
Systematic Investigation of the Multifaceted Role of SOX11 in Cancer

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Processing and SOX11 Expression Analysis
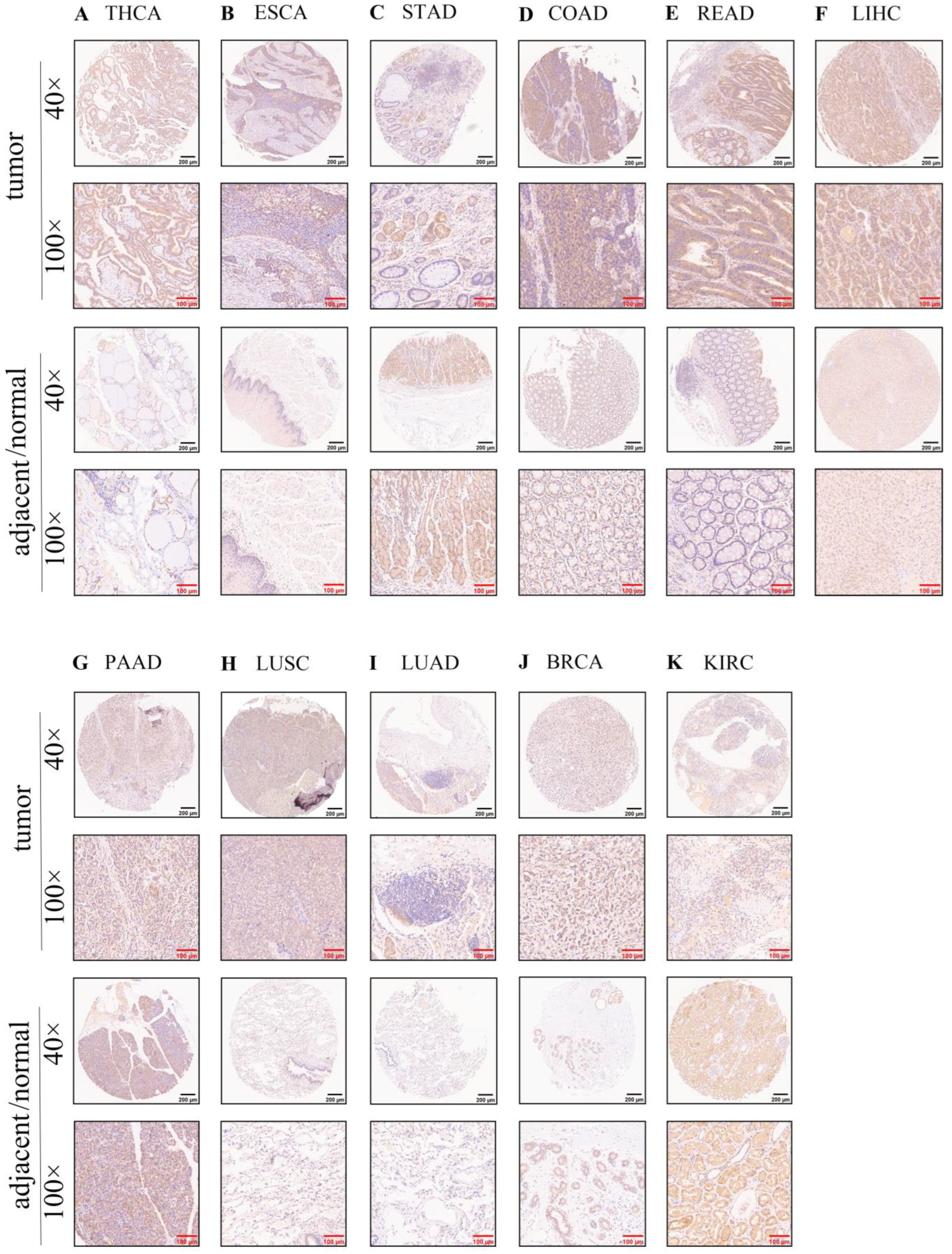
2.2. Tissue Microarray and Human Protein Atlas (HPA) Database
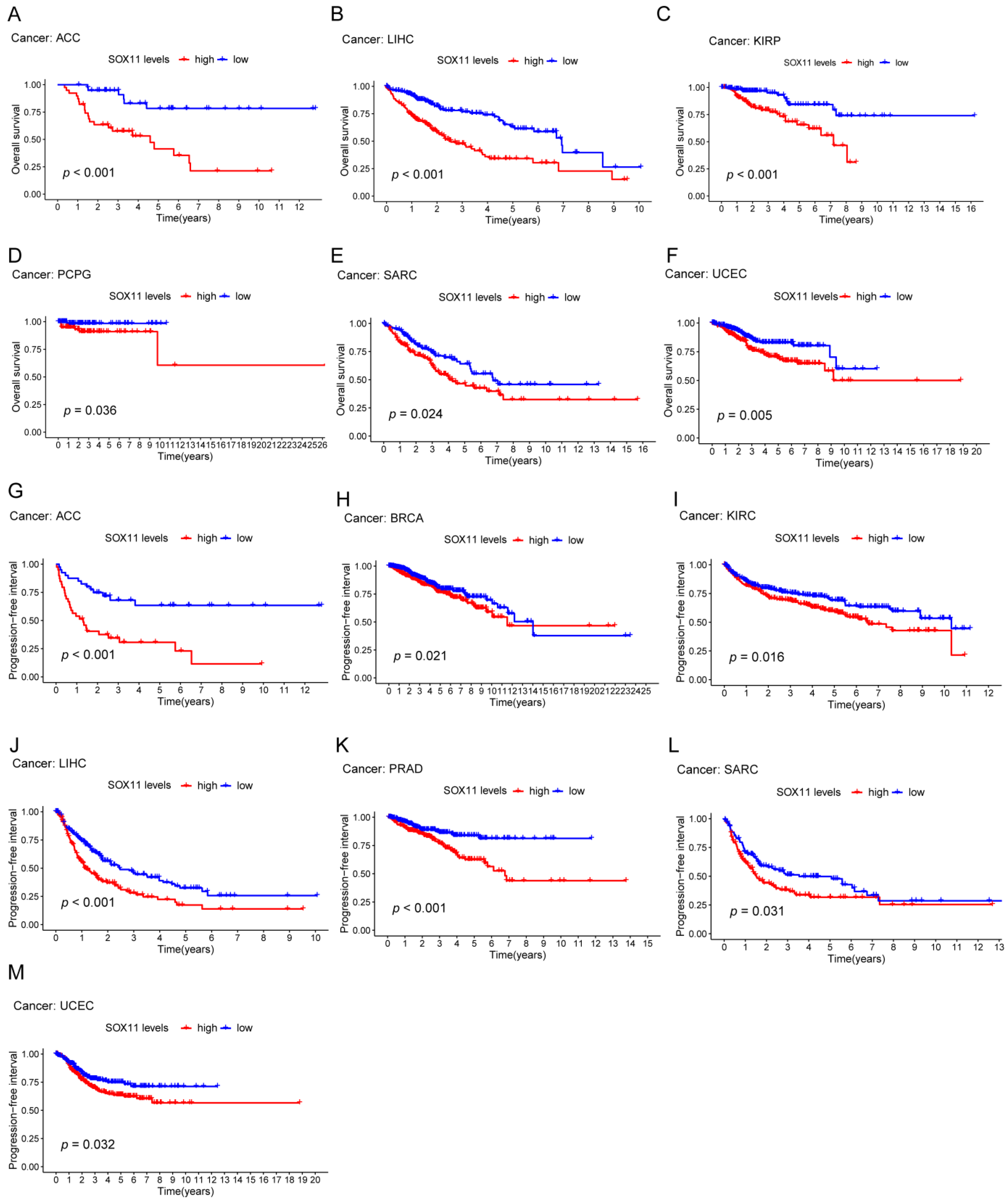
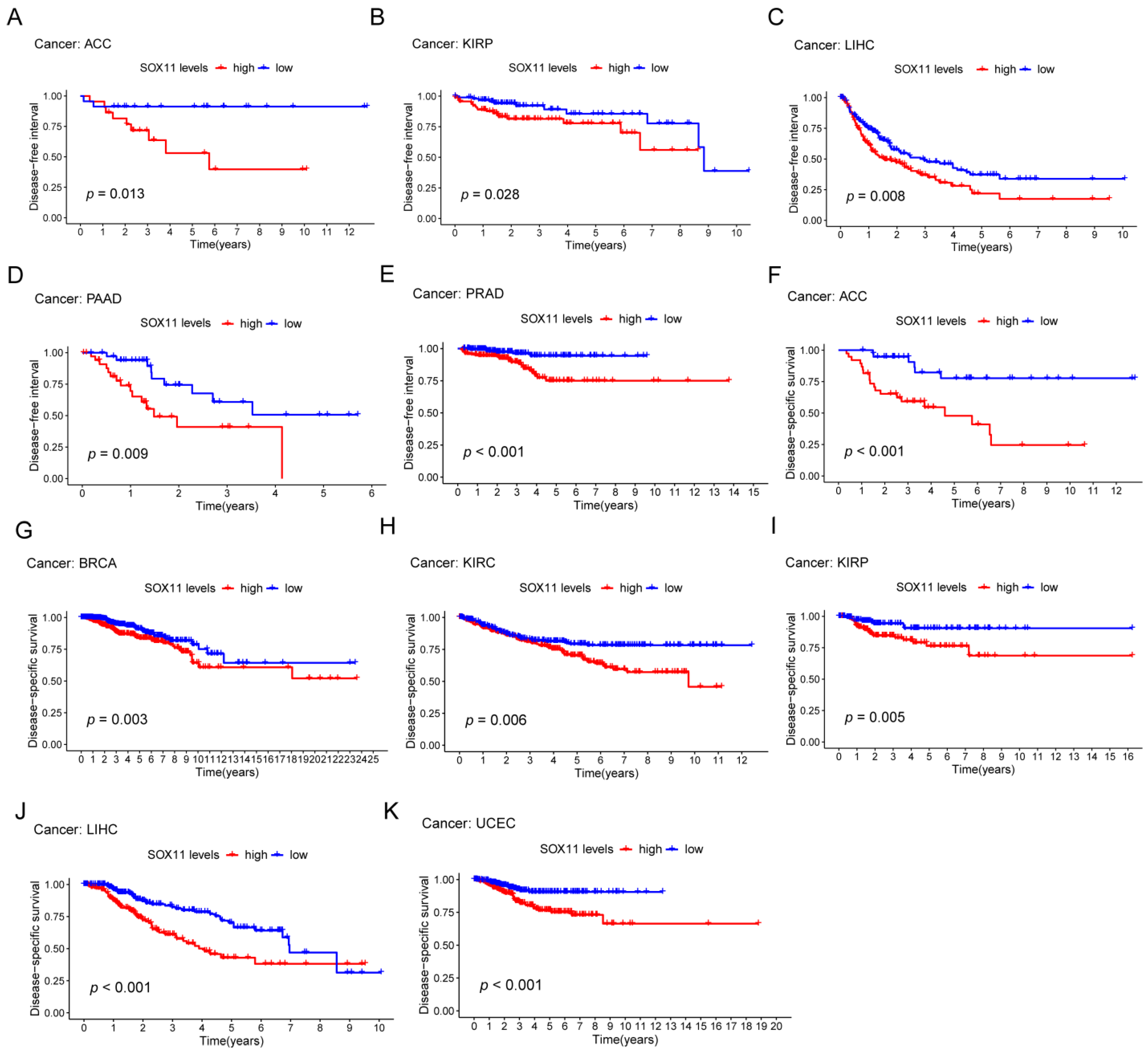
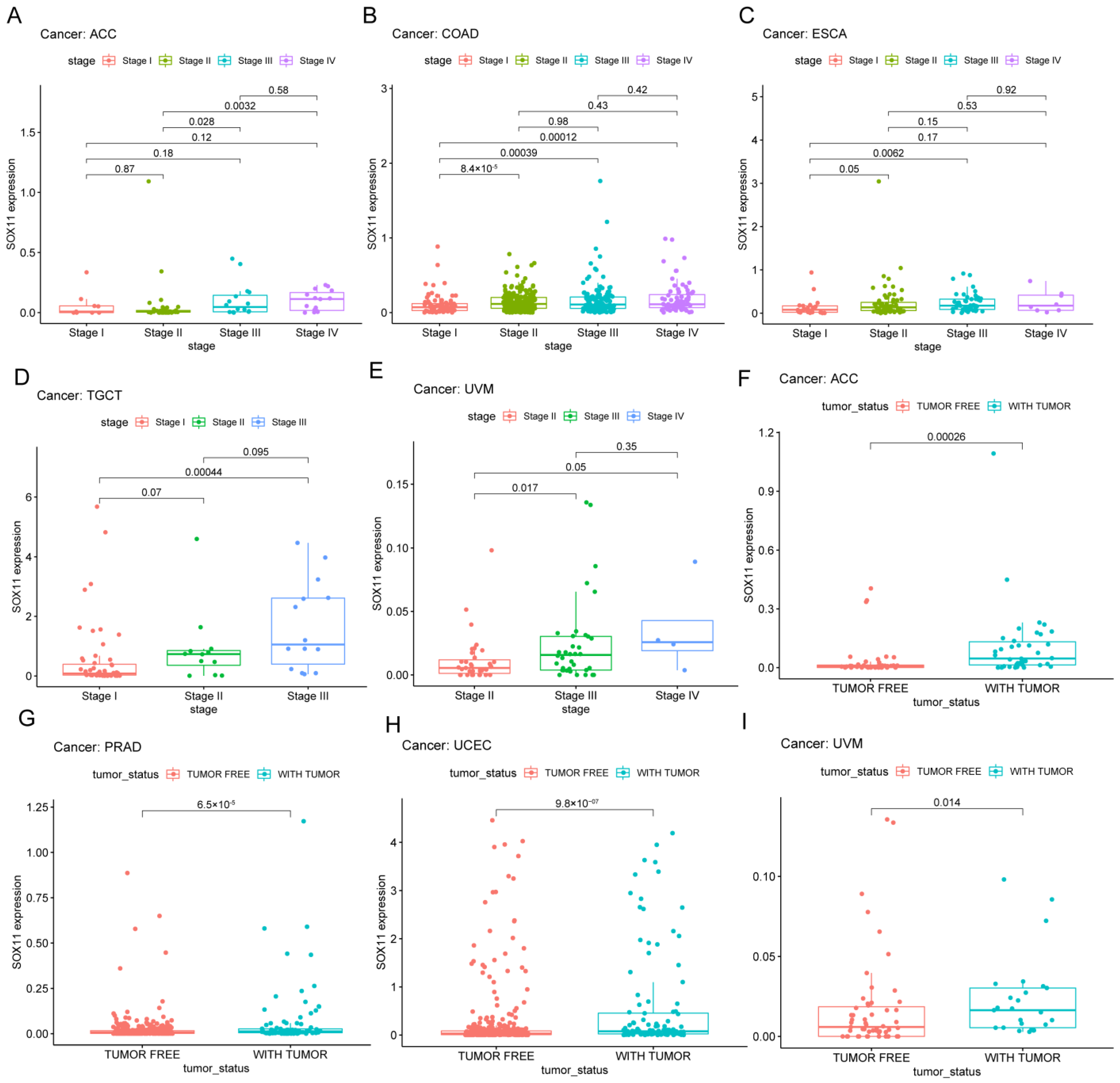
2.3. Relationship of SOX11 Expression with Survival Outcomes and Clinical Characteristics
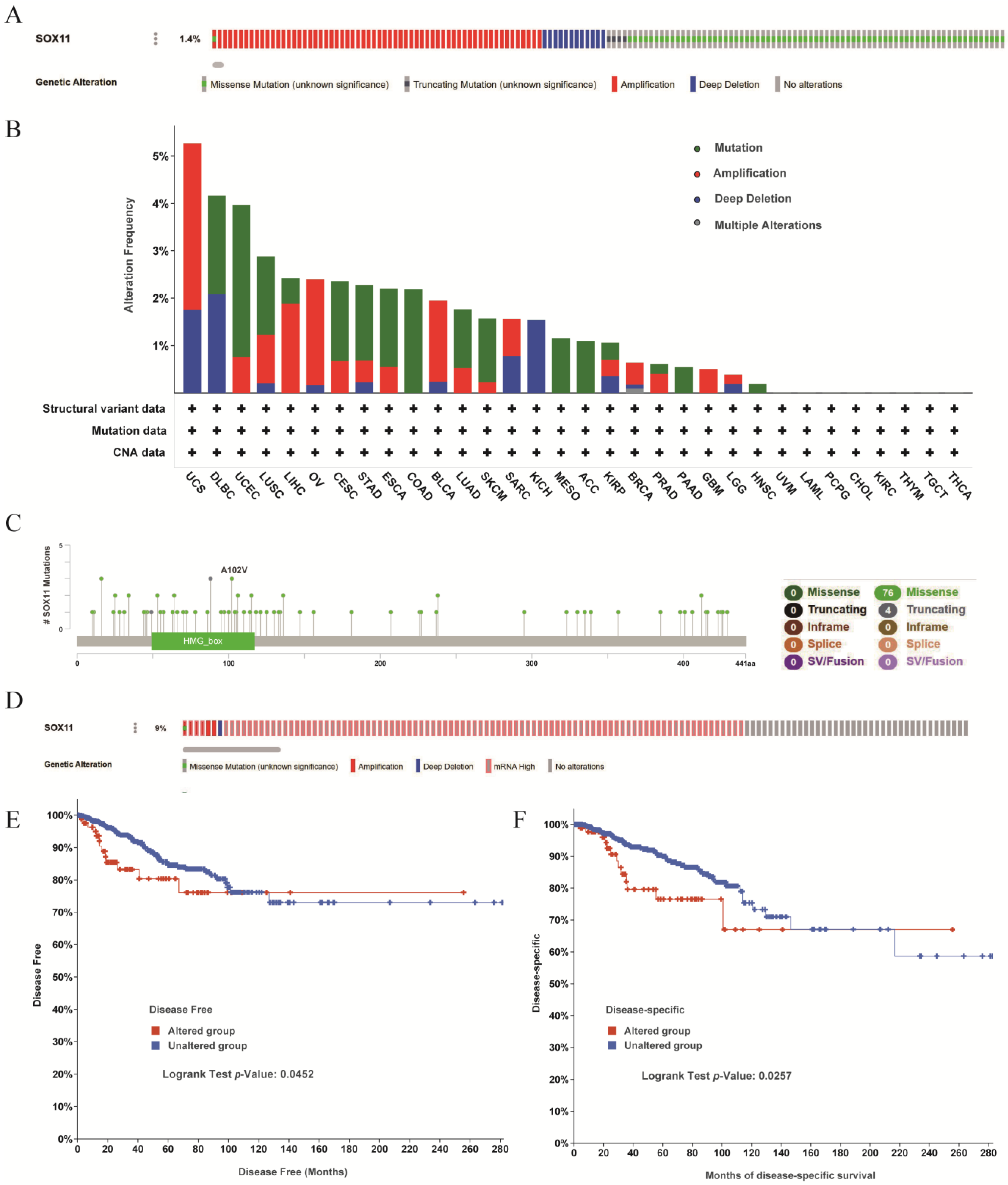
2.4. Genomic Alterations of SOX11 in Pan-Cancer and Breast Cancer
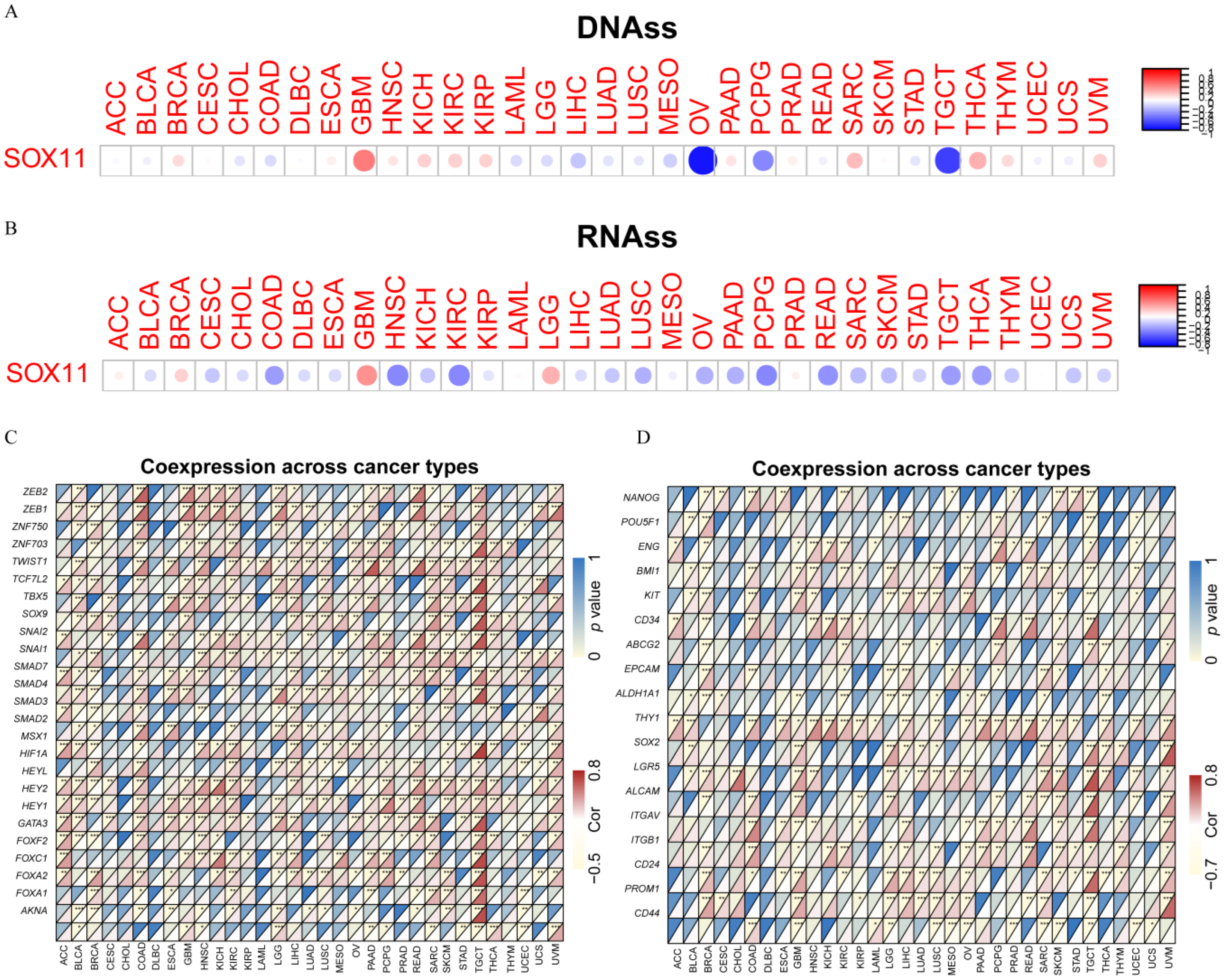
2.5. Analysis of SOX11 Expression with Stemness Score and EMT-Related Genes in Cancers
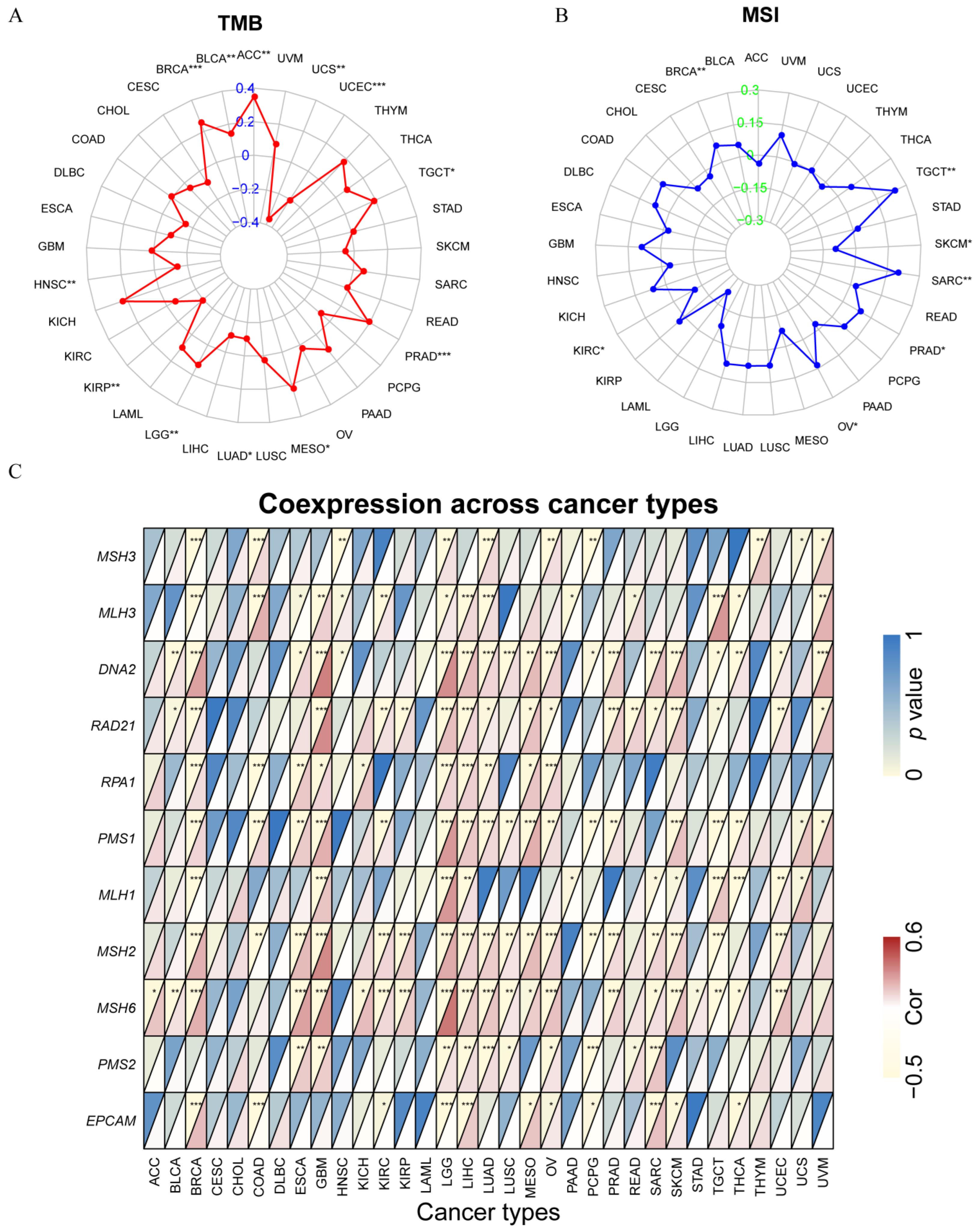
2.6. Analysis of SOX11 Expression with TMB and MSI in Pan-Cancer
2.7. Relationship of SOX11 Expression with the Tumor Immune Microenvironment in Pan-Cancer
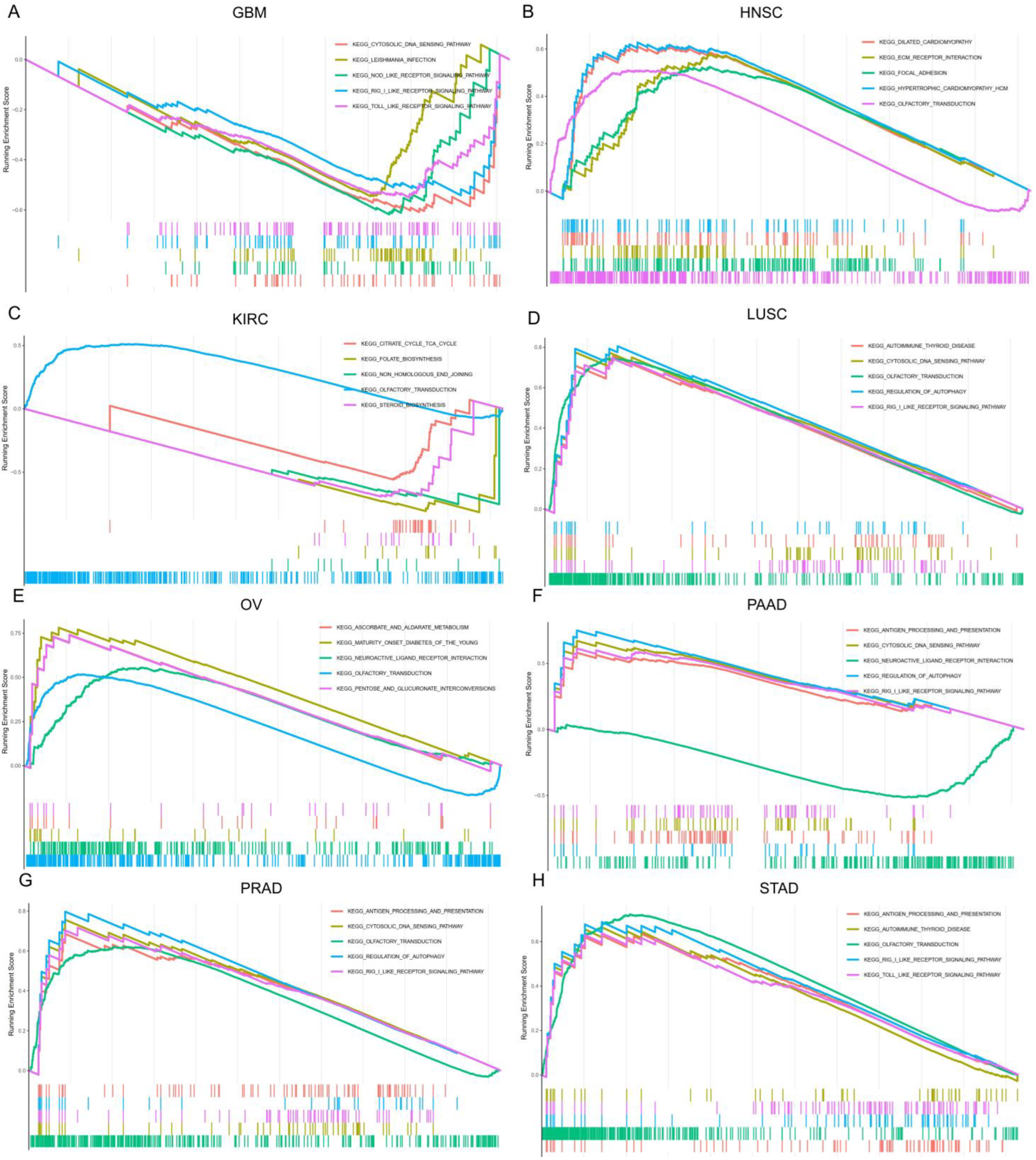
2.8. Gene Set Enrichment Analysis (GSEA)
2.9. Statistical Analysis
3. Results
3.1. Expression Levels of SOX11 in Normal Tissues and Pan-Cancer
3.2. Protein Expression of the SOX11 in Human Tissues
3.3. Prognostic Value of SOX11 in Pan-Cancer
3.4. Associations between the SOX11 Expression with Clinicopathological Features in Pan-Cancer
3.5. Analysis of Genetic Alterations in the SOX11 Gene of Pan-Cancer Patients
3.6. Relationship between the SOX11 Expression and EMT-Related Genes and Stemness Scores in Pan-Cancer
3.7. Correlation between SOX11 Expression and MSI, TMB and MMR-Related Genes in Cancer
3.8. Relevance of SOX11 Expression to the Tumor Immune Microenvironment
3.9. The Biological Function of SOX11 in Pan-Cancer
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Xia, C.; Dong, X.; Li, H.; Cao, M.; Sun, D.; He, S.; Yang, F.; Yan, X.; Zhang, S.; Li, N.; et al. Cancer statistics in China and United States, 2022: Profiles, trends, and determinants. Chin. Med. J. 2022, 135, 584–590. [Google Scholar] [CrossRef] [PubMed]
- Grimm, D.; Bauer, J.; Wise, P.; Kruger, M.; Simonsen, U.; Wehland, M.; Infanger, M.; Corydon, T.J. The role of SOX family members in solid tumours and metastasis. Semin. Cancer Biol. 2020, 67, 122–153. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre, V.; Dumitriu, B.; Penzo-Méndez, A.; Han, Y.; Pallavi, B. Control of cell fate and differentiation by Sry-related high-mobility-group box (SOX) transcription factors. Int. J. Biochem. Cell Biol. 2007, 39, 2195–2214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cizelsky, W.; Hempel, A.; Metzig, M.; Tao, S.; Hollemann, T.; Kühl, M.; Kühl, S.J. SOX4 and SOX11 function during Xenopus laevis eye development. PLoS ONE 2013, 8, e69372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowles, J.; Schepers, G.; Koopman, P. Phylogeny of the SOX family of developmental transcription factors based on sequence and structural indicators. Dev. Biol. 2000, 227, 239–255. [Google Scholar] [CrossRef] [Green Version]
- Hoser, M.; Potzner, M.R.; Koch, J.M.; Bösl, M.R.; Wegner, M.; Sock, E. SOX12 deletion in the mouse reveals nonreciprocal redundancy with the related SOX4 and SOX11 transcription factors. Mol. Cell. Biol. 2008, 28, 4675–4687. [Google Scholar] [CrossRef] [Green Version]
- Tsang, S.M.; Oliemuller, E.; Howard, B.A. Regulatory roles for SOX11 in development, stem cells and cancer. Semin. Cancer Biol. 2020, 67, 3–11. [Google Scholar] [CrossRef]
- Jiang, Y.; Ding, Q.; Xie, X.; Libby, R.T.; Lefebvre, V.; Gan, L. Transcription factors SOX4 and SOX11 function redundantly to regulate the development of mouse retinal ganglion cells. J. Biol. Chem. 2013, 288, 18429–18438. [Google Scholar] [CrossRef] [Green Version]
- Kamachi, Y.; Kondoh, H. SOX proteins: Regulators of cell fate specification and differentiation. Development 2013, 140, 4129–4144. [Google Scholar] [CrossRef] [Green Version]
- Wasik, A.M.; Lord, M.; Wang, X.; Zong, F.; Andersson, P.; Kimby, E.; Christensson, B.; Karimi, M.; Sander, B. SOXC transcription factors in mantle cell lymphoma: The role of promoter methylation in SOX11 expression. Sci. Rep. 2013, 3, 1400. [Google Scholar] [CrossRef]
- Vegliante, M.C.; Royo, C.; Palomero, J.; Salaverria, I.; Balint, B.; Martín-Guerrero, I.; Agirre, X.; Lujambio, A.; Richter, J.; Xargay-Torrent, S.; et al. Epigenetic activation of SOX11 in lymphoid neoplasms by histone modifications. PLoS ONE 2011, 6, e21382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potzner, M.R.; Tsarovina, K.; Binder, E.; Penzo-Méndez, A.; Lefebvre, V.; Rohrer, H.; Wegner, M.; Sock, E. Sequential requirement of SOX4 and SOX11 during development of the sympathetic nervous system. Development 2010, 137, 775–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sock, E.; Rettig, S.D.; Enderich, J.; Bösl, M.R.; Tamm, E.R.; Wegner, M. Gene targeting reveals a widespread role for the high-mobility-group transcription factor SOX11 in tissue remodeling. Mol. Cell. Biol. 2004, 24, 6635–6644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pamnani, M.; Sinha, P.; Nara, S.; Sachan, M. Study of promoter DNA methylation of SOX11 and its correlation with tissue-specific expression in the laboratory mouse. Gene 2014, 552, 133–139. [Google Scholar] [CrossRef]
- Yang, Z.; Jiang, S.; Lu, C.; Ji, T.; Yang, W.; Li, T.; Lv, J.; Hu, W.; Yang, Y.; Jin, Z. SOX11: Friend or foe in tumor prevention and carcinogenesis? Ther. Adv. Med. Oncol. 2019, 11, 1758835919853449. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Zhou, H.; Zhang, Y.; Chen, J.; Han, X.; Huang, D.; Ren, X.; Jia, Y.; Fan, Q.; Tian, W.; et al. Aberrant methylation of FAT4 and SOX11 in peripheral blood leukocytes and their association with gastric cancer risk. J. Cancer 2018, 9, 2275–2283. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.W.; Ao, X.; Fu, S.M. Expression of SOX11 and HER2 and their association with recurrent breast cancer. Transl. Cancer Res. 2019, 8, 248–254. [Google Scholar] [CrossRef]
- He, J.X.; Xi, Y.F.; Su, L.P.; Gao, N.; Xu, E.W.; Xie, L.W.; Wang, L.Y.; Zheng, Y.P.; Han, W.E.; Chang, J.; et al. Association of SOX11 gene expression withclinical features and prognosis of mantle cell lymphoma. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 2556–2563. [Google Scholar] [CrossRef]
- Brennan, D.J.; Ek, S.; Doyle, E.; Drew, T.; Foley, M.; Flannelly, G.; O’Connor, D.P.; Gallagher, W.M.; Kilpinen, S.; Kallioniemi, O.P.; et al. The transcription factor SOX11 is a prognostic factor for improved recurrence-free survival in epithelial ovarian cancer. Eur. J. Cancer 2009, 45, 1510–1517. [Google Scholar] [CrossRef]
- Teng, Y.; Fan, Y.C.; Mu, N.N.; Zhao, J.; Sun, F.K.; Wang, K. Serum SOX11 promoter methylation is a novel biomarker for the diagnosis of Hepatitis B virus-related hepatocellular carcinoma. Neoplasma 2016, 63, 419–426. [Google Scholar] [CrossRef]
- Zhang, S.; Li, S.; Gao, J.L. Promoter methylation status of the tumor suppressor gene SOX11 is associated with cell growth and invasion in nasopharyngeal carcinoma. Cancer Cell Int. 2013, 13, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, Z.; Sun, B.; Hong, Q.; Yan, J.; Mu, D.; Li, J.; Sheng, H.; Guo, H. The role of tumor suppressor gene SOX11 in prostate cancer. Tumour Biol. 2015, 36, 6133–6138. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Chang, X.; Li, Z.; Wang, J.; Deng, P.; Zhu, X.; Liu, J.; Zhang, C.; Chen, S.; Dai, D. Aberrant SOX11 promoter methylation is associated with poor prognosis in gastric cancer. Cell. Oncol. 2015, 38, 183–194. [Google Scholar] [CrossRef]
- Chung, W.; Bondaruk, J.; Jelinek, J.; Lotan, Y.; Liang, S.; Czerniak, B.; Issa, J.P. Detection of bladder cancer using novel DNA methylation biomarkers in urine sediments. Cancer Epidemiol. Biomark. Prev. 2011, 20, 1483–1491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beekman, R.; Amador, V.; Campo, E. SOX11, a key oncogenic factor in mantle cell lymphoma. Curr. Opin. Hematol. 2018, 25, 299–306. [Google Scholar] [CrossRef]
- Zvelebil, M.; Oliemuller, E.; Gao, Q.; Wansbury, O.; Mackay, A.; Kendrick, H.; Smalley, M.J.; Reis-Filho, J.S.; Howard, B.A. Embryonic mammary signature subsets are activated in Brca1−/− and basal-like breast cancers. Breast Cancer Res. 2013, 15, R25. [Google Scholar] [CrossRef] [Green Version]
- Shepherd, J.H.; Uray, I.P.; Mazumdar, A.; Tsimelzon, A.; Savage, M.; Hilsenbeck, S.G.; Brown, P.H. The SOX11 transcription factor is a critical regulator of basal-like breast cancer growth, invasion, and basal-like gene expression. Oncotarget 2016, 7, 13106–13121. [Google Scholar] [CrossRef]
- Oliemuller, E.; Kogata, N.; Bland, P.; Kriplani, D.; Daley, F.; Haider, S.; Shah, V.; Sawyer, E.J.; Howard, B.A. SOX11 promotes invasive growth and ductal carcinoma in situ progression. J. Pathol. 2017, 243, 193–207. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef] [Green Version]
- Malta, T.M.; Sokolov, A.; Gentles, A.J.; Burzykowski, T.; Poisson, L.; Weinstein, J.N.; Kamińska, B.; Huelsken, J.; Omberg, L.; Gevaert, O.; et al. Machine Learning Identifies Stemness Features Associated with Oncogenic Dedifferentiation. Cell 2018, 173, 338–354.e315. [Google Scholar] [CrossRef]
- Cohen, R.; Hain, E.; Buhard, O.; Guilloux, A.; Bardier, A.; Kaci, R.; Bertheau, P.; Renaud, F.; Bibeau, F.; Fléjou, J.F.; et al. Association of Primary Resistance to Immune Checkpoint Inhibitors in Metastatic Colorectal Cancer with Misdiagnosis of Microsatellite Instability or Mismatch Repair Deficiency Status. JAMA Oncol. 2019, 5, 551–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passaro, A.; Stenzinger, A.; Peters, S. Tumor Mutational Burden as a Pan-cancer Biomarker for Immunotherapy: The Limits and Potential for Convergence. Cancer Cell 2020, 38, 624–625. [Google Scholar] [CrossRef] [PubMed]
- Bonneville, R.; Krook, M.A.; Kautto, E.A.; Miya, J.; Wing, M.R.; Chen, H.Z.; Reeser, J.W.; Yu, L.; Roychowdhury, S. Landscape of Microsatellite Instability Across 39 Cancer Types. JCO Precis. Oncol. 2017, 2017, 1–15. [Google Scholar] [CrossRef]
- Davies, H.; Glodzik, D.; Morganella, S.; Yates, L.R.; Staaf, J.; Zou, X.; Ramakrishna, M.; Martin, S.; Boyault, S.; Sieuwerts, A.M.; et al. HRDetect is a predictor of BRCA1 and BRCA2 deficiency based on mutational signatures. Nat. Med. 2017, 23, 517–525. [Google Scholar] [CrossRef]
- Yoshihara, K.; Shahmoradgoli, M.; Martínez, E.; Vegesna, R.; Kim, H.; Torres-Garcia, W.; Treviño, V.; Shen, H.; Laird, P.W.; Levine, D.A.; et al. Inferring tumour purity and stromal and immune cell admixture from expression data. Nat. Commun. 2013, 4, 2612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, A.M.; Liu, C.L.; Green, M.R.; Gentles, A.J.; Feng, W.; Xu, Y.; Hoang, C.D.; Diehn, M.; Alizadeh, A.A. Robust enumeration of cell subsets from tissue expression profiles. Nat. Methods 2015, 12, 453–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mozos, A.; Royo, C.; Hartmann, E.; De Jong, D.; Baró, C.; Valera, A.; Fu, K.; Weisenburger, D.D.; Delabie, J.; Chuang, S.S.; et al. SOX11 expression is highly specific for mantle cell lymphoma and identifies the cyclin D1-negative subtype. Haematologica 2009, 94, 1555–1562. [Google Scholar] [CrossRef] [Green Version]
- Balta, E.A.; Wittmann, M.T.; Jung, M.; Sock, E.; Haeberle, B.M.; Heim, B.; von Zweydorf, F.; Heppt, J.; von Wittgenstein, J.; Gloeckner, C.J.; et al. Phosphorylation Modulates the Subcellular Localization of SOX11. Front. Mol. Neurosci. 2018, 11, 211. [Google Scholar] [CrossRef]
- Cao, X.; Fan, L.; Fang, C.; Zhu, D.X.; Dong, H.J.; Wang, D.M.; Wang, Y.H.; Xu, W.; Li, J.Y. The expression of SOX11, cyclin D1, cyclin D2, and cyclin D3 in B-cell lymphocytic proliferative diseases. Med. Oncol. 2012, 29, 1190–1196. [Google Scholar] [CrossRef]
- Oliemuller, E.; Newman, R.; Tsang, S.M.; Foo, S.; Muirhead, G.; Noor, F.; Haider, S.; Aurrekoetxea-Rodríguez, I.; Vivanco, M.D.; Howard, B.A. SOX11 promotes epithelial/mesenchymal hybrid state and alters tropism of invasive breast cancer cells. eLife 2020, 9, e58374. [Google Scholar] [CrossRef]
- Pan, J.K.; Lin, C.H.; Kuo, Y.L.; Ger, L.P.; Cheng, H.C.; Yao, Y.C.; Hsiao, M.; Lu, P.J. MiR-211 determines brain metastasis specificity through SOX11/NGN2 axis in triple-negative breast cancer. Oncogene 2021, 40, 1737–1751. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Liu, G.; Jin, H.; Chen, X.; He, J.; Xiao, J.; Qin, Y.; Mao, Y.; Zhao, L. The Dysregulation of SOX Family Correlates with DNA Methylation and Immune Microenvironment Characteristics to Predict Prognosis in Hepatocellular Carcinoma. Dis. Markers 2022, 2022, 2676114. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Zhou, C.; Zhang, J.; Cai, Q.; Li, J.; Du, T.; Zhu, Z.; Cui, X.; Liu, B. The metastasis suppressor SOX11 is an independent prognostic factor for improved survival in gastric cancer. Int. J. Oncol. 2014, 44, 1512–1520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Dai, Y.; Zhang, D.; Zhang, X.; He, Z.; Xie, X.; Cai, C. LINC00961 inhibits the migration and invasion of colon cancer cells by sponging miR-223-3p and targeting SOX11. Cancer Med. 2020, 9, 2514–2523. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Chen, G.; Chen, Y.; Guo, Z.; Chai, H.; Tang, Y.; Zheng, L.; Wei, K.; Pan, C.; Ma, Z.; et al. MiR-937-3p promotes metastasis and angiogenesis and is activated by MYC in lung adenocarcinoma. Cancer Cell Int. 2022, 22, 31. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.T.; Peng, Z.; Han, J.Y.; Lin, F.Z.; Bu, X.M.; Xu, Q.X. Clinical and prognostic significance of SOX11 in breast cancer. Asian Pac. J. Cancer Prev. 2014, 15, 5483–5486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Wang, L.; Li, J.; Saksena, A.; Wang, S.A.; Shen, J.; Hu, Z.; Lin, P.; Tang, G.; Yin, C.C.; et al. SOX11-negative Mantle Cell Lymphoma: Clinicopathologic and Prognostic Features of 75 Patients. Am. J. Surg. Pathol. 2019, 43, 710–716. [Google Scholar] [CrossRef]
- Seok, J.; Gil, M.; Dayem, A.A.; Saha, S.K.; Cho, S.G. Multi-Omics Analysis of SOX4, SOX11, and SOX12 Expression and the Associated Pathways in Human Cancers. J. Pers. Med. 2021, 11, 823. [Google Scholar] [CrossRef]
- Weinstein, J.N.; Collisson, E.A.; Mills, G.B.; Shaw, K.R.; Ozenberger, B.A.; Ellrott, K.; Shmulevich, I.; Sander, C.; Stuart, J.M. The Cancer Genome Atlas Pan-Cancer analysis project. Nat. Genet. 2013, 45, 1113–1120. [Google Scholar] [CrossRef]
- Saunus, J.M.; McCart Reed, A.E.; Lim, Z.L.; Lakhani, S.R. Breast Cancer Brain Metastases: Clonal Evolution in Clinical Context. Int. J. Mol. Sci. 2017, 18, 152. [Google Scholar] [CrossRef]
- Zawerton, A.; Yao, B.; Yeager, J.P.; Pippucci, T.; Haseeb, A.; Smith, J.D.; Wischmann, L.; Kühl, S.J.; Dean, J.C.S.; Pilz, D.T.; et al. De Novo SOX4 Variants Cause a Neurodevelopmental Disease Associated with Mild Dysmorphism. Am. J. Hum. Genet. 2019, 104, 246–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walcher, L.; Kistenmacher, A.K.; Suo, H.; Kitte, R.; Dluczek, S.; Strauß, A.; Blaudszun, A.R.; Yevsa, T.; Fricke, S.; Kossatz-Boehlert, U. Cancer Stem Cells-Origins and Biomarkers: Perspectives for Targeted Personalized Therapies. Front. Immunol. 2020, 11, 1280. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Huang, S.; Hou, Y.; Liu, Y.; Ni, M.; Meng, F.; Wang, K.; Rui, Y.; Jiang, X.; Li, G. SOX11-modified mesenchymal stem cells (MSCs) accelerate bone fracture healing: SOX11 regulates differentiation and migration of MSCs. FASEB J. 2015, 29, 1143–1152. [Google Scholar] [CrossRef] [Green Version]
- Larson, B.L.; Ylostalo, J.; Lee, R.H.; Gregory, C.; Prockop, D.J. SOX11 is expressed in early progenitor human multipotent stromal cells and decreases with extensive expansion of the cells. Tissue Eng. Part A 2010, 16, 3385–3394. [Google Scholar] [CrossRef] [Green Version]
- Bhattaram, P.; Muschler, G.; Wixler, V.; Lefebvre, V. Inflammatory Cytokines Stabilize SOXC Transcription Factors to Mediate the Transformation of Fibroblast-Like Synoviocytes in Arthritic Disease. Arthritis Rheumatol. 2018, 70, 371–382. [Google Scholar] [CrossRef] [Green Version]
- Venkov, C.; Plieth, D.; Ni, T.; Karmaker, A.; Bian, A.; George, A.L., Jr.; Neilson, E.G. Transcriptional networks in epithelial-mesenchymal transition. PLoS ONE 2011, 6, e25354. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Xie, Q.; Qin, Q.; Liang, Y.; Lin, H.; Zeng, D. Upregulation of SOX11 enhances tamoxifen resistance and promotes epithelial-to-mesenchymal transition via slug in MCF-7 breast cancer cells. J. Cell. Physiol. 2020, 235, 7295–7308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luchini, C.; Bibeau, F.; Ligtenberg, M.J.L.; Singh, N.; Nottegar, A.; Bosse, T.; Miller, R.; Riaz, N.; Douillard, J.Y.; Andre, F.; et al. ESMO recommendations on microsatellite instability testing for immunotherapy in cancer, and its relationship with PD-1/PD-L1 expression and tumour mutational burden: A systematic review-based approach. Ann. Oncol. 2019, 30, 1232–1243. [Google Scholar] [CrossRef] [Green Version]
- Ganesh, K.; Stadler, Z.K.; Cercek, A.; Mendelsohn, R.B.; Shia, J.; Segal, N.H.; Diaz, L.A., Jr. Immunotherapy in colorectal cancer: Rationale, challenges and potential. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 361–375. [Google Scholar] [CrossRef]
- Shan, T.; Uyar, D.S.; Wang, L.-S.; Mutch, D.G.; Huang, T.H.M.; Rader, J.S.; Sheng, X.; Huang, Y.-W. SOX11 hypermethylation as a tumor biomarker in endometrial cancer. Biochimie 2019, 162, 8–14. [Google Scholar] [CrossRef]
- Korneev, K.V.; Atretkhany, K.N.; Drutskaya, M.S.; Grivennikov, S.I.; Kuprash, D.V.; Nedospasov, S.A. TLR-signaling and proinflammatory cytokines as drivers of tumorigenesis. Cytokine 2017, 89, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Balsas, P.; Palomero, J.; Eguileor, Á.; Rodríguez, M.L.; Vegliante, M.C.; Planas-Rigol, E.; Sureda-Gómez, M.; Cid, M.C.; Campo, E.; Amador, V. SOX11 promotes tumor protective microenvironment interactions through CXCR4 and FAK regulation in mantle cell lymphoma. Blood 2017, 130, 501–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balsas, P.; Veloza, L.; Clot, G.; Sureda-Gómez, M.; Rodríguez, M.L.; Masaoutis, C.; Frigola, G.; Navarro, A.; Beà, S.; Nadeu, F.; et al. SOX11, CD70, and Treg cells configure the tumor-immune microenvironment of aggressive mantle cell lymphoma. Blood 2021, 138, 2202–2215. [Google Scholar] [CrossRef] [PubMed]
- Su, D.; Gao, Q.; Guan, L.; Sun, P.; Li, Q.; Shi, C.; Ma, X. Downregulation of SOX11 in fetal heart tissue, under hyperglycemic environment, mediates cardiomyocytes apoptosis. J. Biochem. Mol. Toxicol. 2021, 35, e22629. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhong, Y.; Chen, Y.J.; Chen, H. SOX11 regulates apoptosis and cell cycle in hepatocellular carcinoma via Wnt/β-catenin signaling pathway. Biotechnol. Appl. Biochem. 2019, 66, 240–246. [Google Scholar] [CrossRef] [PubMed]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Q.; Du, J.; Dong, J.; Pan, S.; Jin, H.; Han, X.; Zhang, J. Systematic Investigation of the Multifaceted Role of SOX11 in Cancer. Cancers 2022, 14, 6103. https://doi.org/10.3390/cancers14246103
Sun Q, Du J, Dong J, Pan S, Jin H, Han X, Zhang J. Systematic Investigation of the Multifaceted Role of SOX11 in Cancer. Cancers. 2022; 14(24):6103. https://doi.org/10.3390/cancers14246103
Chicago/Turabian StyleSun, Qingqing, Jun Du, Jie Dong, Shuaikang Pan, Hongwei Jin, Xinghua Han, and Jinguo Zhang. 2022. "Systematic Investigation of the Multifaceted Role of SOX11 in Cancer" Cancers 14, no. 24: 6103. https://doi.org/10.3390/cancers14246103