
Transcriptomic Profiling of Gene Expression Associated with Granulosa Cell Tumor Development in a Mouse Model

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement and Mice
2.2. RNA Isolation and Quantitative Reverse Transcription PCR (qRT-PCR)
2.3. PCR Profiler Array of Oncogenes and Tumor Suppressor Genes
2.4. RNA-Sequencing and Bioinformatics Analysis
2.5. Immunohistochemistry
2.6. Statistical Analysis
3. Results
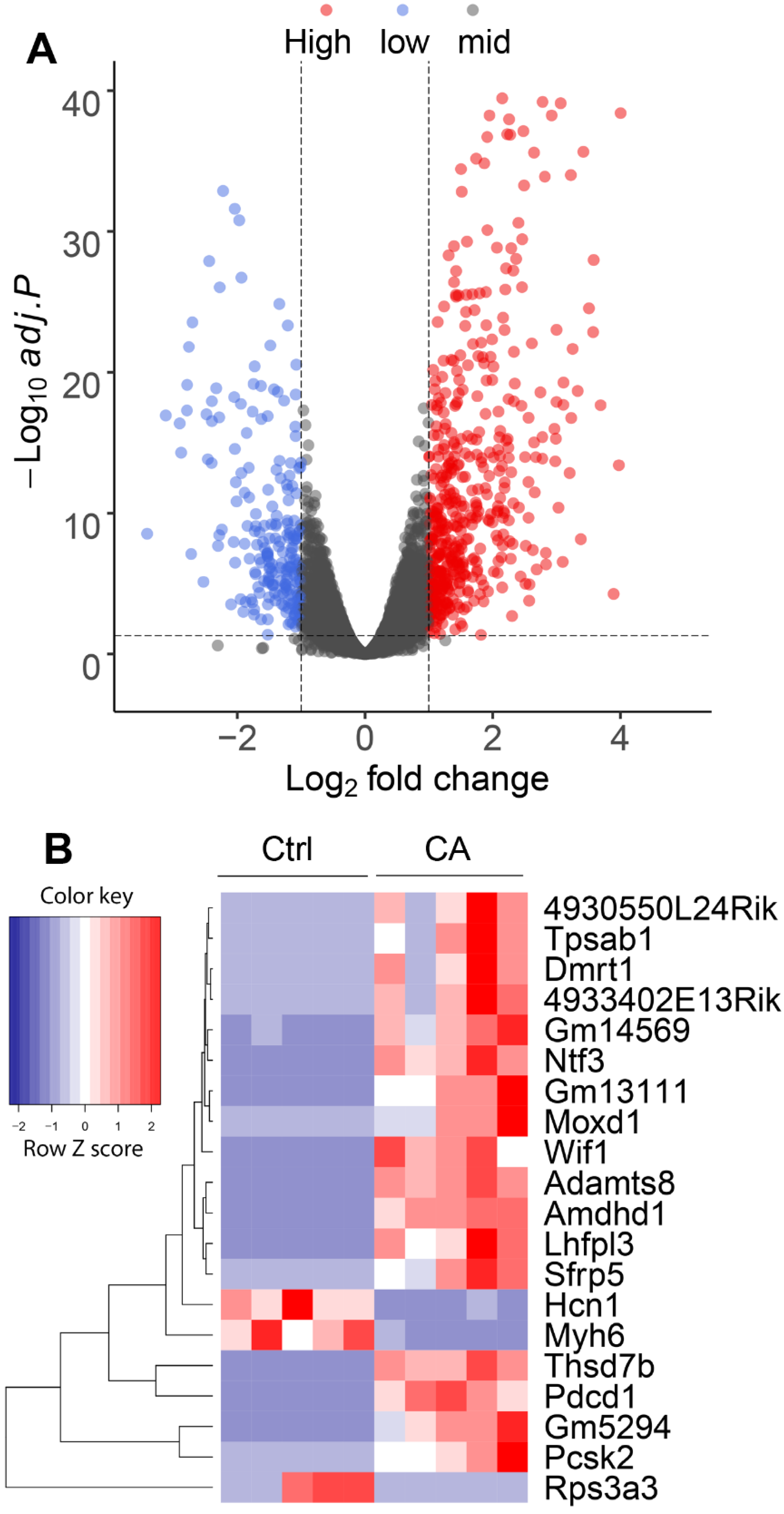
3.1. Transcriptomic Profiles of GCTs Harboring Constitutively Active TGFBR1
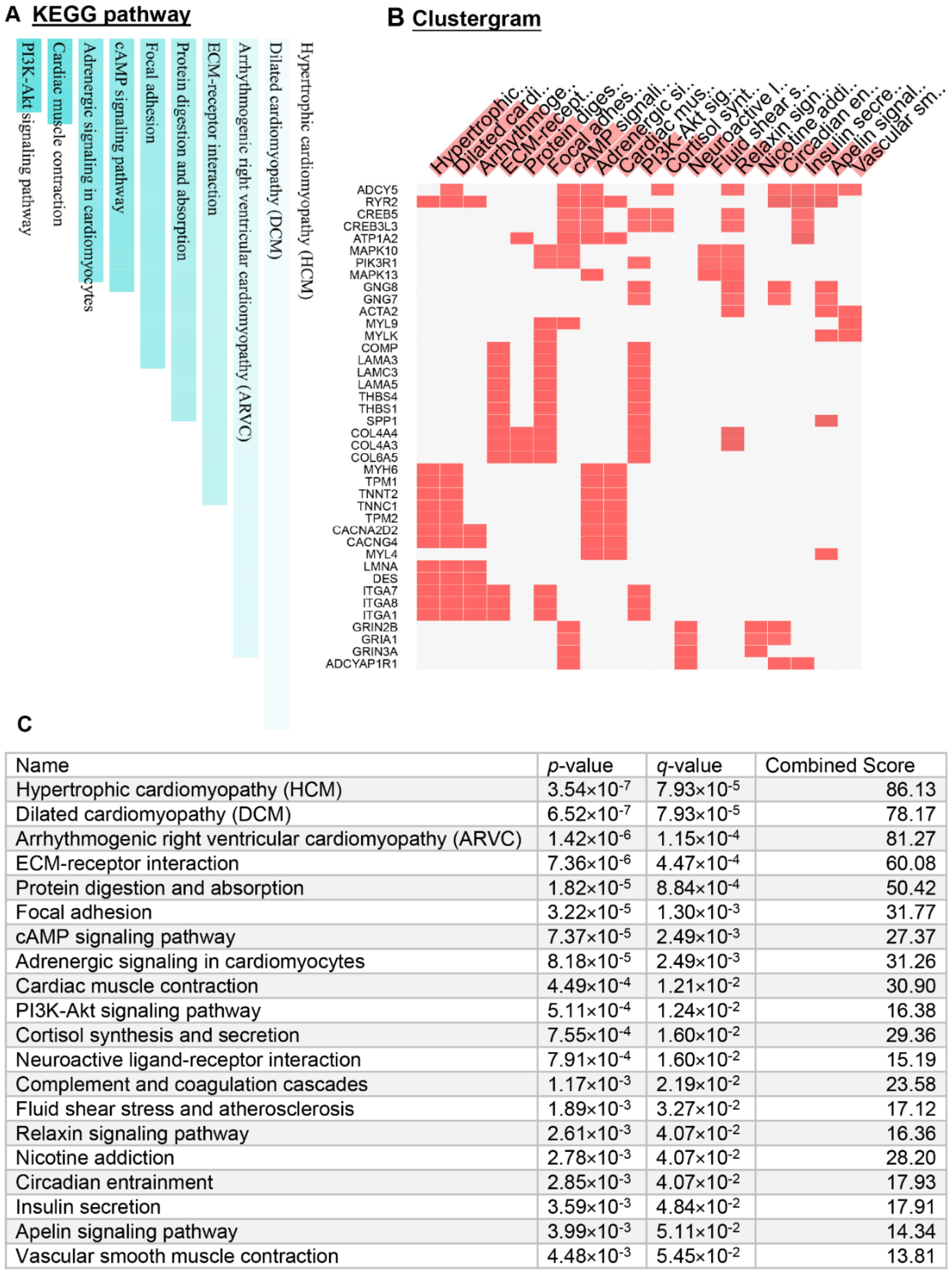
3.2. Identification of Signaling Pathways and Regulators Associated with GCT Development
3.3. Identification of Potential Clinically Relevant Regulators of Ovarian GCT Development
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pectasides, D.; Pectasides, E.; Psyrri, A. Granulosa cell tumor of the ovary. Cancer Treat. Rev. 2008, 34, 1–12. [Google Scholar] [CrossRef]
- Jamieson, S.; Fuller, P.J. Molecular pathogenesis of granulosa cell tumors of the ovary. Endocr. Rev. 2012, 33, 109–144. [Google Scholar] [CrossRef]
- Boerboom, D.; Paquet, M.; Hsieh, M.; Liu, J.; Jamin, S.P.; Behringer, R.R.; Sirois, J.; Taketo, M.M.; Richards, J.S. Misregulated wnt/beta-catenin signaling leads to ovarian granulosa cell tumor development. Cancer Res. 2005, 65, 9206–9215. [Google Scholar] [CrossRef] [Green Version]
- Pangas, S.A.; Li, X.; Umans, L.; Zwijsen, A.; Huylebroeck, D.; Gutierrez, C.; Wang, D.; Martin, J.F.; Jamin, S.P.; Behringer, R.R.; et al. Conditional deletion of smad1 and smad5 in somatic cells of male and female gonads leads to metastatic tumor development in mice. Mol. Cell. Biol. 2008, 28, 248–257. [Google Scholar] [CrossRef] [Green Version]
- Edson, M.A.; Nalam, R.L.; Clementi, C.; Franco, H.L.; Demayo, F.J.; Lyons, K.M.; Pangas, S.A.; Matzuk, M.M. Granulosa cell-expressed bmpr1a and bmpr1b have unique functions in regulating fertility but act redundantly to suppress ovarian tumor development. Mol. Endocrinol. 2010, 24, 1251–1266. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Ren, Y.A.; Pangas, S.A.; Adams, J.; Zhou, W.; Castrillon, D.H.; Wilhelm, D.; Richards, J.S. Foxo1/3 and pten depletion in granulosa cells promotes ovarian granulosa cell tumor development. Mol. Endocrinol. 2015, 29, 1006–1024. [Google Scholar] [CrossRef]
- Fang, X.; Gao, Y.; Li, Q. Smad3 activation: A converging point of dysregulated tgf-beta superfamily signaling and genetic aberrations in granulosa cell tumor development? Biol. Reprod. 2016, 95, 105. [Google Scholar] [CrossRef]
- Mansouri-Attia, N.; Tripurani, S.K.; Gokul, N.; Piard, H.; Anderson, M.L.; Eldin, K.; Pangas, S.A. Tgfbeta signaling promotes juvenile granulosa cell tumorigenesis by suppressing apoptosis. Mol. Endocrinol. 2014, 28, 1887–1898. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Vincent, D.F.; Davis, A.J.; Sansom, O.J.; Bartholin, L.; Li, Q. Constitutively active transforming growth factor β receptor 1 in the mouse ovary promotes tumorigenesis. Oncotarget 2016, 7, 40904–40918. [Google Scholar] [CrossRef] [Green Version]
- Shah, S.P.; Kobel, M.; Senz, J.; Morin, R.D.; Clarke, B.A.; Wiegand, K.C.; Leung, G.; Zayed, A.; Mehl, E.; Kalloger, S.E.; et al. Mutation of foxl2 in granulosa-cell tumors of the ovary. N. Engl. J. Med. 2009, 360, 2719–2729. [Google Scholar] [CrossRef]
- Secchi, C.; Benaglio, P.; Mulas, F.; Belli, M.; Stupack, D.; Shimasaki, S. FOXO1 mitigates the SMAD3/FOXL2c134w transcriptomic effect in a model of human adult granulosa cell tumor. J. Transl. Med. 2021, 19, 90. [Google Scholar] [CrossRef]
- Groeneweg, J.W.; Roze, J.F.; Peters, E.D.J.; Sereno, F.; Brink, A.G.J.; Paijens, S.T.; Nijman, H.W.; van Meurs, H.S.; van Lonkhuijzen, L.; Piek, J.M.J.; et al. FOXL2 and tert promoter mutation detection in circulating tumor DNA of adult granulosa cell tumors as biomarker for disease monitoring. Gynecol. Oncol. 2021, 162, 413–420. [Google Scholar] [CrossRef]
- Pierini, S.; Tanyi, J.L.; Simpkins, F.; George, E.; Uribe-Herranz, M.; Drapkin, R.; Burger, R.; Morgan, M.A.; Facciabene, A. Ovarian granulosa cell tumor characterization identifies FOXL2 as an immunotherapeutic target. JCI Insight 2020, 5, e136773. [Google Scholar] [CrossRef]
- Weis-Banke, S.E.; Lerdrup, M.; Kleine-Kohlbrecher, D.; Mohammad, F.; Sidoli, S.; Jensen, O.N.; Yanase, T.; Nakamura, T.; Iwase, A.; Stylianou, A.; et al. Mutant FOXL2c134w hijacks SMAD4 and SMAD2/3 to drive adult granulosa cell tumors. Cancer Res. 2020, 80, 3466–3479. [Google Scholar] [CrossRef]
- Gao, Y.; Wen, H.; Wang, C.; Li, Q. Smad7 antagonizes key tgfbeta superfamily signaling in mouse granulosa cells in vitro. Reproduction 2013, 146, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Spandidos, A.; Wang, H.; Seed, B. Primerbank: A pcr primer database for quantitative gene expression analysis, 2012 update. Nucleic Acids Res. 2012, 40, D1144–D1149. [Google Scholar] [CrossRef] [Green Version]
- Spandidos, A.; Wang, X.W.; Wang, H.J.; Seed, B. Primerbank: A resource of human and mouse pcr primer pairs for gene expression detection and quantification. Nucleic Acids Res. 2010, 38, D792–D799. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative pcr and the 2(-delta delta c(t)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Fang, X.; Ni, N.; Gao, Y.; Lydon, J.P.; Ivanov, I.; Rijnkels, M.; Bayless, K.J.; Li, Q. Transforming growth factor beta signaling and decidual integrity in mice. Biol. Reprod. 2020, 103, 1186–1198. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. Star: Ultrafast universal rna-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Putri, G.H.; Anders, S.; Pyl, P.T.; Pimanda, J.E.; Zanini, F. Analysing high-throughput sequencing data in python with htseq 2.0. arXiv 2021, arXiv:2112.00939. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. Edger: A bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Z.; Bailey, A.; Kuleshov, M.V.; Clarke, D.J.B.; Evangelista, J.E.; Jenkins, S.L.; Lachmann, A.; Wojciechowicz, M.L.; Kropiwnicki, E.; Jagodnik, K.M.; et al. Gene set knowledge discovery with enrichr. Curr. Protoc. 2021, 1, e90. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and collaborative html5 gene list enrichment analysis tool. BMC Bioinformat. 2013, 14, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexiadis, M.; Chu, S.; Leung, D.; Gould, J.A.; Jobling, T.; Fuller, P.J. Transcriptomic analysis of stage 1 versus advanced adult granulosa cell tumors. Oncotarget 2016, 7, 14207–14219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poddubskaya, E.V.; Baranova, M.P.; Allina, D.O.; Sekacheva, M.I.; Makovskaia, L.A.; Kamashev, D.E.; Suntsova, M.V.; Barbara, V.S.; Kochergina-Nikitskaya, I.N.; Aleshin, A.A. Personalized prescription of imatinib in recurrent granulosa cell tumor of the ovary: Case report. Cold Spring Harb. Mol. Case Stud. 2019, 5, a003434. [Google Scholar] [CrossRef]
- Ambartsumian, N.; Klingelhofer, J.; Grigorian, M. The multifaceted s100a4 protein in cancer and inflammation. Methods Mol. Biol. 2019, 1929, 339–365. [Google Scholar]
- Ruas, M.; Peters, G. The p16ink4a/cdkn2a tumor suppressor and its relatives. Biochim. Biophys. Acta 1998, 1378, F115–F177. [Google Scholar] [CrossRef]
- Arcellana-Panlilio, M.Y.; Egeler, R.M.; Ujack, E.; Magliocco, A.; Stuart, G.C.; Robbins, S.M.; Coppes, M.J. Evidence of a role for the ink4 family of cyclin-dependent kinase inhibitors in ovarian granulosa cell tumors. Genes Chromosomes Cancer 2002, 35, 176–181. [Google Scholar] [CrossRef]
- Wang, X.; Yang, X.; Zhang, C.; Wang, Y.; Cheng, T.; Duan, L.; Tong, Z.; Tan, S.; Zhang, H.; Saw, P.E.; et al. Tumor cell-intrinsic pd-1 receptor is a tumor suppressor and mediates resistance to pd-1 blockade therapy. Proc. Natl. Acad. Sci. USA 2020, 117, 6640–6650. [Google Scholar] [CrossRef]
- Demidyuk, I.V.; Shubin, A.V.; Gasanov, E.V.; Kurinov, A.M.; Demkin, V.V.; Vinogradova, T.V.; Zinovyeva, M.V.; Sass, A.V.; Zborovskaya, I.B.; Kostrov, S.V. Alterations in gene expression of proprotein convertases in human lung cancer have a limited number of scenarios. PLoS ONE 2013, 8, e55752. [Google Scholar] [CrossRef] [PubMed]
- Peng, Q.; Liu, L.; Pei, H.; Zhang, J.; Chen, M.; Zhai, X. A lhfpl3-as1/mir-580-3p/stat3 feedback loop promotes the malignancy in melanoma via activation of jak2/stat3 signaling. Mol Cancer Res. 2020, 18, 1724–1734. [Google Scholar] [CrossRef] [PubMed]
- Henke, E.; Nandigama, R.; Ergun, S. Extracellular matrix in the tumor microenvironment and its impact on cancer therapy. Front. Mol. Biosci. 2019, 6, 160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prevarskaya, N.; Ouadid-Ahidouch, H.; Skryma, R.; Shuba, Y. Remodelling of Ca2+ transport in cancer: How it contributes to cancer hallmarks? Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130097. [Google Scholar] [CrossRef] [Green Version]
- Izdebska, M.; Zielinska, W.; Halas-Wisniewska, M.; Grzanka, A. Involvement of actin and actin-binding proteins in carcinogenesis. Cells 2020, 9, 2245. [Google Scholar] [CrossRef]
- Rodat-Despoix, L.; Chamlali, M.; Ouadid-Ahidouch, H. Ion channels as key partners of cytoskeleton in cancer disease. Biochim. Biophys. Acta Rev. Cancer 2021, 1876, 188627. [Google Scholar] [CrossRef]
- Xu, S.; Xu, H.; Wang, W.; Li, S.; Li, H.; Li, T.; Zhang, W.; Yu, X.; Liu, L. The role of collagen in cancer: From bench to bedside. J. Transl. Med. 2019, 17, 309. [Google Scholar] [CrossRef] [Green Version]
- Kottarathil, V.D.; Antony, M.A.; Nair, I.R.; Pavithran, K. Recent advances in granulosa cell tumor ovary: A review. Indian J. Surg. Oncol. 2013, 4, 37–47. [Google Scholar] [CrossRef] [Green Version]
- Richards, J.S.; Fan, H.Y.; Liu, Z.; Tsoi, M.; Lague, M.N.; Boyer, A.; Boerboom, D. Either kras activation or pten loss similarly enhance the dominant-stable ctnnb1-induced genetic program to promote granulosa cell tumor development in the ovary and testis. Oncogene 2012, 31, 1504–1520. [Google Scholar] [CrossRef] [Green Version]
- Melgarejo da Rosa, M.; Clara Sampaio, M.; Virginia Cavalcanti Santos, R.; Sharjeel, M.; Araujo, C.; Galdino da Rocha Pitta, M.; Cristiny Pereira, M.; Jesus Barreto de Melo Rego, M. Unveiling the pathogenesis of perineural invasion from the perspective of neuroactive molecules. Biochem. Pharmacol. 2021, 188, 114547. [Google Scholar] [CrossRef] [PubMed]
- Przygodzka, P.; Soboska, K.; Sochacka, E.; Boncela, J. Neuromedin u: A small peptide in the big world of cancer. Cancers 2019, 11, 1312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afshar-Kharghan, V. The role of the complement system in cancer. J. Clin. Investig. 2017, 127, 780–789. [Google Scholar] [CrossRef] [Green Version]
- Gkrozou, F.; Pappa, C.; Tsonis, O.; Dimitriou, E.; Paschopoulos, M. Relaxin as a potential diagnostic biomarker for ovarian cancer—A prospective study. Eur. J. Obstet. Gynecol. Reprod. Biol. 2021, 260, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Djiogue, S.; Nwabo Kamdje, A.H.; Vecchio, L.; Kipanyula, M.J.; Farahna, M.; Aldebasi, Y.; Seke Etet, P.F. Insulin resistance and cancer: The role of insulin and igfs. Endocr. Relat. Cancer 2013, 20, R1–R17. [Google Scholar] [CrossRef] [Green Version]
- Dzialo, E.; Tkacz, K.; Blyszczuk, P. Crosstalk between the tgf-beta and wnt signalling pathways during cardiac fibrogenesis. Acta Biochim. Pol. 2018, 65, 341–349. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Corcoran, R.B.; Welsh, J.W.; Pennica, D.; Levine, A.J. Wisp-1 is a wnt-1- and beta-catenin-responsive oncogene. Genes Dev. 2000, 14, 585–595. [Google Scholar] [CrossRef]
- Desnoyers, L.; Arnott, D.; Pennica, D. Wisp-1 binds to decorin and biglycan. J. Biol. Chem. 2001, 276, 47599–47607. [Google Scholar] [CrossRef] [Green Version]
- Hwang, J.; Arafeh, R.; Seo, J.-H.; Baca, S.C.; Ludwig, M.; Arnoff, T.E.; Richter, C.; Bergom, H.E.; McSweeney, S.; Rennhack, J.P.; et al. Creb5 reprograms nuclear interactions to promote resistance to androgen receptor targeting therapies. bioRxiv 2021. [Google Scholar] [CrossRef]
- Kojima, T.; Shimazui, T.; Hinotsu, S.; Joraku, A.; Oikawa, T.; Kawai, K.; Horie, R.; Suzuki, H.; Nagashima, R.; Yoshikawa, K.; et al. Decreased expression of cxxc4 promotes a malignant phenotype in renal cell carcinoma by activating wnt signaling. Oncogene 2009, 28, 297–305. [Google Scholar] [CrossRef] [Green Version]
- Paluszczak, J.; Sarbak, J.; Kostrzewska-Poczekaj, M.; Kiwerska, K.; Jarmuz-Szymczak, M.; Grenman, R.; Mielcarek-Kuchta, D.; Baer-Dubowska, W. The negative regulators of wnt pathway-dach1, dkk1, and wif1 are methylated in oral and oropharyngeal cancer and wif1 methylation predicts shorter survival. Tumour Biol. 2015, 36, 2855–2861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Chatterjee, M.; Schmid, H.; Beck, S.; Gawaz, M. Cxcl14 as an emerging immune and inflammatory modulator. J. Inflamm. 2016, 13, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skinner, M.K.; Schmidt, M.; Savenkova, M.I.; Sadler-Riggleman, I.; Nilsson, E.E. Regulation of granulosa and theca cell transcriptomes during ovarian antral follicle development. Mol. Reprod. Dev. 2008, 75, 1457–1472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, B.; Zhou, M.; Wu, H.; Xiong, Z. Spp1 promotes ovarian cancer progression via integrin beta1/fak/akt signaling pathway. Onco Targets Ther. 2018, 11, 1333–1343. [Google Scholar] [CrossRef] [Green Version]
- Vaittinen, M.; Kolehmainen, M.; Schwab, U.; Uusitupa, M.; Pulkkinen, L. Microfibrillar-associated protein 5 is linked with markers of obesity-related extracellular matrix remodeling and inflammation. Nutr. Diabetes 2011, 1, e15. [Google Scholar] [CrossRef] [Green Version]
- Hsu, Y.L.; Hung, J.Y.; Lee, Y.L.; Chen, F.W.; Chang, K.F.; Chang, W.A.; Tsai, Y.M.; Chong, I.W.; Kuo, P.L. Identification of novel gene expression signature in lung adenocarcinoma by using next-generation sequencing data and bioinformatics analysis. Oncotarget 2017, 8, 104831–104854. [Google Scholar] [CrossRef] [Green Version]
- Martin, T.A.; Li, A.X.; Sanders, A.J.; Ye, L.; Frewer, K.; Hargest, R.; Jiang, W.G. Nupr1 and its potential role in cancer and pathological conditions (review). Int. J. Oncol. 2021, 58, 21. [Google Scholar] [CrossRef]
- Zhu, Z.; He, X.; Johnson, C.; Stoops, J.; Eaker, A.E.; Stoffer, D.S.; Bell, A.; Zarnegar, R.; DeFrances, M.C. Pi3k is negatively regulated by pik3ip1, a novel p110 interacting protein. Biochem. Biophys. Res. Commun. 2007, 358, 66–72. [Google Scholar] [CrossRef] [Green Version]
- Lague, M.N.; Paquet, M.; Fan, H.Y.; Kaartinen, M.J.; Chu, S.; Jamin, S.P.; Behringer, R.R.; Fuller, P.J.; Mitchell, A.; Dore, M.; et al. Synergistic effects of pten loss and wnt/ctnnb1 signaling pathway activation in ovarian granulosa cell tumor development and progression. Carcinogenesis 2008, 29, 2062–2072. [Google Scholar] [CrossRef] [Green Version]
- Lu, P.; Weaver, V.M.; Werb, Z. The extracellular matrix: A dynamic niche in cancer progression. J. Cell Biol. 2012, 196, 395–406. [Google Scholar] [CrossRef]
- Cho, A.; Howell, V.M.; Colvin, E.K. The extracellular matrix in epithelial ovarian cancer—A piece of a puzzle. Front. Oncol. 2015, 5, 245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, C.P.; Wu, B.H.; Chen, S.P.; Fu, M.Y.; Yang, M.; Liu, F.; Wang, B.Q. High COL4A3 expression correlates with poor prognosis after cisplatin plus gemcitabine chemotherapy in non-small cell lung cancer. Tumour Biol. 2013, 34, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Li, J.D.; Zeng, J.J.; Zou, X.P.; Tang, D.; Tang, W.; Rong, M.H.; Li, Y.; Dai, W.B.; Tang, Z.Q.; et al. Clinical value and potential mechanisms of COL8A1 upregulation in breast cancer: A comprehensive analysis. Cancer Cell Int. 2020, 20, 392. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Ge, G.; Izzi, V.; Greenspan, D.S. Alpha3 chains of type v collagen regulate breast tumour growth via glypican-1. Nat. Commun. 2017, 8, 14351. [Google Scholar] [CrossRef] [PubMed]
- Kimura, K.; Nakayama, M.; Naito, I.; Komiyama, T.; Ichimura, K.; Asano, H.; Tsukuda, K.; Ohtsuka, A.; Oohashi, T.; Miyoshi, S.; et al. Human collagen xv is a prominent histopathological component of sinusoidal capillarization in hepatocellular carcinogenesis. Int. J. Clin. Oncol. 2016, 21, 302–309. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Song, E.C.; Resnick, M.B. Elastin in the tumor microenvironment. Adv. Exp. Med. Biol. 2020, 1272, 1–16. [Google Scholar]
- Gordon-Weeks, A.; Lim, S.Y.; Yuzhalin, A.; Lucotti, S.; Vermeer, J.A.F.; Jones, K.; Chen, J.; Muschel, R.J. Tumour-derived laminin alpha5 (lama5) promotes colorectal liver metastasis growth, branching angiogenesis and notch pathway inhibition. Cancers 2019, 11, 630. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Q.; Ge, G. Lysyl oxidase, extracellular matrix remodeling and cancer metastasis. Cancer Microenviron. 2012, 5, 261–273. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.A.; Wagle, M.; Tran, K.; Zhan, X.; Dixon, M.A.; Liu, S.; Gros, D.; Korver, W.; Yonkovich, S.; Tomasevic, N.; et al. R-spondin family members regulate the wnt pathway by a common mechanism. Mol. Biol. Cell 2008, 19, 2588–2596. [Google Scholar] [CrossRef] [Green Version]
- De Cian, M.C.; Pauper, E.; Bandiera, R.; Vidal, V.P.; Sacco, S.; Gregoire, E.P.; Chassot, A.A.; Panzolini, C.; Wilhelm, D.; Pailhoux, E.; et al. Amplification of r-spondin1 signaling induces granulosa cell fate defects and cancers in mouse adult ovary. Oncogene 2017, 36, 208–218. [Google Scholar] [CrossRef] [Green Version]
- Pennica, D.; Swanson, T.A.; Welsh, J.W.; Roy, M.A.; Lawrence, D.A.; Lee, J.; Brush, J.; Taneyhill, L.A.; Deuel, B.; Lew, M.; et al. Wisp genes are members of the connective tissue growth factor family that are up-regulated in wnt-1-transformed cells and aberrantly expressed in human colon tumors. Proc. Natl. Acad. Sci. USA 1998, 95, 14717–14722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Rankin, S.A.; Sinner, D.; Kenny, A.P.; Krieg, P.A.; Zorn, A.M. Sfrp5 coordinates foregut specification and morphogenesis by antagonizing both canonical and noncanonical wnt11 signaling. Genes Dev. 2008, 22, 3050–3063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malinauskas, T.; Aricescu, A.R.; Lu, W.; Siebold, C.; Jones, E.Y. Modular mechanism of wnt signaling inhibition by wnt inhibitory factor 1. Nat. Struct. Mol. Biol. 2011, 18, 886–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daud, A.I.; Bumpus, F.M.; Husain, A. Characterization of angiotensin i-converting enzyme (ace)-containing follicles in the rat ovary during the estrous cycle and effects of ace inhibitor on ovulation. Endocrinology 1990, 126, 2927–2935. [Google Scholar] [CrossRef] [PubMed]
- Deshayes, F.; Nahmias, C. Angiotensin receptors: A new role in cancer? Trends Endocrinol. Metab. 2005, 16, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Beyazit, F.; Ayhan, S.; Celik, H.T.; Gungor, T. Assessment of serum angiotensin-converting enzyme in patients with epithelial ovarian cancer. Arch. Gynecol. Obstet. 2015, 292, 415–420. [Google Scholar] [CrossRef]
- Kong, D.H.; Kim, Y.K.; Kim, M.R.; Jang, J.H.; Lee, S. Emerging roles of vascular cell adhesion molecule-1 (vcam-1) in immunological disorders and cancer. Int. J. Mol. Sci. 2018, 19, 1057. [Google Scholar] [CrossRef] [Green Version]
- Farooqi, A.A.; Siddik, Z.H. Platelet-derived growth factor (pdgf) signalling in cancer: Rapidly emerging signalling landscape. Cell Biochem. Funct. 2015, 33, 257–265. [Google Scholar] [CrossRef] [Green Version]
- Kalin, R.E.; Kretz, M.P.; Meyer, A.M.; Kispert, A.; Heppner, F.L.; Brandli, A.W. Paracrine and autocrine mechanisms of apelin signaling govern embryonic and tumor angiogenesis. Dev. Biol. 2007, 305, 599–614. [Google Scholar] [CrossRef] [Green Version]
- Helfman, D.M.; Flynn, P.; Khan, P.; Saeed, A. Tropomyosin as a regulator of cancer cell transformation. Adv. Exp. Med. Biol. 2008, 644, 124–131. [Google Scholar]
- Leung, C.S.; Yeung, T.L.; Yip, K.P.; Pradeep, S.; Balasubramanian, L.; Liu, J.; Wong, K.K.; Mangala, L.S.; Armaiz-Pena, G.N.; Lopez-Berestein, G.; et al. Calcium-dependent fak/creb/tnnc1 signalling mediates the effect of stromal mfap5 on ovarian cancer metastatic potential. Nat. Commun. 2014, 5, 5092. [Google Scholar] [CrossRef] [PubMed]
- Dang, C.V. Links between metabolism and cancer. Genes Dev. 2012, 26, 877–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassanein, M.; Hoeksema, M.D.; Shiota, M.; Qian, J.; Harris, B.K.; Chen, H.; Clark, J.E.; Alborn, W.E.; Eisenberg, R.; Massion, P.P. Slc1a5 mediates glutamine transport required for lung cancer cell growth and survival. Clin. Cancer Res. 2013, 19, 560–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Ansari, R.; Alfarsi, L.; Craze, M.L.; Masisi, B.K.; Ellis, I.O.; Rakha, E.A.; Green, A.R. The solute carrier SLC7A8 is a marker of favourable prognosis in er-positive low proliferative invasive breast cancer. Breast Cancer Res. Treat 2020, 181, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Sherwood, O.D. Relaxin’s physiological roles and other diverse actions. Endocr. Rev. 2004, 25, 205–234. [Google Scholar] [CrossRef] [Green Version]
- Feng, S.; Agoulnik, I.U.; Bogatcheva, N.V.; Kamat, A.A.; Kwabi-Addo, B.; Li, R.; Ayala, G.; Ittmann, M.M.; Agoulnik, A.I. Relaxin promotes prostate cancer progression. Clin Cancer Res. 2007, 13, 1695–1702. [Google Scholar] [CrossRef] [Green Version]
- Fue, M.; Miki, Y.; Takagi, K.; Hashimoto, C.; Yaegashi, N.; Suzuki, T.; Ito, K. Relaxin 2/rxfp1 signaling induces cell invasion via the beta-catenin pathway in endometrial cancer. Int. J. Mol. Sci. 2018, 19, 2438. [Google Scholar] [CrossRef] [Green Version]
- Ng, H.H.; Shen, M.; Samuel, C.S.; Schlossmann, J.; Bennett, R.G. Relaxin and extracellular matrix remodeling: Mechanisms and signaling pathways. Mol. Cell. Endocrinol. 2019, 487, 59–65. [Google Scholar] [CrossRef]
- Mardhian, D.F.; Storm, G.; Bansal, R.; Prakash, J. Nano-targeted relaxin impairs fibrosis and tumor growth in pancreatic cancer and improves the efficacy of gemcitabine in vivo. J. Control. Release 2018, 290, 1–10. [Google Scholar] [CrossRef]
- Chang, L.; Chiang, S.H.; Saltiel, A.R. Insulin signaling and the regulation of glucose transport. Mol. Med. 2004, 10, 65–71. [Google Scholar] [CrossRef]
- Ray, A.; Alalem, M.; Ray, B.K. Insulin signaling network in cancer. Indian J. Biochem. Biophys. 2014, 51, 493–498. [Google Scholar] [PubMed]
- Hodson, D.J.; Mitchell, R.K.; Marselli, L.; Pullen, T.J.; Gimeno Brias, S.; Semplici, F.; Everett, K.L.; Cooper, D.M.; Bugliani, M.; Marchetti, P.; et al. Adcy5 couples glucose to insulin secretion in human islets. Diabetes 2014, 63, 3009–3021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Revel, M.; Daugan, M.V.; Sautes-Fridman, C.; Fridman, W.H.; Roumenina, L.T. Complement system: Promoter or suppressor of cancer progression? Antibodies 2020, 9, 57. [Google Scholar] [CrossRef]
- Cho, K. Emerging roles of complement protein c1q in neurodegeneration. Aging Dis. 2019, 10, 652–663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Wang, Y.; Yu, L.; Sakakura, K.; Visus, C.; Schwab, J.H.; Ferrone, C.R.; Favoino, E.; Koya, Y.; Campoli, M.R.; et al. Cspg4 in cancer: Multiple roles. Curr. Mol. Med. 2010, 10, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.R.; Dong, J.; Li, L. Cancer-associated fibroblasts-derived gamma-glutamyltransferase 5 promotes tumor growth and drug resistance in lung adenocarcinoma. Aging 2020, 12, 13220–13233. [Google Scholar] [CrossRef] [PubMed]
- Sercu, S.; Zhang, L.; Merregaert, J. The extracellular matrix protein 1: Its molecular interaction and implication in tumor progression. Cancer Investig. 2008, 26, 375–384. [Google Scholar] [CrossRef]
- Chun, M.G.; Hanahan, D. Genetic deletion of the desmosomal component desmoplakin promotes tumor microinvasion in a mouse model of pancreatic neuroendocrine carcinogenesis. PLoS Genet. 2010, 6, e1001120. [Google Scholar] [CrossRef] [Green Version]
- Pommier, R.M.; Gout, J.; Vincent, D.F.; Cano, C.E.; Kaniewski, B.; Martel, S.; Rodriguez, J.; Fourel, G.; Valcourt, U.; Marie, J.C.; et al. The human NUPR1/P8 gene is transcriptionally activated by transforming growth factor beta via the smad signalling pathway. Biochem. J. 2012, 445, 285–293. [Google Scholar] [CrossRef]
- Elsafadi, M.; Manikandan, M.; Dawud, R.A.; Alajez, N.M.; Hamam, R.; Alfayez, M.; Kassem, M.; Aldahmash, A.; Mahmood, A. Transgelin is a tgfbeta-inducible gene that regulates osteoblastic and adipogenic differentiation of human skeletal stem cells through actin cytoskeleston organization. Cell Death Dis. 2016, 7, e2321. [Google Scholar] [CrossRef] [Green Version]
- Elsafadi, M.; Manikandan, M.; Almalki, S.; Mahmood, A.; Shinwari, T.; Vishnubalaji, R.; Mobarak, M.; Alfayez, M.; Aldahmash, A.; Kassem, M.; et al. Transgelin is a poor prognostic factor associated with advanced colorectal cancer (crc) stage promoting tumor growth and migration in a tgf beta-dependent manner. Cell Death Dis. 2020, 11, 341. [Google Scholar] [CrossRef] [PubMed]
- Chavez, R.D.; Coricor, G.; Perez, J.; Seo, H.S.; Serra, R. Sox9 protein is stabilized by tgf-beta and regulates papss2 mrna expression in chondrocytes. Osteoarthr. Cartil. 2017, 25, 332–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Zou, X.; Qian, W.; Weng, X.; Zhang, L.; Zhang, L.; Wang, S.; Cao, X.; Ma, L.; Wei, G.; et al. Enhanced papss2/vcan sulfation axis is essential for snail-mediated breast cancer cell migration and metastasis. Cell Death Differ. 2019, 26, 565–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-Ze’ev, A.; Amsterdam, A. In vitro regulation of granulosa cell differentiation. Involvement of cytoskeletal protein expression. J. Biol. Chem. 1987, 262, 5366–5376. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ni, N.; Fang, X.; Mullens, D.A.; Cai, J.J.; Ivanov, I.; Bartholin, L.; Li, Q. Transcriptomic Profiling of Gene Expression Associated with Granulosa Cell Tumor Development in a Mouse Model. Cancers 2022, 14, 2184. https://doi.org/10.3390/cancers14092184
Ni N, Fang X, Mullens DA, Cai JJ, Ivanov I, Bartholin L, Li Q. Transcriptomic Profiling of Gene Expression Associated with Granulosa Cell Tumor Development in a Mouse Model. Cancers. 2022; 14(9):2184. https://doi.org/10.3390/cancers14092184
Chicago/Turabian StyleNi, Nan, Xin Fang, Destiny A. Mullens, James J. Cai, Ivan Ivanov, Laurent Bartholin, and Qinglei Li. 2022. "Transcriptomic Profiling of Gene Expression Associated with Granulosa Cell Tumor Development in a Mouse Model" Cancers 14, no. 9: 2184. https://doi.org/10.3390/cancers14092184