | Temp. (°C) | Heating Time (min) | CEM43 | Timepoint (Hours) | Total Transcripts | Upregulated (%) | Downregulated (%) | |
| Tabuchi, 2008 | Lymphoma (U937) | cDNA Microarray. Human Genome U133A array | Conductive heating: Water bath | 41 | 30 | 1875 | 3 | 12,815 | 2970 (23.18%) | 1708 (13.3%) | |
| Furusawa, 2011 (a) | Lymphoma (U937) | cDNA Microarray. Human Genome U133-plus 2.0 | Conductive heating: Water bath | 42 | 90 | 22.5 | 0 | 20,594 | 5466 (26.54%) | 5992 (29.10%) | |
| Furusawa, 2011 (b) | Lymphoma (U937) | cDNA Microarray. Human Genome U133A array | Conductive heating: Water bath | 42 | 15 | 3.75 | 1 | 20,594 | 4613 (22.40%) | 7199 (39.69%) | |
| Furusawa, 2011 (c) | Lymphoma (U937) | cDNA Microarray. Human Genome U133-plus 2.0 | Conductive heating: Water bath | 42 | 15 | 3.75 | 3 | 20,594 | 3084 (14.98%) | 13,003 (63.14%) | |
| Furusawa, 2011 (d) | Lymphoma (U937) | cDNA Microarray. Human Genome U133A array | Conductive heating: Water bath | 42 | 15 | 3.75 | 6 | 20,594 | 3772 (18.32%) | 11,589 (56.27%) | |
| Furusawa, 2011 (e) | Lymphoma (U937) | cDNA Microarray. Human Genome U133-plus 2.0 | Conductive heating: Water bath | 44 | 15 | 30 | 0 | 20,594 | 6008 (29.17%) | 4770 (23.16%) | |
| Furusawa, 2011 (f) | Lymphoma (U937) | cDNA Microarray. Human Genome U133A array | Conductive heating: Water bath | 44 | 15 | 30 | 1 | 20,594 | 8779 (42.63%) | 3190 (15.49%) | |
| Furusawa, 2011 (g) | Lymphoma (U937) | cDNA Microarray. Human Genome U133-plus 2.0 | Conductive heating: Water bath | 44 | 15 | 30 | 3 | 20,594 | 7611 (36.96%) | 5155 (25.03%) | |
| Furusawa, 2011 (h) | Lymphoma (U937) | cDNA Microarray. Human Genome U133A array | Conductive heating: Water bath | 44 | 15 | 30 | 6 | 20,594 | 7017 (34.07%) | 5155 (25.03%) | |
| Amaya, 2014 (a) | Breast cancer (MCF7) | cDNA Microarray. Illumina HumanHT-12 V4.0 | Conductive heating: Water bath | 45 | 30 | 120 | 4 | 20,909 | 1137 (5.44%) | 1190 (5.69%) | |
| Amaya, 2014 (b) | Breast cancer (MDA-MB-231) | cDNA Microarray. Illumina HumanHT-12 V4.0 beadchip | Conductive heating: Water bath | 45 | 30 | 120 | 4 | 20,909 | 1336 (6.39%) | 16,385 (78.36%) | |
| Amaya, 2014 (c) | Breast cancer (MDA-MB-468) | cDNA Microarray. Illumina Human HT-12 V4.0 beadchip | Conductive heating: Water bath | 45 | 30 | 120 | 4 | 20,909 | 132 (0.63%) | 1929 (9.23%) | |
| Court, 2017 | Ovarian cancer (HeyA8) | cDNA Microarray. Human HT-12 v4 Beadchip | Electromagnetic: Magnetic fluid hyperthermia | 43 | 30 | 30 | 0 | 18,299 | 0 (0%) | 1 (0.01%) | |
| Andocs, 2015 (a) | Colorectal cancer (HT-29) | cDNA Microarray. HGU133 Plus 2.0 arrays | Electromagnetic: Modulated electrohyperthermia | 42 | 30 | 7.5 | 4 | 12,372 | 8 (0.06%) | 1 (<0.01%) | |
| Andocs, 2015 (b) | Colorectal cancer (HT-29) | cDNA Microarray. HGU133 Plus 2.0 arrays | Electromagnetic: Modulated electrohyperthermia | 42 | 30 | 7.5 | 24 | 12,232 | 0 (0%) | 1 (<0.01%) | |
| Yunoki, 2016 | Oral squamous cell carcinoma (HSC-3) | cDNA Microarray. Human Genome U133-plus 2.0 array | Conductive heating: Water bath | 44 | 90 | 180 | 24 | 12,815 | 89 (0.69%) | 23 (0.18%) | |
| Scutigliani, 2022 (a) * | Bladder cancer (T24) | RNA sequencing. Illumina Novaseq 6000, paired-end, read depth of 40M. | Conductive heating: Water bath | 42 | 60 | 15 | 6 | 24,418 | 2370 (9.71%) | 2112 (8.65%) | |
| Scutigliani, 2022 (b) * | Cervical cancer (HeLa) | RNA sequencing. Illumina Novaseq 6000, paired-end, read depth of 40 M. | Conductive heating: Water bath | 42 | 60 | 15 | 24 | 17,673 | 71 (0.40%) | 126 (0.71%) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scutigliani, E.M.; Lobo-Cerna, F.; Mingo Barba, S.; Scheidegger, S.; Krawczyk, P.M. The Effects of Heat Stress on the Transcriptome of Human Cancer Cells: A Meta-Analysis. Cancers 2023, 15, 113. https://doi.org/10.3390/cancers15010113
Scutigliani EM, Lobo-Cerna F, Mingo Barba S, Scheidegger S, Krawczyk PM. The Effects of Heat Stress on the Transcriptome of Human Cancer Cells: A Meta-Analysis. Cancers. 2023; 15(1):113. https://doi.org/10.3390/cancers15010113
Chicago/Turabian StyleScutigliani, Enzo M., Fernando Lobo-Cerna, Sergio Mingo Barba, Stephan Scheidegger, and Przemek M. Krawczyk. 2023. "The Effects of Heat Stress on the Transcriptome of Human Cancer Cells: A Meta-Analysis" Cancers 15, no. 1: 113. https://doi.org/10.3390/cancers15010113