

miR-214-Enriched Extracellular Vesicles Released by Acid-Adapted Melanoma Cells Promote Inflammatory Macrophage-Dependent Tumor Trans-Endothelial Migration
 ,
,  ,
,  , , ,
, , ,  , , , and
, , , and
Abstract
:Simple Summary
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Treatments
2.2. Extracellular Vesicles (EV)
2.2.1. Isolation of EV through the Ultracentrifugation Method
2.2.2. Transmission Electron Microscopy
2.2.3. Nanoparticle Tracking Analysis
2.3. RNA Isolation and Quantitative Real-Time Polymerase Chain Reaction (qPCR)
2.3.1. miRNA
2.3.2. mRNA
2.4. Immunofluorescence
2.5. Western Blot
2.6. Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. Nitric Oxide (NO) Assay
2.8. Lactate Measure
2.9. Seahorse Analysis
2.10. Flow Cytometry
2.11. In Vitro Permeability Assay
2.12. In Vitro Trans-Endothelial Migration
2.13. Statistical Analysis
3. Results
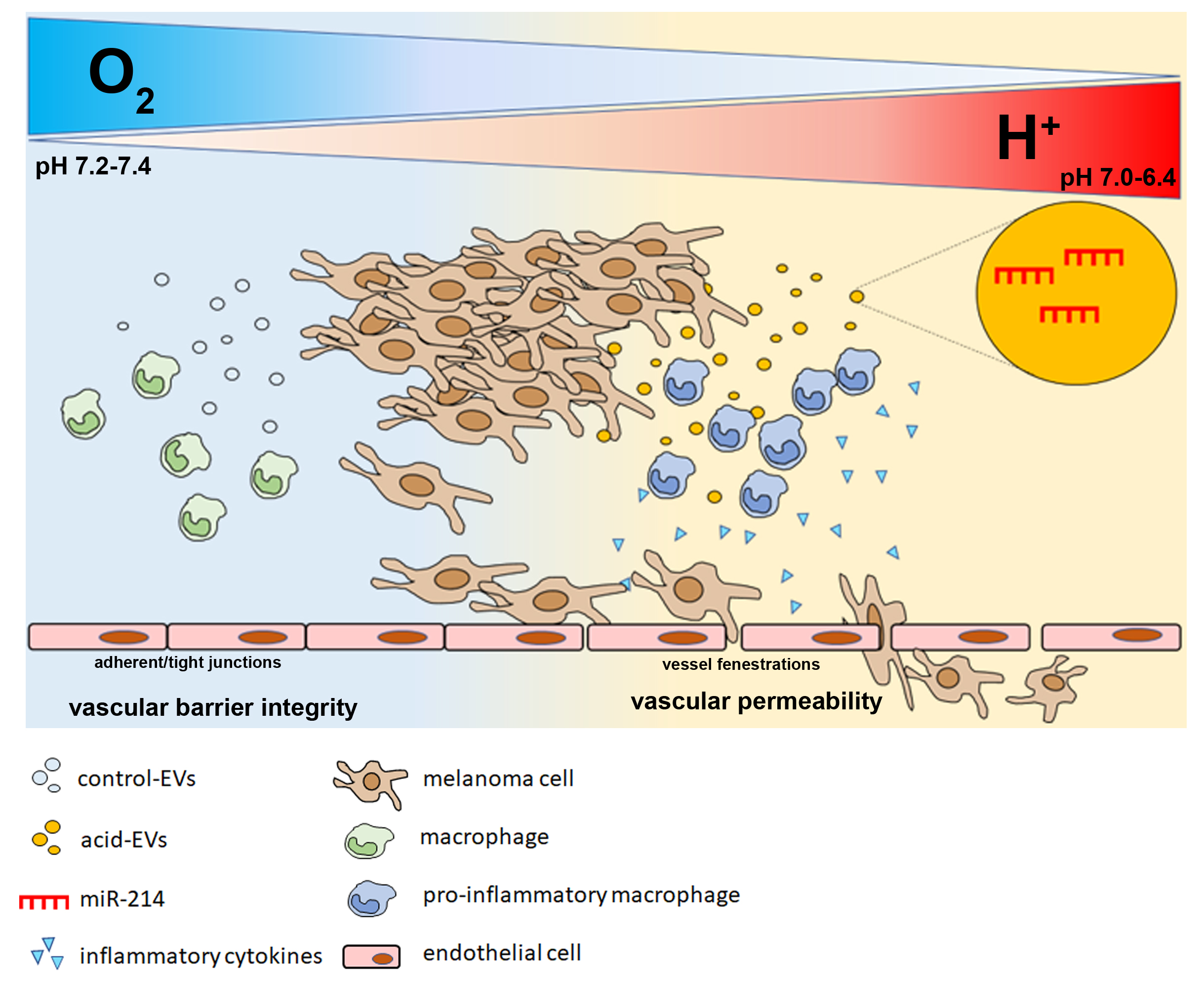
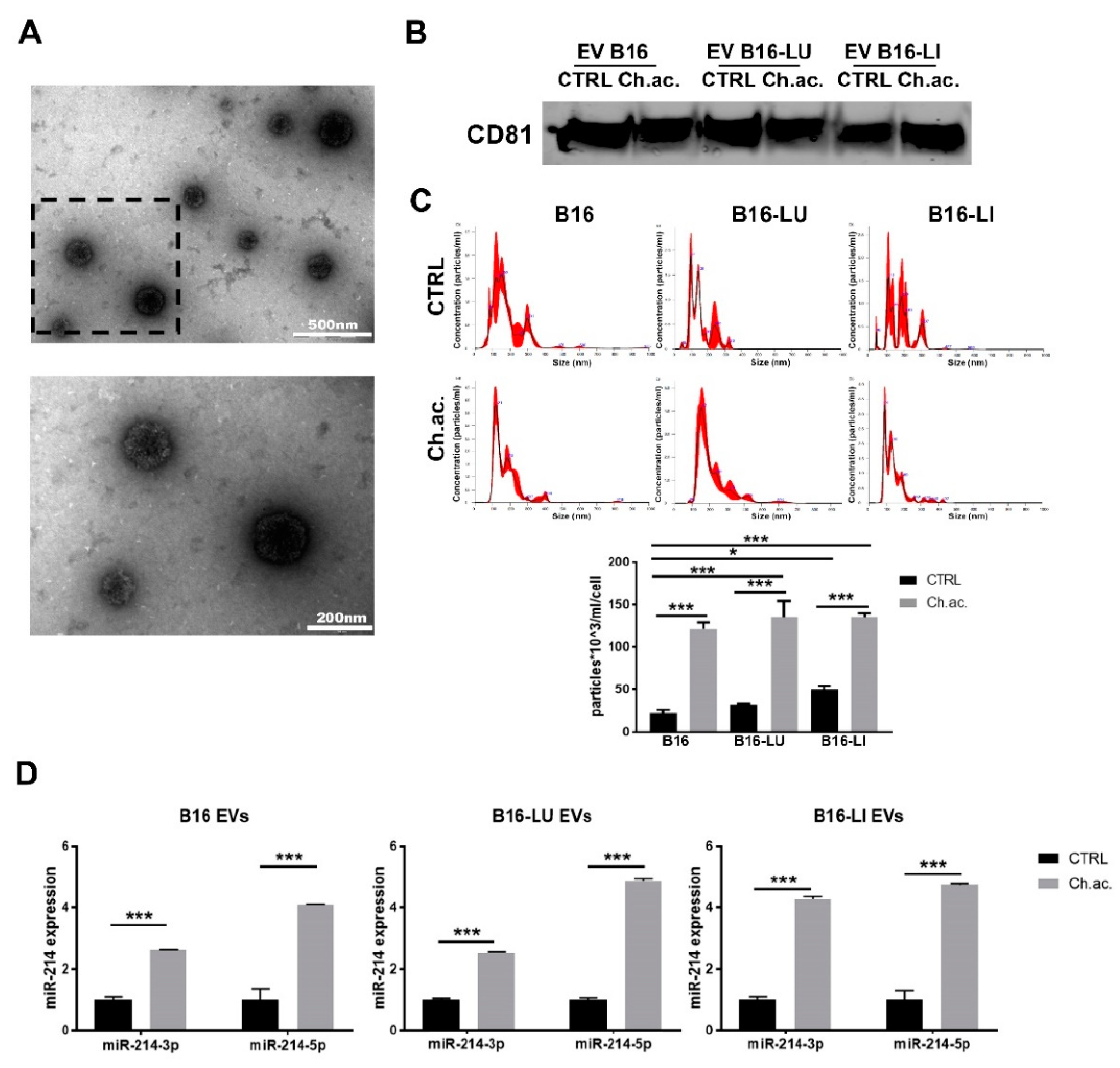
3.1. Acid-Adapted Melanoma Cells Released a Higher Amount of miR-214-Carrying EV Compared to Controls
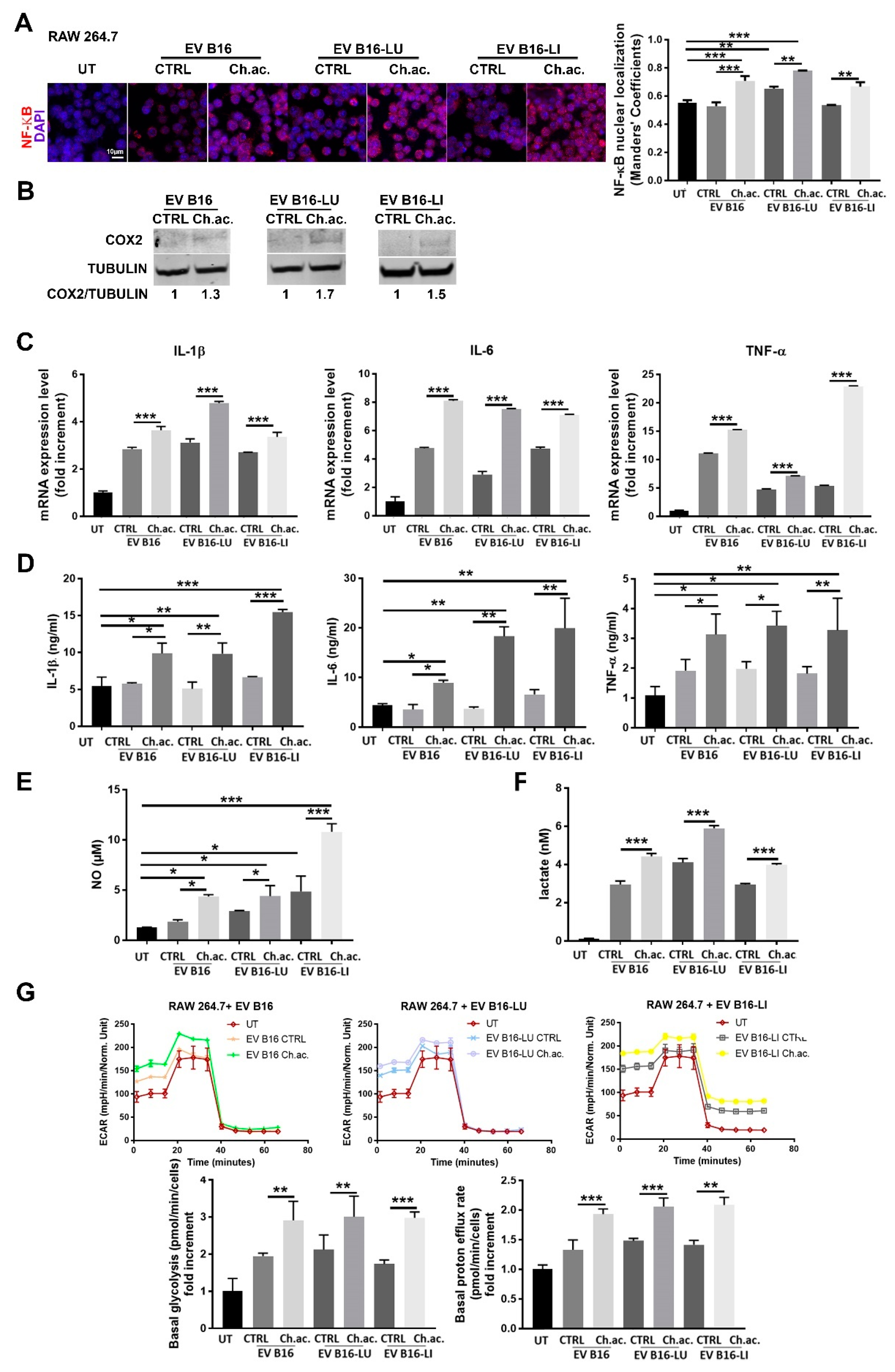
3.2. miR-214-Enriched EV Released by Acid-Adapted Melanoma Cells Induce a Pro-Inflammatory Response in RAW 264.7 Macrophages
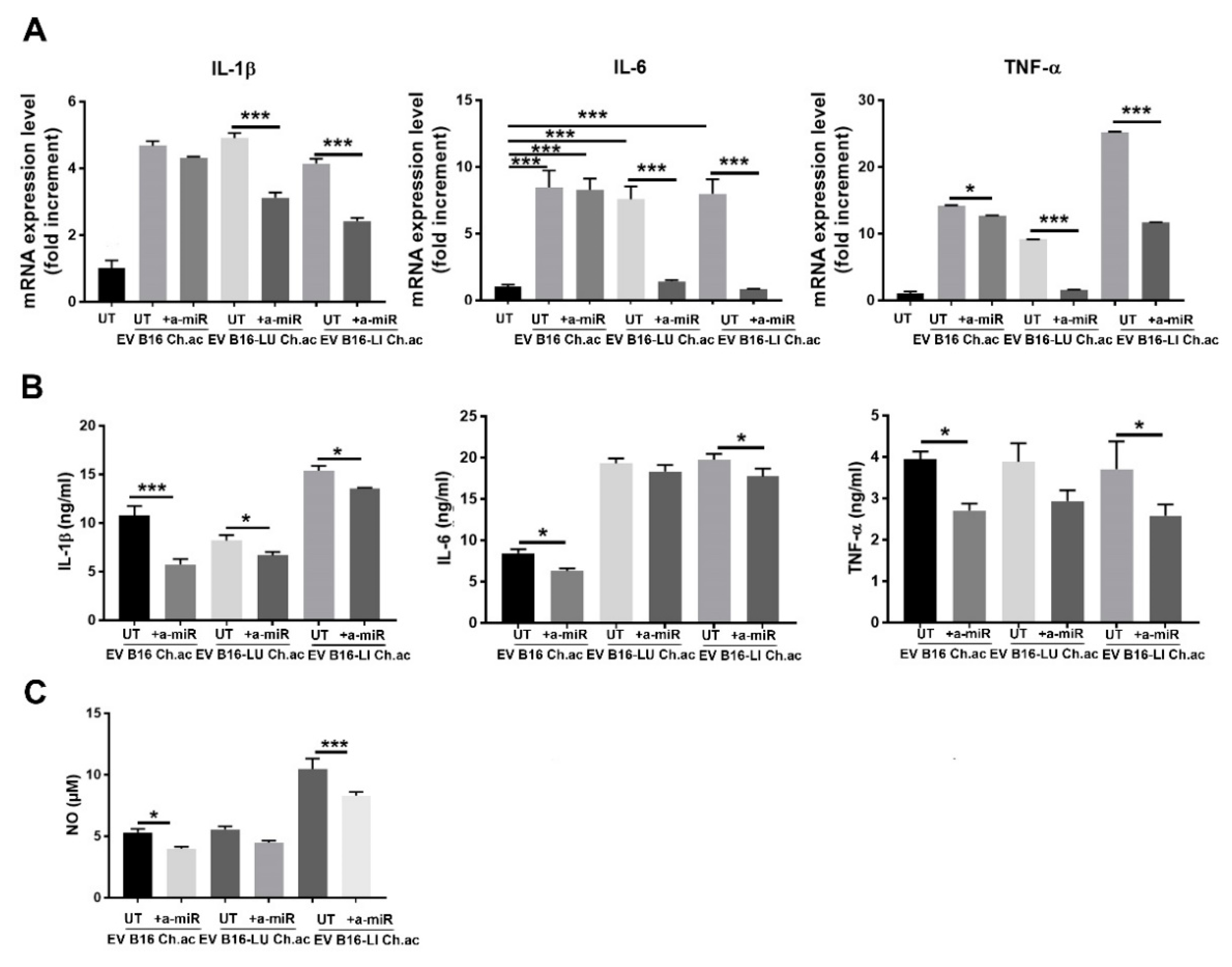
3.3. miR-214 Interfering Counteracts the Pro-Inflammatory Response of Macrophages upon Acid-EV Uptake
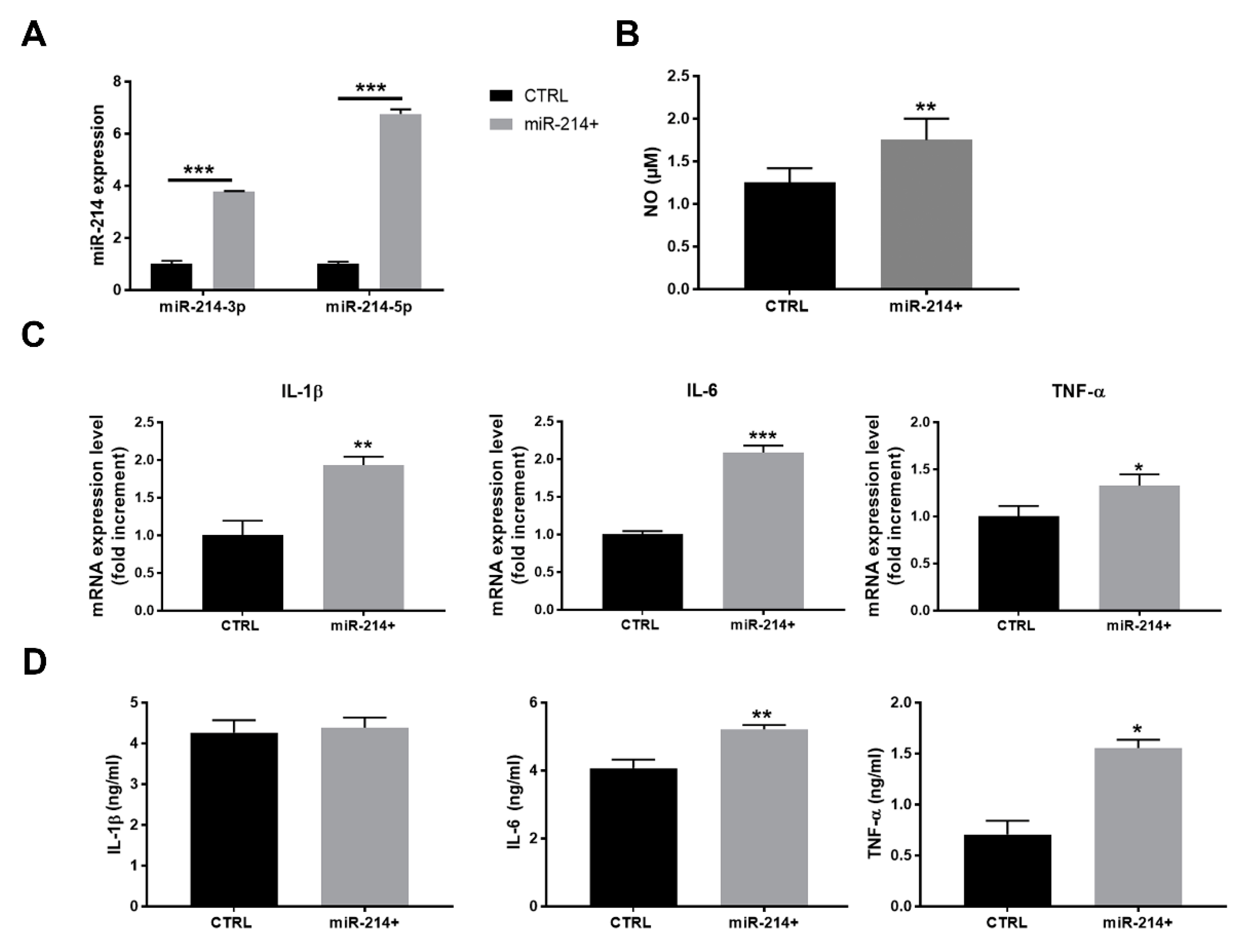
3.4. miR-214 Forced Expression in Macrophages Promotes the Pro-Inflammatory Phenotype
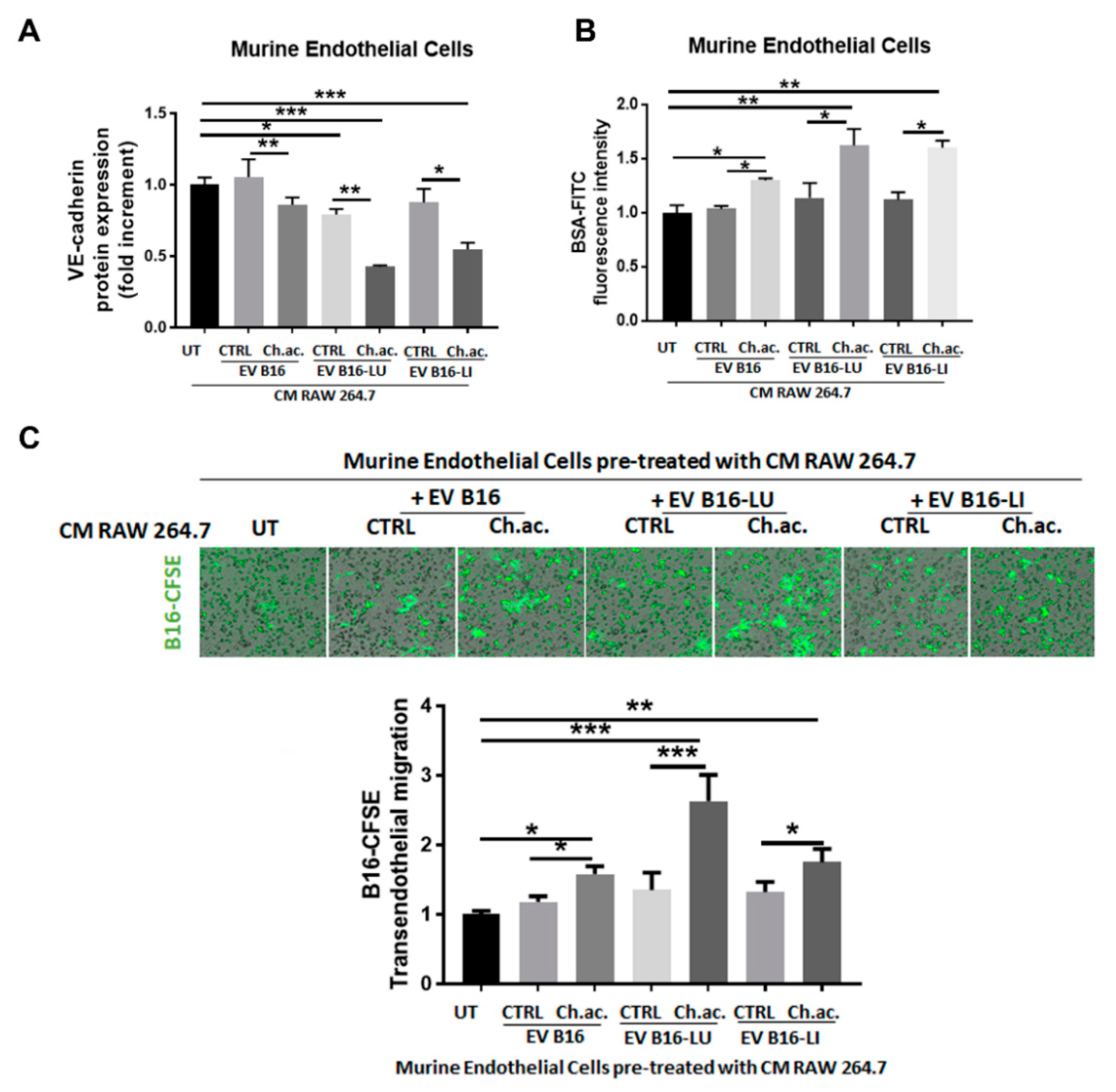
3.5. Conditioned Media of Acid-EV-Treated Macrophages Increase Vascular Permeability and Facilitate the Transendothelial Migration of Melanoma Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Ali, Z.; Yousaf, N.; Larkin, J. Melanoma epidemiology, biology and prognosis. EJC Suppl. 2013, 11, 81–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davey, M.G.; Miller, N.; McInerney, N.M. A review of epidemiology and cancer biology of malignant melanoma. Cureus 2021, 13, e15087. [Google Scholar] [CrossRef]
- Minciacchi, V.R.; Freeman, M.R.; Di Vizio, D. Extracellular vesicles in cancer: Exosomes, microvesicles and the emerging role of large oncosomes. Semin. Cell Dev. Biol. 2015, 40, 41–51. [Google Scholar] [CrossRef] [Green Version]
- van der Pol, E.; Böing, A.N.; Harrison, P.; Sturk, A.; Nieuwland, R. Classification, functions, and clinical relevance of extracellular vesicles. Pharmacol. Rev. 2012, 64, 676–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noren Hooten, N.; Yáñez-Mó, M.; DeRita, R.; Russell, A.; Quesenberry, P.; Ramratnam, B.; Robbins, P.D.; Di Vizio, D.; Wen, S.; Witwer, K.W.; et al. Hitting the bullseye: Are extracellular vesicles on target? J. Extracell. Vesicles 2020, 10, e12032. [Google Scholar] [CrossRef] [PubMed]
- Becker, A.; Thakur, B.K.; Weiss, J.M.; Kim, H.S.; Peinado, H.; Lyden, D. Extracellular vesicles in cancer: Cell-to-cell mediators of metastasis. Cancer Cell 2016, 30, 836–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA biogenesis, mechanisms of actions, and circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biagioni, A.; Tavakol, S.; Ahmadirad, N.; Zahmatkeshan, M.; Magnelli, L.; Mandegary, A.; Samareh Fekri, H.; Asadi, M.H.; Mohammadinejad, R.; Ahn, K.S. Small nucleolar RNA host genes promoting epithelial-mesenchymal transition lead cancer progression and metastasis. IUBMB Life 2021, 73, 825–842. [Google Scholar] [CrossRef]
- Penna, E.; Orso, F.; Cimino, D.; Tenaglia, E.; Lembo, A.; Quaglino, E.; Poliseno, L.; Haimovic, A.; Osella-Abate, S.; De Pittà, C.; et al. MicroRNA-214 contributes to melanoma tumor progression through suppression of TFAP2C: MiR-214 and melanoma progression. EMBO J. 2011, 30, 1990–2007. [Google Scholar] [CrossRef]
- Orso, F.; Quirico, L.; Virga, F.; Penna, E.; Dettori, D.; Cimino, D.; Coppo, R.; Grassi, E.; Elia, A.R.; Brusa, D.; et al. MiR-214 and MiR-148b targeting inhibits dissemination of melanoma and breast cancer. Cancer Res. 2016, 76, 5151–5162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penna, E.; Orso, F.; Cimino, D.; Vercellino, I.; Grassi, E.; Quaglino, E.; Turco, E.; Taverna, D. MiR-214 Coordinates Melanoma Progression by Upregulating ALCAM through TFAP2 and MiR-148b Downmodulation. Cancer Res. 2013, 73, 4098–4111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dettori, D.; Orso, F.; Penna, E.; Baruffaldi, D.; Brundu, S.; Maione, F.; Turco, E.; Giraudo, E.; Taverna, D. Therapeutic silencing of MiR-214 inhibits tumor progression in multiple mouse models. Mol. Ther. 2018, 26, 2008–2018. [Google Scholar] [CrossRef] [Green Version]
- Peppicelli, S.; Bianchini, F.; Calorini, L. Inflammatory cytokines induce vascular endothelial growth factor-C expression in melanoma-associated macrophages and stimulate melanoma lymph node metastasis. Oncol. Lett. 2014, 8, 1133–1138. [Google Scholar] [CrossRef] [Green Version]
- Andreucci, E.; Laurenzana, A.; Peppicelli, S.; Biagioni, A.; Margheri, F.; Ruzzolini, J.; Bianchini, F.; Fibbi, G.; Del Rosso, M.; Nediani, C.; et al. UPAR controls vasculogenic mimicry ability expressed by drug-resistant melanoma cells. Oncol. Res. 2022, 28, 873–884. [Google Scholar] [CrossRef]
- Biagioni, A.; Laurenzana, A.; Menicacci, B.; Peppicelli, S.; Andreucci, E.; Bianchini, F.; Guasti, D.; Paoli, P.; Serratì, S.; Mocali, A.; et al. UPAR-expressing melanoma exosomes promote angiogenesis by VE-cadherin, EGFR and UPAR overexpression and rise of ERK1,2 signaling in endothelial cells. Cell Mol. Life Sci. 2021, 78, 3057–3072. [Google Scholar] [CrossRef]
- Onogi, Y.; Wada, T.; Okekawa, A.; Matsuzawa, T.; Watanabe, E.; Ikeda, K.; Nakano, M.; Kitada, M.; Koya, D.; Tsuneki, H.; et al. Pro-inflammatory macrophages coupled with glycolysis remodel adipose vasculature by producing platelet-derived growth factor-B in obesity. Sci. Rep. 2020, 10, 670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duong, C.N.; Vestweber, D. Mechanisms ensuring endothelial junction integrity beyond VE-cadherin. Front. Physiol. 2020, 11, 519. [Google Scholar] [CrossRef]
- Vander Heiden, M.G.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg effect: The metabolic requirements of cell proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [Green Version]
- Vaupel, P. Tumor microenvironmental physiology and its implications for radiation oncology. Semin. Radiat. Oncol. 2004, 14, 198–206. [Google Scholar] [CrossRef]
- Reshetnyak, Y.K.; Yao, L.; Zheng, S.; Kuznetsov, S.; Engelman, D.M.; Andreev, O.A. Measuring tumor aggressiveness and targeting metastatic lesions with fluorescent PHLIP. Mol. Imaging Biol. 2011, 13, 1146–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pillai, S.R.; Damaghi, M.; Marunaka, Y.; Spugnini, E.P.; Fais, S.; Gillies, R.J. Causes, consequences, and therapy of tumors acidosis. Cancer Metastasis Rev. 2019, 38, 205–222. [Google Scholar] [CrossRef] [PubMed]
- Peppicelli, S.; Andreucci, E.; Ruzzolini, J.; Laurenzana, A.; Margheri, F.; Fibbi, G.; Del Rosso, M.; Bianchini, F.; Calorini, L. The acidic microenvironment as a possible niche of dormant tumor cells. Cell. Mol. Life Sci. 2017, 74, 2761–2771. [Google Scholar] [CrossRef]
- Andreucci, E.; Peppicelli, S.; Ruzzolini, J.; Bianchini, F.; Biagioni, A.; Papucci, L.; Magnelli, L.; Mazzanti, B.; Stecca, B.; Calorini, L. The acidic tumor microenvironment drives a stem-like phenotype in melanoma cells. J. Mol. Med. 2020, 98, 1431–1446. [Google Scholar] [CrossRef] [PubMed]
- Webb, B.A.; Chimenti, M.; Jacobson, M.P.; Barber, D.L. Dysregulated PH: A perfect storm for cancer progression. Nat. Rev. Cancer 2011, 11, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Ruzzolini, J.; Peppicelli, S.; Andreucci, E.; Bianchini, F.; Margheri, F.; Laurenzana, A.; Fibbi, G.; Pimpinelli, N.; Calorini, L. Everolimus selectively targets vemurafenib resistant BRAFV600E melanoma cells adapted to low pH. Cancer Lett. 2017, 408, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Peppicelli, S.; Ruzzolini, J.; Bianchini, F.; Andreucci, E.; Nediani, C.; Laurenzana, A.; Margheri, F.; Fibbi, G.; Calorini, L. Anoikis resistance as a further trait of acidic-adapted melanoma cells. J. Oncol. 2019, 2019, 8340926. [Google Scholar] [CrossRef]
- Peppicelli, S.; Bianchini, F.; Torre, E.; Calorini, L. Contribution of acidic melanoma cells undergoing epithelial-to-mesenchymal transition to aggressiveness of non-acidic melanoma cells. Clin. Exp. Metastasis 2014, 31, 423–433. [Google Scholar] [CrossRef]
- Han, L.; Lam, E.W.-F.; Sun, Y. Extracellular vesicles in the tumor microenvironment: Old stories, but new tales. Mol. Cancer 2019, 18, 59. [Google Scholar] [CrossRef] [Green Version]
- Logozzi, M.; Spugnini, E.; Mizzoni, D.; Di Raimo, R.; Fais, S. Extracellular acidity and increased exosome release as key phenotypes of malignant tumors. Cancer Metastasis Rev. 2019, 38, 93–101. [Google Scholar] [CrossRef]
- Logozzi, M.; Mizzoni, D.; Angelini, D.; Di Raimo, R.; Falchi, M.; Battistini, L.; Fais, S. Microenvironmental pH and exosome levels interplay in human cancer cell lines of different histotypes. Cancers 2018, 10, 370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parolini, I.; Federici, C.; Raggi, C.; Lugini, L.; Palleschi, S.; De Milito, A.; Coscia, C.; Iessi, E.; Logozzi, M.; Molinari, A.; et al. Microenvironmental pH is a key factor for exosome traffic in tumor cells. J. Biol. Chem. 2009, 284, 34211–34222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.-J.; Li, W.-W.; Wen, C.-J.; Diao, Y.-L.; Zhao, T.-L. MicroRNA-214 Promotes the EMT process in melanoma by downregulating CADM1 expression. Mol. Med. Rep. 2020, 22, 3795–3803. [Google Scholar] [CrossRef] [PubMed]
- Guanen, Q.; Junjie, S.; Baolin, W.; Chaoyang, W.; Yajuan, Y.; Jing, L.; Junpeng, L.; Gaili, N.; Zhongping, W.; Jun, W. MiR-214 promotes cell meastasis and inhibites apoptosis of esophageal squamous cell carcinoma via PI3K/AKT/MTOR signaling pathway. Biomed. Pharmacother. 2018, 105, 350–361. [Google Scholar] [CrossRef] [PubMed]
- Yahya, S.M.M.; Yahya, S.M.M. The effect of MiR-98 and MiR-214 on apoptotic and angiogenic pathways in hepatocellular carcinoma HepG2 cells. Indian J. Clin. Biochem. 2020, 35, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xu, R.; Gu, H.; Zhang, E.; Qu, J.; Cao, W.; Huang, X.; Yan, H.; He, J.; Cai, Z. Metabolic reprogramming in macrophage responses. Biomark. Res. 2021, 9, 1. [Google Scholar] [CrossRef]
- Li, L.-M.; Hou, D.-X.; Guo, Y.-L.; Yang, J.-W.; Liu, Y.; Zhang, C.-Y.; Zen, K. Role of microRNA-214–targeting phosphatase and tensin homolog in advanced glycation end product-induced apoptosis delay in monocytes. J. Immunol. 2011, 186, 2552–2560. [Google Scholar] [CrossRef] [Green Version]
- Jindra, P.T.; Bagley, J.; Godwin, J.G.; Iacomini, J. Costimulation-dependent expression of microRNA-214 increases the ability of T cells to proliferate by targeting. Pten. J. Immunol. 2010, 185, 990–997. [Google Scholar] [CrossRef] [Green Version]
- Li, X.-F.; Chen, X.; Bao, J.; Xu, L.; Zhang, L.; Huang, C.; Meng, X.-M.; Li, J. PTEN negatively regulates the expression of pro-inflammatory cytokines and chemokines of fibroblast-like synoviocytes in adjuvant-induced arthritis. Artif. Cells Nanomed. Biotechnol. 2019, 47, 3687–3696. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.; Jiang, L.; Zou, H.; Chen, T.; Liang, H.; Tang, L. PTEN suppresses the inflammation, viability, and motility of AP-AR42J cells by activating the Wnt/β-catenin pathway. RSC Adv. 2019, 9, 5460–5469. [Google Scholar] [CrossRef]
- Polytarchou, C.; Hommes, D.W.; Palumbo, T.; Hatziapostolou, M.; Koutsioumpa, M.; Koukos, G.; van der Meulen-de Jong, A.E.; Oikonomopoulos, A.; van Deen, W.K.; Vorvis, C.; et al. MicroRNA214 is associated with progression of ulcerative colitis, and inhibition reduces development of colitis and colitis-associated cancer in mice. Gastroenterology 2015, 149, 981–992.e11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Liu, Y.-W.; Yang, T.; Gan, L.; Yang, N.; Dai, S.-S.; He, F. The mutual regulation between MiR-214 and A2AR signaling plays an important role in inflammatory response. Cell. Signal. 2015, 27, 2026–2034. [Google Scholar] [CrossRef] [PubMed]
- Xue, N.; Qi, L.; Zhang, G.; Zhang, Y. MiRNA-125b regulates osteogenic differentiation of periodontal ligament cells through NKIRAS2/NF-ΚB pathway. Cell Physiol. Biochem. 2018, 48, 1771–1781. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-Y.; Li, L.-F.; Yang, C.-T.; Lu, K.-H.; Huang, C.-C.; Kao, K.-C.; Chiou, S.-H. Suppressing NF-ΚB and NKRF pathways by induced pluripotent stem cell therapy in mice with ventilator-induced lung injury. PLoS ONE 2013, 8, e66760. [Google Scholar] [CrossRef]
- Bychkov, M.L.; Kirichenko, A.V.; Mikhaylova, I.N.; Paramonov, A.S.; Yastremsky, E.V.; Kirpichnikov, M.P.; Shulepko, M.A.; Lyukmanova, E.N. Extracellular vesicles derived from acidified metastatic melanoma cells stimulate growth, migration, and stemness of normal keratinocytes. Biomedicines 2022, 10, 660. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andreucci, E.; Ruzzolini, J.; Bianchini, F.; Versienti, G.; Biagioni, A.; Lulli, M.; Guasti, D.; Nardini, P.; Serratì, S.; Margheri, F.; et al. miR-214-Enriched Extracellular Vesicles Released by Acid-Adapted Melanoma Cells Promote Inflammatory Macrophage-Dependent Tumor Trans-Endothelial Migration. Cancers 2022, 14, 5090. https://doi.org/10.3390/cancers14205090
Andreucci E, Ruzzolini J, Bianchini F, Versienti G, Biagioni A, Lulli M, Guasti D, Nardini P, Serratì S, Margheri F, et al. miR-214-Enriched Extracellular Vesicles Released by Acid-Adapted Melanoma Cells Promote Inflammatory Macrophage-Dependent Tumor Trans-Endothelial Migration. Cancers. 2022; 14(20):5090. https://doi.org/10.3390/cancers14205090
Chicago/Turabian StyleAndreucci, Elena, Jessica Ruzzolini, Francesca Bianchini, Giampaolo Versienti, Alessio Biagioni, Matteo Lulli, Daniele Guasti, Patrizia Nardini, Simona Serratì, Francesca Margheri, and et al. 2022. "miR-214-Enriched Extracellular Vesicles Released by Acid-Adapted Melanoma Cells Promote Inflammatory Macrophage-Dependent Tumor Trans-Endothelial Migration" Cancers 14, no. 20: 5090. https://doi.org/10.3390/cancers14205090