
The Significance of MicroRNAs in the Molecular Pathology of Brain Metastases

 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
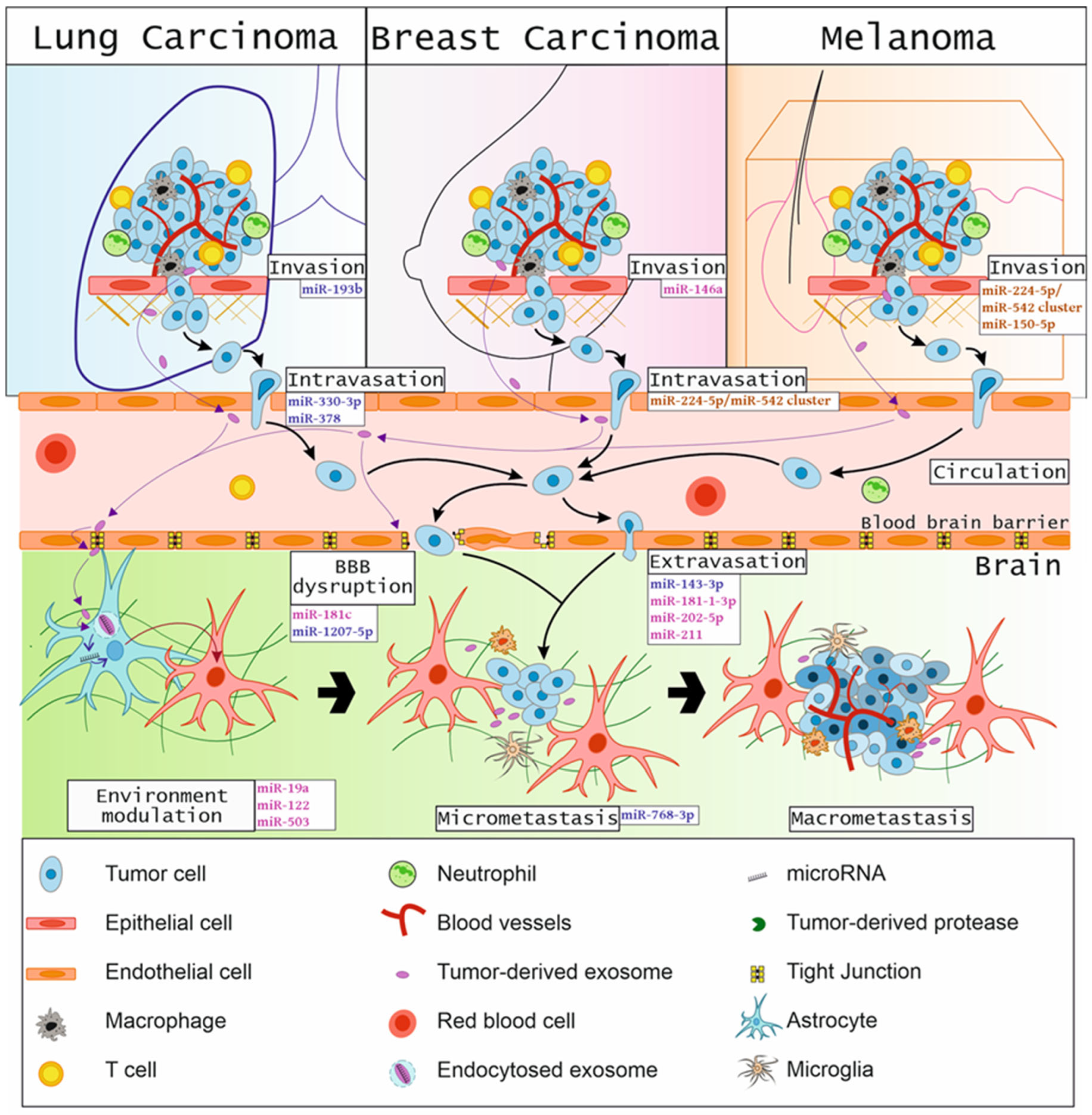
2. The Metastatic Cascade and the Development of Metastases
Specifics of the Development of Brain Metastases
3. MicroRNAs
4. MicroRNAs Involved in the Metastatic Cascade
4.1. MicroRNAs with a Suppresive Function for Metastases Development
4.2. Metastases Promoting microRNAs
4.3. MicroRNAs Involved in the Premetastaci Niche Formation and Microenvironment Modulation
5. MicroRNAs Involved in Brain Cancer Metastases
5.1. MicroRNAs in Lung Cancer Brain Metastases
5.1.1. MicroRNAs with a Suppresive Function for Metastases Development in Lung Cancer
5.1.2. Metastases Promoting MicroRNAs in Lung Cancer
5.1.3. MicroRNAs with Diagnostic and Prognostic Potential in Lung Cancer Metastases
5.2. MicroRNAs in Breast Cancer Brain Metastases
5.2.1. MicroRNAs with a Suppresive Function for Metastases Development in Breast Cancer
5.2.2. MicroRNAs Involved in the Extravasation and Colonization of the Brain in Breast Cancer Metastases
5.3. MicroRNAs in Melanoma Brain Metastases
5.4. MicroRNAs in Colorectal Carcinoma Brain Metastases
5.5. MicroRNAs in Renal Cell Carcinoma Brain Metastases
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Nayak, L.; Lee, E.Q.; Wen, P.Y. Epidemiology of Brain Metastases. Curr. Oncol. Rep. 2012, 14, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Smedby, K.E.; Brandt, L.; Bäcklund, M.L.; Blomqvist, P. Brain Metastases Admissions in Sweden between 1987 and 2006. Br. J. Cancer 2009, 101, 1919–1924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niemiec, M.; Głogowski, M.; Tyc-Szczepaniak, D.; Wierzchowski, M.; Kępka, L. Characteristics of Long-Term Survivors of Brain Metastases from Lung Cancer. Rep. Pract. Oncol. Radiother. 2011, 16, 49–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watabe, K. Non-Coding RNAs in Cancer Brain Metastasis. Front. Biosci. 2016, 8, 187–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sperduto, P.W.; Kased, N.; Roberge, D.; Xu, Z.; Shanley, R.; Luo, X.; Sneed, P.K.; Chao, S.T.; Weil, R.J.; Suh, J.; et al. Summary Report on the Graded Prognostic Assessment: An Accurate and Facile Diagnosis-Specific Tool to Estimate Survival for Patients with Brain Metastases. J. Clin. Oncol. 2012, 30, 419–425. [Google Scholar] [CrossRef] [Green Version]
- Barnholtz-Sloan, J.S.; Sloan, A.E.; Davis, F.G.; Vigneau, F.D.; Lai, P.; Sawaya, R.E. Incidence Proportions of Brain Metastases in Patients Diagnosed (1973 to 2001) in the Metropolitan Detroit Cancer Surveillance System. J. Clin. Oncol. 2004, 22, 2865–2872. [Google Scholar] [CrossRef]
- Schouten, L.J.; Rutten, J.; Huveneers, H.A.M.; Twijnstra, A. Incidence of Brain Metastases in a Cohort of Patients with Carcinoma of the Breast, Colon, Kidney, and Lung and Melanoma. Cancer 2002, 94, 2698–2705. [Google Scholar] [CrossRef]
- Ostrom, Q.T.; Wright, C.H.; Barnholtz-Sloan, J.S. Brain Metastases: Epidemiology. In Handbook of Clinical Neurology; Elsevier: Amsterdam, The Netherlands, 2018; Volume 149, pp. 27–42. ISBN 978-0-12-811161-1. [Google Scholar]
- Cagney, D.N.; Martin, A.M.; Catalano, P.J.; Redig, A.J.; Lin, N.U.; Lee, E.Q.; Wen, P.Y.; Dunn, I.F.; Bi, W.L.; Weiss, S.E.; et al. Incidence and Prognosis of Patients with Brain Metastases at Diagnosis of Systemic Malignancy: A Population-Based Study. Neuro-Oncol. 2017, 19, 1511–1521. [Google Scholar] [CrossRef] [Green Version]
- Martin, A.M.; Cagney, D.N.; Catalano, P.J.; Warren, L.E.; Bellon, J.R.; Punglia, R.S.; Claus, E.B.; Lee, E.Q.; Wen, P.Y.; Haas-Kogan, D.A.; et al. Brain Metastases in Newly Diagnosed Breast Cancer: A Population-Based Study. JAMA Oncol. 2017, 3, 1069. [Google Scholar] [CrossRef]
- Ramakrishna, N.; Temin, S.; Chandarlapaty, S.; Crews, J.R.; Davidson, N.E.; Esteva, F.J.; Giordano, S.H.; Gonzalez-Angulo, A.M.; Kirshner, J.J.; Krop, I.; et al. Recommendations on Disease Management for Patients with Advanced Human Epidermal Growth Factor Receptor 2–Positive Breast Cancer and Brain Metastases: American Society of Clinical Oncology Clinical Practice Guideline. J. Clin. Oncol. 2014, 32, 2100–2108. [Google Scholar] [CrossRef]
- Langley, R.R.; Fidler, I.J. The Seed and Soil Hypothesis Revisited-The Role of Tumor-Stroma Interactions in Metastasis to Different Organs. Int. J. Cancer 2011, 128, 2527–2535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuzet, S.-E.; Gaggioli, C. Fibroblast Activation in Cancer: When Seed Fertilizes Soil. Cell Tissue Res. 2016, 365, 607–619. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, R.N.; Riba, R.D.; Zacharoulis, S.; Bramley, A.H.; Vincent, L.; Costa, C.; MacDonald, D.D.; Jin, D.K.; Shido, K.; Kerns, S.A.; et al. VEGFR1-Positive Haematopoietic Bone Marrow Progenitors Initiate the Pre-Metastatic Niche. Nature 2005, 438, 820–827. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, S.; Yao, J.; Lowery, F.J.; Zhang, Q.; Huang, W.-C.; Li, P.; Li, M.; Wang, X.; Zhang, C.; et al. Microenvironment-Induced PTEN Loss by Exosomal MicroRNA Primes Brain Metastasis Outgrowth. Nature 2015, 527, 100–104. [Google Scholar] [CrossRef]
- Draffin, J.E.; McFarlane, S.; Hill, A.; Johnston, P.G.; Waugh, D.J.J. CD44 Potentiates the Adherence of Metastatic Prostate and Breast Cancer Cells to Bone Marrow Endothelial Cells. Cancer Res. 2004, 64, 5702–5711. [Google Scholar] [CrossRef] [Green Version]
- Brabletz, T. To Differentiate or Not—Routes towards Metastasis. Nat. Rev. Cancer 2012, 12, 425–436. [Google Scholar] [CrossRef]
- Rettig, M.; Trinidad, K.; Pezeshkpour, G.; Frost, P.; Sharma, S.; Moatamed, F.; Tamanoi, F.; Mortazavi, F. PAK1 Kinase Promotes Cell Motility and Invasiveness through CRK-II Serine Phosphorylation in Non-Small Cell Lung Cancer Cells. PLoS ONE 2012, 7, e42012. [Google Scholar] [CrossRef]
- Craene, B.D.; Berx, G. Regulatory Networks Defining EMT during Cancer Initiation and Progression. Nat. Rev. Cancer 2013, 13, 97–110. [Google Scholar] [CrossRef]
- Mani, S.A.; Guo, W.; Liao, M.-J.; Eaton, E.N.; Ayyanan, A.; Zhou, A.Y.; Brooks, M.; Reinhard, F.; Zhang, C.C.; Shipitsin, M.; et al. The Epithelial-Mesenchymal Transition Generates Cells with Properties of Stem Cells. Cell 2008, 133, 704–715. [Google Scholar] [CrossRef] [Green Version]
- Simpson, C.D.; Anyiwe, K.; Schimmer, A.D. Anoikis Resistance and Tumor Metastasis. Cancer Lett. 2008, 272, 177–185. [Google Scholar] [CrossRef]
- Luzzi, K.J.; MacDonald, I.C.; Schmidt, E.E.; Kerkvliet, N.; Morris, V.L.; Chambers, A.F.; Groom, A.C. Multistep Nature of Metastatic Inefficiency. Am. J. Pathol. 1998, 153, 865–873. [Google Scholar] [CrossRef]
- Thiery, J.P.; Acloque, H.; Huang, R.Y.J.; Nieto, M.A. Epithelial-Mesenchymal Transitions in Development and Disease. Cell 2009, 139, 871–890. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Bu, F.; Royer, C.; Serres, S.; Larkin, J.R.; Soto, M.S.; Sibson, N.R.; Salter, V.; Fritzsche, F.; Turnquist, C.; et al. ASPP2 Controls Epithelial Plasticity and Inhibits Metastasis through β-Catenin-Dependent Regulation of ZEB1. Nat. Cell Biol. 2014, 16, 1092–1104. [Google Scholar] [CrossRef]
- Díaz-López, A.; Díaz-Martín, J.; Moreno-Bueno, G.; Cuevas, E.P.; Santos, V.; Olmeda, D.; Portillo, F.; Palacios, J.; Cano, A. Zeb1 and Snail1 Engage MiR-200f Transcriptional and Epigenetic Regulation during EMT: EMT Players Controlling Epithelial Plasticity. Int. J. Cancer 2015, 136, E62–E73. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Zhao, N.; Zhao, X.; Gu, Q.; Zhang, S.; Che, N.; Wang, X.; Du, J.; Liu, Y.; Sun, B. Expression and Functional Significance of Twist1 in Hepatocellular Carcinoma: Its Role in Vasculogenic Mimicry. Hepatology 2010, 51, 545–556. [Google Scholar] [CrossRef]
- Ocaña, O.H.; Córcoles, R.; Fabra, Á.; Moreno-Bueno, G.; Acloque, H.; Vega, S.; Barrallo-Gimeno, A.; Cano, A.; Nieto, M.A. Metastatic Colonization Requires the Repression of the Epithelial-Mesenchymal Transition Inducer Prrx1. Cancer Cell 2012, 22, 709–724. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; He, H.; Jiang, Q.; Wang, Y.; Jia, S. CBX6 Promotes HCC Metastasis Via Transcription Factors Snail/Zeb1-Mediated EMT Mechanism. Onco Targets Ther. 2020, 13, 12489–12500. [Google Scholar] [CrossRef]
- Morrison, C.D.; Parvani, J.G.; Schiemann, W.P. The Relevance of the TGF-β Paradox to EMT-MET Programs. Cancer Lett. 2013, 341, 30–40. [Google Scholar] [CrossRef] [Green Version]
- Tan, E.-J.; Olsson, A.-K.; Moustakas, A. Reprogramming during Epithelial to Mesenchymal Transition under the Control of TGFβ. Cell Adhes. Migr. 2015, 9, 233–246. [Google Scholar] [CrossRef] [Green Version]
- VanderVorst, K.; Dreyer, C.A.; Konopelski, S.E.; Lee, H.; Ho, H.-Y.H.; Carraway, K.L. Wnt/PCP Signaling Contribution to Carcinoma Collective Cell Migration and Metastasis. Cancer Res. 2019, 79, 1719–1729. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Tang, P.; Li, S.; Qin, X.; Yang, H.; Wu, C.; Liu, Y. Notch Signaling Pathway Networks in Cancer Metastasis: A New Target for Cancer Therapy. Med. Oncol. 2017, 34, 180. [Google Scholar] [CrossRef] [PubMed]
- Joseph, J.P.; Harishankar, M.K.; Pillai, A.A.; Devi, A. Hypoxia Induced EMT: A Review on the Mechanism of Tumor Progression and Metastasis in OSCC. Oral Oncol. 2018, 80, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Brizel, D.M.; Schroeder, T.; Scher, R.L.; Walenta, S.; Clough, R.W.; Dewhirst, M.W.; Mueller-Klieser, W. Elevated Tumor Lactate Concentrations Predict for an Increased Risk of Metastases in Head-and-Neck Cancer. Int. J. Radiat. Oncol. 2001, 51, 349–353. [Google Scholar] [CrossRef]
- Goetze, K. Lactate Enhances Motility of Tumor Cells and Inhibits Monocyte Migration and Cytokine Release. Int. J. Oncol. 2011, 39, 453–463. [Google Scholar] [CrossRef]
- Knudsen, E.S.; Ertel, A.; Davicioni, E.; Kline, J.; Schwartz, G.F.; Witkiewicz, A.K. Progression of Ductal Carcinoma in Situ to Invasive Breast Cancer Is Associated with Gene Expression Programs of EMT and Myoepithelia. Breast Cancer Res. Treat. 2012, 133, 1009–1024. [Google Scholar] [CrossRef]
- Thiery, J.P. Epithelial–Mesenchymal Transitions in Tumour Progression. Nat. Rev. Cancer 2002, 2, 442–454. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, B.P. Epithelial-Mesenchymal Transition in Breast Cancer Progression and Metastasis. Chin. J. Cancer 2011, 30, 603–611. [Google Scholar] [CrossRef]
- Roh, M.R.; Zheng, Z.; Kim, H.S.; Kwon, J.E.; Jeung, H.-C.; Rha, S.Y.; Chung, K.Y. Differential Expression Patterns of MMPs and Their Role in the Invasion of Epithelial Premalignant Tumors and Invasive Cutaneous Squamous Cell Carcinoma. Exp. Mol. Pathol. 2012, 92, 236–242. [Google Scholar] [CrossRef]
- Rahman, M.; Mohammed, S. Breast Cancer Metastasis and the Lymphatic System. Oncol. Lett. 2015, 10, 1233–1239. [Google Scholar] [CrossRef]
- Wong, A.D.; Searson, P.C. Live-Cell Imaging of Invasion and Intravasation in an Artificial Microvessel Platform. Cancer Res. 2014, 74, 4937–4945. [Google Scholar] [CrossRef] [Green Version]
- Bolós, V.; Mira, E.; Martínez-Poveda, B.; Luxán, G.; Cañamero, M.; Martínez-A, C.; Mañes, S.; de la Pompa, J.L. Notch Activation Stimulates Migration of Breast Cancer Cells and Promotes Tumor Growth. Breast Cancer Res. 2013, 15, R54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonoshita, M.; Aoki, M.; Fuwa, H.; Aoki, K.; Hosogi, H.; Sakai, Y.; Hashida, H.; Takabayashi, A.; Sasaki, M.; Robine, S.; et al. Suppression of Colon Cancer Metastasis by Aes through Inhibition of Notch Signaling. Cancer Cell 2011, 19, 125–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khuon, S.; Liang, L.; Dettman, R.W.; Sporn, P.H.S.; Wysolmerski, R.B.; Chew, T.-L. Myosin Light Chain Kinase Mediates Transcellular Intravasation of Breast Cancer Cells through the Underlying Endothelial Cells: A Three-Dimensional FRET Study. J. Cell Sci. 2010, 123, 431–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arvanitis, C.; Khuon, S.; Spann, R.; Ridge, K.M.; Chew, T.-L. Structure and Biomechanics of the Endothelial Transcellular Circumferential Invasion Array in Tumor Invasion. PLoS ONE 2014, 9, e89758. [Google Scholar] [CrossRef] [Green Version]
- Labelle, M.; Hynes, R.O. The Initial Hours of Metastasis: The Importance of Cooperative Host–Tumor Cell Interactions during Hematogenous Dissemination. Cancer Discov. 2012, 2, 1091–1099. [Google Scholar] [CrossRef] [Green Version]
- Mamessier, E.; Sylvain, A.; Thibult, M.-L.; Houvenaeghel, G.; Jacquemier, J.; Castellano, R.; Gonçalves, A.; André, P.; Romagné, F.; Thibault, G.; et al. Human Breast Cancer Cells Enhance Self Tolerance by Promoting Evasion from NK Cell Antitumor Immunity. J. Clin. Investig. 2011, 121, 3609–3622. [Google Scholar] [CrossRef] [Green Version]
- Moose, D.L.; Krog, B.L.; Kim, T.-H.; Zhao, L.; Williams-Perez, S.; Burke, G.; Rhodes, L.; Vanneste, M.; Breheny, P.; Milhem, M.; et al. Cancer Cells Resist Mechanical Destruction in Circulation via RhoA/Actomyosin-Dependent Mechano-Adaptation. Cell Rep. 2020, 30, 3864–3874.e6. [Google Scholar] [CrossRef]
- Kopp, H.-G.; Placke, T.; Salih, H.R. Platelet-Derived Transforming Growth Factor-β Down-Regulates NKG2D Thereby Inhibiting Natural Killer Cell Antitumor Reactivity. Cancer Res. 2009, 69, 7775–7783. [Google Scholar] [CrossRef] [Green Version]
- Kienast, Y.; von Baumgarten, L.; Fuhrmann, M.; Klinkert, W.E.F.; Goldbrunner, R.; Herms, J.; Winkler, F. Real-Time Imaging Reveals the Single Steps of Brain Metastasis Formation. Nat. Med. 2010, 16, 116–122. [Google Scholar] [CrossRef]
- Valiente, M.; Obenauf, A.C.; Jin, X.; Chen, Q.; Zhang, X.H.-F.; Lee, D.J.; Chaft, J.E.; Kris, M.G.; Huse, J.T.; Brogi, E.; et al. Serpins Promote Cancer Cell Survival and Vascular Co-Option in Brain Metastasis. Cell 2014, 156, 1002–1016. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Kosaka, A.; Ikeura, M.; Kohanbash, G.; Fellows-Mayle, W.; Snyder, L.A.; Okada, H. Premetastatic Soil and Prevention of Breast Cancer Brain Metastasis. Neuro-Oncol. 2013, 15, 891–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanibuchi, M.; Kim, S.-J.; Fidler, I.J.; Nishioka, Y. The Molecular Biology of Lung Cancer Brain Metastasis: An Overview of Current Comprehensions and Future Perspectives. J. Med. Investig. 2014, 61, 241–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soto, M.S.; Serres, S.; Anthony, D.C.; Sibson, N.R. Functional Role of Endothelial Adhesion Molecules in the Early Stages of Brain Metastasis. Neuro-Oncol. 2014, 16, 540–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, K.; Fukuda, K.; Xing, F.; Zhang, Y.; Sharma, S.; Liu, Y.; Chan, M.D.; Zhou, X.; Qasem, S.A.; Pochampally, R.; et al. Roles of the Cyclooxygenase 2 Matrix Metalloproteinase 1 Pathway in Brain Metastasis of Breast Cancer. J. Biol. Chem. 2015, 290, 9842–9854. [Google Scholar] [CrossRef] [Green Version]
- Gunasinghe, N.P.A.D.; Wells, A.; Thompson, E.W.; Hugo, H.J. Mesenchymal–Epithelial Transition (MET) as a Mechanism for Metastatic Colonisation in Breast Cancer. Cancer Metastasis Rev. 2012, 31, 469–478. [Google Scholar] [CrossRef]
- Yoshida, T.; Ozawa, Y.; Kimura, T.; Sato, Y.; Kuznetsov, G.; Xu, S.; Uesugi, M.; Agoulnik, S.; Taylor, N.; Funahashi, Y.; et al. Eribulin Mesilate Suppresses Experimental Metastasis of Breast Cancer Cells by Reversing Phenotype from Epithelial–Mesenchymal Transition (EMT) to Mesenchymal–Epithelial Transition (MET) States. Br. J. Cancer 2014, 110, 1497–1505. [Google Scholar] [CrossRef]
- Chao, Y.; Wu, Q.; Acquafondata, M.; Dhir, R.; Wells, A. Partial Mesenchymal to Epithelial Reverting Transition in Breast and Prostate Cancer Metastases. Cancer Microenviron. 2012, 5, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Wanner, I.B.; Anderson, M.A.; Song, B.; Levine, J.; Fernandez, A.; Gray-Thompson, Z.; Ao, Y.; Sofroniew, M.V. Glial Scar Borders Are Formed by Newly Proliferated, Elongated Astrocytes That Interact to Corral Inflammatory and Fibrotic Cells via STAT3-Dependent Mechanisms after Spinal Cord Injury. J. Neurosci. 2013, 33, 12870–12886. [Google Scholar] [CrossRef] [Green Version]
- Lorger, M.; Felding-Habermann, B. Capturing Changes in the Brain Microenvironment during Initial Steps of Breast Cancer Brain Metastasis. Am. J. Pathol. 2010, 176, 2958–2971. [Google Scholar] [CrossRef]
- Fitzgerald, D.P.; Palmieri, D.; Hua, E.; Hargrave, E.; Herring, J.M.; Qian, Y.; Vega-Valle, E.; Weil, R.J.; Stark, A.M.; Vortmeyer, A.O.; et al. Reactive Glia Are Recruited by Highly Proliferative Brain Metastases of Breast Cancer and Promote Tumor Cell Colonization. Clin. Exp. Metastasis 2008, 25, 799–810. [Google Scholar] [CrossRef] [Green Version]
- Xing, F.; Kobayashi, A.; Okuda, H.; Watabe, M.; Pai, S.K.; Pandey, P.R.; Hirota, S.; Wilber, A.; Mo, Y.; Moore, B.E.; et al. Reactive Astrocytes Promote the Metastatic Growth of Breast Cancer Stem-like Cells by Activating Notch Signalling in Brain. EMBO Mol. Med. 2013, 5, 384–396. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Xu, S.; Xin, Y.; Yu, S.; Ping, Y.; Chen, L.; Xiao, H.; Wang, B.; Yi, L.; Wang, Q.; et al. Tumor-Associated Microglia/Macrophages Enhance the Invasion of Glioma Stem-like Cells via TGF-Β1 Signaling Pathway. J. Immunol. 2012, 189, 444–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demeule, M.; Bertrand, Y.; Michaud-Levesque, J.; Jodoin, J.; Rolland, Y.; Gabathuler, R.; Béliveau, R. Regulation of Plasminogen Activation: A Role for Melanotransferrin (P97) in Cell Migration. Blood 2003, 102, 1723–1731. [Google Scholar] [CrossRef] [Green Version]
- Dunn, L.L.; Sekyere, E.O.; Suryo Rahmanto, Y.; Richardson, D.R. The Function of Melanotransferrin: A Role in Melanoma Cell Proliferation and Tumorigenesis. Carcinogenesis 2006, 27, 2157–2169. [Google Scholar] [CrossRef]
- Kim, S.W.; Choi, H.J.; Lee, H.-J.; He, J.; Wu, Q.; Langley, R.R.; Fidler, I.J.; Kim, S.-J. Role of the Endothelin Axis in Astrocyte- and Endothelial Cell-Mediated Chemoprotection of Cancer Cells. Neuro-Oncol. 2014, 16, 1585–1598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Boire, A.; Jin, X.; Valiente, M.; Er, E.E.; Lopez-Soto, A.; Jacob, L.S.; Patwa, R.; Shah, H.; Xu, K.; et al. Carcinoma–Astrocyte Gap Junctions Promote Brain Metastasis by CGAMP Transfer. Nature 2016, 533, 493–498. [Google Scholar] [CrossRef] [Green Version]
- Neman, J.; Termini, J.; Wilczynski, S.; Vaidehi, N.; Choy, C.; Kowolik, C.M.; Li, H.; Hambrecht, A.C.; Roberts, E.; Jandial, R. Human Breast Cancer Metastases to the Brain Display GABAergic Properties in the Neural Niche. Proc. Natl. Acad. Sci. USA 2014, 111, 984–989. [Google Scholar] [CrossRef] [Green Version]
- Grupenmacher, A.T.; Halpern, A.L.; Bonaldo, M.d.F.; Huang, C.-C.; Hamm, C.A.; de Andrade, A.; Tomita, T.; Sredni, S.T. Study of the Gene Expression and MicroRNA Expression Profiles of Malignant Rhabdoid Tumors Originated in the Brain (AT/RT) and in the Kidney (RTK). Childs Nerv. Syst. 2013, 29, 1977–1983. [Google Scholar] [CrossRef]
- Huarte, M.; Rinn, J.L. Large Non-Coding RNAs: Missing Links in Cancer? Hum. Mol. Genet. 2010, 19, R152–R161. [Google Scholar] [CrossRef] [Green Version]
- International Human Genome Sequencing Consortium. Initial Sequencing and Analysis of the Human Genome. Nature 2001, 409, 860–921. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Getz, G.; Miska, E.A.; Alvarez-Saavedra, E.; Lamb, J.; Peck, D.; Sweet-Cordero, A.; Ebert, B.L.; Mak, R.H.; Ferrando, A.A.; et al. MicroRNA Expression Profiles Classify Human Cancers. Nature 2005, 435, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Volinia, S.; Calin, G.A.; Liu, C.-G.; Ambs, S.; Cimmino, A.; Petrocca, F.; Visone, R.; Iorio, M.; Roldo, C.; Ferracin, M.; et al. A MicroRNA Expression Signature of Human Solid Tumors Defines Cancer Gene Targets. Proc. Natl. Acad. Sci. USA 2006, 103, 2257–2261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacRae, I.J.; Zhou, K.; Li, F.; Repic, A.; Brooks, A.N.; Cande, W.Z.; Adams, P.D.; Doudna, J.A. Structural Basis for Double-Stranded RNA Processing by Dicer. Science 2006, 311, 195–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, H.; Tomari, Y. RISC Assembly: Coordination between Small RNAs and Argonaute Proteins. Biochim. Biophys. Acta BBA Gene Regul. Mech. 2016, 1859, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Chendrimada, T.P.; Gregory, R.I.; Kumaraswamy, E.; Norman, J.; Cooch, N.; Nishikura, K.; Shiekhattar, R. TRBP Recruits the Dicer Complex to Ago2 for MicroRNA Processing and Gene Silencing. Nature 2005, 436, 740–744. [Google Scholar] [CrossRef]
- Meister, G.; Tuschl, T. Mechanisms of Gene Silencing by Double-Stranded RNA. Nature 2004, 431, 343–349. [Google Scholar] [CrossRef]
- Eichhorn, S.W.; Guo, H.; McGeary, S.E.; Rodriguez-Mias, R.A.; Shin, C.; Baek, D.; Hsu, S.; Ghoshal, K.; Villén, J.; Bartel, D.P. MRNA Destabilization Is the Dominant Effect of Mammalian MicroRNAs by the Time Substantial Repression Ensues. Mol. Cell 2014, 56, 104–115. [Google Scholar] [CrossRef] [Green Version]
- Lewis, B.P.; Shih, I.; Jones-Rhoades, M.W.; Bartel, D.P.; Burge, C.B. Prediction of Mammalian MicroRNA Targets. Cell 2003, 115, 787–798. [Google Scholar] [CrossRef] [Green Version]
- Tay, Y.; Zhang, J.; Thomson, A.M.; Lim, B.; Rigoutsos, I. MicroRNAs to Nanog, Oct4 and Sox2 Coding Regions Modulate Embryonic Stem Cell Differentiation. Nature 2008, 455, 1124–1128. [Google Scholar] [CrossRef]
- Miska, E.A.; Alvarez-Saavedra, E.; Abbott, A.L.; Lau, N.C.; Hellman, A.B.; McGonagle, S.M.; Bartel, D.P.; Ambros, V.R.; Horvitz, H.R. Most Caenorhabditis Elegans MicroRNAs Are Individually Not Essential for Development or Viability. PLoS Genet. 2007, 3, e215. [Google Scholar] [CrossRef] [Green Version]
- Bernstein, E.; Kim, S.Y.; Carmell, M.A.; Murchison, E.P.; Alcorn, H.; Li, M.Z.; Mills, A.A.; Elledge, S.J.; Anderson, K.V.; Hannon, G.J. Dicer Is Essential for Mouse Development. Nat. Genet. 2003, 35, 215–217. [Google Scholar] [CrossRef] [PubMed]
- Vidigal, J.A.; Ventura, A. The Biological Functions of MiRNAs: Lessons from in Vivo Studies. Trends Cell Biol. 2015, 25, 137–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Zhang, Z.; O’Loughlin, E.; Wang, L.; Fan, X.; Lai, E.C.; Yi, R. MicroRNA-205 Controls Neonatal Expansion of Skin Stem Cells by Modulating the PI(3)K Pathway. Nat. Cell Biol. 2013, 15, 1153–1163. [Google Scholar] [CrossRef] [PubMed]
- Ventura, A.; Young, A.G.; Winslow, M.M.; Lintault, L.; Meissner, A.; Erkeland, S.J.; Newman, J.; Bronson, R.T.; Crowley, D.; Stone, J.R.; et al. Targeted Deletion Reveals Essential and Overlapping Functions of the MiR-17∼92 Family of MiRNA Clusters. Cell 2008, 132, 875–886. [Google Scholar] [CrossRef] [Green Version]
- Calin, G.A.; Sevignani, C.; Dumitru, C.D.; Hyslop, T.; Noch, E.; Yendamuri, S.; Shimizu, M.; Rattan, S.; Bullrich, F.; Negrini, M.; et al. Human MicroRNA Genes Are Frequently Located at Fragile Sites and Genomic Regions Involved in Cancers. Proc. Natl. Acad. Sci. USA 2004, 101, 2999–3004. [Google Scholar] [CrossRef] [Green Version]
- Wan, L.; Pantel, K.; Kang, Y. Tumor Metastasis: Moving New Biological Insights into the Clinic. Nat. Med. 2013, 19, 1450–1464. [Google Scholar] [CrossRef]
- Sun, Y.; Ma, L. The Emerging Molecular Machinery and Therapeutic Targets of Metastasis. Trends Pharmacol. Sci. 2015, 36, 349–359. [Google Scholar] [CrossRef] [Green Version]
- Kanchan, R.K.; Siddiqui, J.A.; Mahapatra, S.; Batra, S.K.; Nasser, M.W. MicroRNAs Orchestrate Pathophysiology of Breast Cancer Brain Metastasis: Advances in Therapy. Mol. Cancer 2020, 19, 29. [Google Scholar] [CrossRef] [Green Version]
- Ma, L. Role of MiR-10b in Breast Cancer Metastasis. Breast Cancer Res. 2010, 12, 210. [Google Scholar] [CrossRef] [Green Version]
- Burk, U.; Schubert, J.; Wellner, U.; Schmalhofer, O.; Vincan, E.; Spaderna, S.; Brabletz, T. A Reciprocal Repression between ZEB1 and Members of the MiR-200 Family Promotes EMT and Invasion in Cancer Cells. EMBO Rep. 2008, 9, 582–589. [Google Scholar] [CrossRef] [Green Version]
- Brabletz, S.; Brabletz, T. The ZEB/MiR-200 Feedback Loop—A Motor of Cellular Plasticity in Development and Cancer? EMBO Rep. 2010, 11, 670–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kundu, S.T.; Byers, L.A.; Peng, D.H.; Roybal, J.D.; Diao, L.; Wang, J.; Tong, P.; Creighton, C.J.; Gibbons, D.L. The MiR-200 Family and the MiR-183~96~182 Cluster Target Foxf2 to Inhibit Invasion and Metastasis in Lung Cancers. Oncogene 2016, 35, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Park, S.I.; McCauley, L.K.; Wang, C.-Y. Signaling between Transforming Growth Factor β (TGF-β) and Transcription Factor SNAI2 Represses Expression of MicroRNA MiR-203 to Promote Epithelial-Mesenchymal Transition and Tumor Metastasis. J. Biol. Chem. 2013, 288, 10241–10253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, S.-J.; Hu, J.-Y.; Kuang, X.-Y.; Luo, J.-M.; Hou, Y.-F.; Di, G.-H.; Wu, J.; Shen, Z.-Z.; Song, H.-Y.; Shao, Z.-M. MicroRNA-200a Promotes Anoikis Resistance and Metastasis by Targeting YAP1 in Human Breast Cancer. Clin. Cancer Res. 2013, 19, 1389–1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansoori, B.; Mohammadi, A.; Ghasabi, M.; Shirjang, S.; Dehghan, R.; Montazeri, V.; Holmskov, U.; Kazemi, T.; Duijf, P.; Gjerstorff, M.; et al. MiR-142-3p as Tumor Suppressor MiRNA in the Regulation of Tumorigenicity, Invasion and Migration of Human Breast Cancer by Targeting Bach-1 Expression. J. Cell. Physiol. 2019, 234, 9816–9825. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Rokavec, M.; Jiang, L.; Horst, D.; Hermeking, H. Antagonistic Effects of P53 and HIF1A on MicroRNA-34a Regulation of PPP1R11 and STAT3 and Hypoxia-Induced Epithelial to Mesenchymal Transition in Colorectal Cancer Cells. Gastroenterology 2017, 153, 505–520. [Google Scholar] [CrossRef]
- Pencheva, N.; Tran, H.; Buss, C.; Huh, D.; Drobnjak, M.; Busam, K.; Tavazoie, S.F. Convergent Multi-MiRNA Targeting of ApoE Drives LRP1/LRP8-Dependent Melanoma Metastasis and Angiogenesis. Cell 2012, 151, 1068–1082. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Li, W.; Wen, C.; Diao, Y.; Zhao, T. MicroRNA-214 Promotes the EMT Process in Melanoma by Downregulating CADM1 Expression. Mol. Med. Rep. 2020, 22, 3795–3803. [Google Scholar] [CrossRef]
- Penna, E.; Orso, F.; Cimino, D.; Tenaglia, E.; Lembo, A.; Quaglino, E.; Poliseno, L.; Haimovic, A.; Osella-Abate, S.; De Pittà, C.; et al. MicroRNA-214 Contributes to Melanoma Tumour Progression through Suppression of TFAP2C: MiR-214 and Melanoma Progression. EMBO J. 2011, 30, 1990–2007. [Google Scholar] [CrossRef] [Green Version]
- Cantini, L.; Bertoli, G.; Cava, C.; Dubois, T.; Zinovyev, A.; Caselle, M.; Castiglioni, I.; Barillot, E.; Martignetti, L. Identification of MicroRNA Clusters Cooperatively Acting on Epithelial to Mesenchymal Transition in Triple Negative Breast Cancer. Nucleic Acids Res. 2019, 47, 2205–2215. [Google Scholar] [CrossRef] [Green Version]
- Lv, Z.-D.; Yang, D.-X.; Liu, X.-P.; Jin, L.-Y.; Wang, X.-G.; Yang, Z.-C.; Liu, D.; Zhao, J.-J.; Kong, B.; Li, F.-N.; et al. MiR-212-5p Suppresses the Epithelial-Mesenchymal Transition in Triple-Negative Breast Cancer by Targeting Prrx2. Cell. Physiol. Biochem. 2017, 44, 1785–1795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Zhao, Y.; He, Y.; Mao, Y. MiR-19b Promotes Breast Cancer Metastasis through Targeting MYLIP and Its Related Cell Adhesion Molecules. Oncotarget 2017, 8, 64330–64343. [Google Scholar] [CrossRef]
- Zhang, L.; Sullivan, P.S.; Goodman, J.C.; Gunaratne, P.H.; Marchetti, D. MicroRNA-1258 Suppresses Breast Cancer Brain Metastasis by Targeting Heparanase. Cancer Res. 2011, 71, 645–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Dong, Y.; Zhu, N.; Tsoi, H.; Zhao, Z.; Wu, C.W.; Wang, K.; Zheng, S.; Ng, S.S.; Chan, F.K.; et al. MicroRNA-139-5p Exerts Tumor Suppressor Function by Targeting NOTCH1 in Colorectal Cancer. Mol. Cancer 2014, 13, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, L.; Wu, Y.; Wang, J.; He, J.; Han, X. Sevoflurane Inhibits the Migration and Invasion of Colorectal Cancer Cells through Regulating ERK/MMP-9 Pathway by up-Regulating MiR-203. Eur. J. Pharmacol. 2019, 850, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Chen, X.; Tang, Y.; Deng, Y. MicroRNA-194 Modulates Epithelial–Mesenchymal Transition in Human Colorectal Cancer Metastasis. Onco Targets Ther. 2017, 10, 1269–1278. [Google Scholar] [CrossRef] [Green Version]
- Martello, G.; Rosato, A.; Ferrari, F.; Manfrin, A.; Cordenonsi, M.; Dupont, S.; Enzo, E.; Guzzardo, V.; Rondina, M.; Spruce, T.; et al. A MicroRNA Targeting Dicer for Metastasis Control. Cell 2010, 141, 1195–1207. [Google Scholar] [CrossRef] [Green Version]
- Shao, Y.; Chen, T.; Zheng, X.; Yang, S.; Xu, K.; Chen, X.; Xu, F.; Wang, L.; Shen, Y.; Wang, T.; et al. Colorectal Cancer-Derived Small Extracellular Vesicles Establish an Inflammatory Premetastatic Niche in Liver Metastasis. Carcinogenesis 2018, 39, 1368–1379. [Google Scholar] [CrossRef]
- Wang, D.; Wang, X.; Si, M.; Yang, J.; Sun, S.; Wu, H.; Cui, S.; Qu, X.; Yu, X. Exosome-Encapsulated MiRNAs Contribute to CXCL12/CXCR4-Induced Liver Metastasis of Colorectal Cancer by Enhancing M2 Polarization of Macrophages. Cancer Lett. 2020, 474, 36–52. [Google Scholar] [CrossRef]
- Yang, M.; Chen, J.; Su, F.; Yu, B.; Su, F.; Lin, L.; Liu, Y.; Huang, J.-D.; Song, E. Microvesicles Secreted by Macrophages Shuttle Invasion-Potentiating MicroRNAs into Breast Cancer Cells. Mol. Cancer 2011, 10, 117. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Fong, M.Y.; Min, Y.; Somlo, G.; Liu, L.; Palomares, M.R.; Yu, Y.; Chow, A.; O’Connor, S.T.F.; Chin, A.R.; et al. Cancer-Secreted MiR-105 Destroys Vascular Endothelial Barriers to Promote Metastasis. Cancer Cell 2014, 25, 501–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegel, R.; Ward, E.; Brawley, O.; Jemal, A. Cancer Statistics, 2011: The Impact of Eliminating Socioeconomic and Racial Disparities on Premature Cancer Deaths. CA. Cancer J. Clin. 2011, 61, 212–236. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2016: Cancer Statistics, 2016. CA. Cancer J. Clin. 2016, 66, 7–30. [Google Scholar] [CrossRef] [Green Version]
- Mujoomdar, A.; Austin, J.H.M.; Malhotra, R.; Powell, C.A.; Pearson, G.D.N.; Shiau, M.C.; Raftopoulos, H. Clinical Predictors of Metastatic Disease to the Brain from Non–Small Cell Lung Carcinoma: Primary Tumor Size, Cell Type, and Lymph Node Metastases. Radiology 2007, 242, 882–888. [Google Scholar] [CrossRef] [PubMed]
- Budczies, J.; von Winterfeld, M.; Klauschen, F.; Bockmayr, M.; Lennerz, J.K.; Denkert, C.; Wolf, T.; Warth, A.; Dietel, M.; Anagnostopoulos, I.; et al. The Landscape of Metastatic Progression Patterns across Major Human Cancers. Oncotarget 2015, 6, 570–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sørensen, J.B.; Hansen, H.H.; Hansen, M.; Dombernowsky, P. Brain Metastases in Adenocarcinoma of the Lung: Frequency, Risk Groups, and Prognosis. J. Clin. Oncol. 1988, 6, 1474–1480. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Li, Q.; Xu, M.; Qi, Z. Effect of Whole-Brain and Intensity-Modulated Radiotherapy on Serum Levels of MiR-21 and Prognosis for Lung Cancer Metastatic to the Brain. Med. Sci. Monit. 2020, 26, e924640. [Google Scholar] [CrossRef]
- Dong, J.; Zhang, Z.; Gu, T.; Xu, S.-F.; Dong, L.-X.; Li, X.; Fu, B.-H.; Fu, Z.-Z. The Role of MicroRNA-21 in Predicting Brain Metastases from Non-Small Cell Lung Cancer. Onco Targets Ther. 2016, 10, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Singh, M.; Garg, N.; Venugopal, C.; Hallett, R.; Tokar, T.; McFarlane, N.; Mahendram, S.; Bakhshinyan, D.; Manoranjan, B.; Vora, P.; et al. STAT3 Pathway Regulates Lung-Derived Brain Metastasis Initiating Cell Capacity through MiR-21 Activation. Oncotarget 2015, 6, 27461–27477. [Google Scholar] [CrossRef] [Green Version]
- Subramani, A.; Alsidawi, S.; Jagannathan, S.; Sumita, K.; Sasaki, A.T.; Aronow, B.; Warnick, R.E.; Lawler, S.; Driscoll, J.J. The Brain Microenvironment Negatively Regulates MiRNA-768-3p to Promote K-Ras Expression and Lung Cancer Metastasis. Sci. Rep. 2013, 3, 2392. [Google Scholar] [CrossRef] [Green Version]
- Choi, K.H.; Shin, C.H.; Lee, W.J.; Ji, H.; Kim, H.H. Dual-Strand Tumor Suppressor MiR-193b-3p and -5p Inhibit Malignant Phenotypes of Lung Cancer by Suppressing Their Common Targets. Biosci. Rep. 2019, 39, BSR20190634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, C.; Zhao, H.; Yang, B.; Sun, Z.; Li, X.; Hu, X. Lnc-REG3G-3-1/MiR-215-3p Promotes Brain Metastasis of Lung Adenocarcinoma by Regulating Leptin and SLC2A5. Front. Oncol. 2020, 10, 1344. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Hou, L.; Wei, J.; Du, Y.; Zhao, Y.; Deng, X.; Lin, X. Hsa-MiR-217 Inhibits the Proliferation, Migration, and Invasion in Non-Small Cell Lung Cancer Cells Via Targeting SIRT1 and P53/KAI1 Signaling. Balk. Med. J. 2021, 37, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Donzelli, S.; Mori, F.; Bellissimo, T.; Sacconi, A.; Casini, B.; Frixa, T.; Roscilli, G.; Aurisicchio, L.; Facciolo, F.; Pompili, A.; et al. Epigenetic Silencing of MiR-145-5p Contributes to Brain Metastasis. Oncotarget 2015, 6, 35183–35201. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Xu, Y.; Zhang, Y.; Tan, W.; Xue, J.; Yang, Z.; Zhang, Y.; Lu, Y.; Hu, X. Downregulation of MiR-145 Contributes to Lung Adenocarcinoma Cell Growth to Form Brain Metastases. Oncol. Rep. 2013, 30, 2027–2034. [Google Scholar] [CrossRef] [Green Version]
- Hwang, S.J.; Lee, H.W.; Kim, H.R.; Song, H.J.; Lee, D.H.; Lee, H.; Shin, C.H.; Joung, J.-G.; Kim, D.-H.; Joo, K.M.; et al. Overexpression of MicroRNA-95-3p Suppresses Brain Metastasis of Lung Adenocarcinoma through Downregulation of Cyclin D1. Oncotarget 2015, 6, 20434–20448. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Xu, S.; Xu, H.; Zhang, J.; Ning, J.; Wang, S. MicroRNA-378 Is Associated with Non-Small Cell Lung Cancer Brain Metastasis by Promoting Cell Migration, Invasion and Tumor Angiogenesis. Med. Oncol. 2012, 29, 1673–1680. [Google Scholar] [CrossRef]
- Arora, S.; Ranade, A.R.; Tran, N.L.; Nasser, S.; Sridhar, S.; Korn, R.L.; Ross, J.T.D.; Dhruv, H.; Foss, K.M.; Sibenaller, Z.; et al. MicroRNA-328 Is Associated with (Non-Small) Cell Lung Cancer (NSCLC) Brain Metastasis and Mediates NSCLC Migration. Int. J. Cancer 2011, 129, 2621–2631. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Deng, Q.; Lv, Z.; Ling, Y.; Hou, X.; Chen, Z.; Dinglin, X.; Ma, S.; Li, D.; Wu, Y.; et al. N6-Methyladenosine Induced MiR-143-3p Promotes the Brain Metastasis of Lung Cancer via Regulation of VASH1. Mol. Cancer 2019, 18, 181. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.-K.; Liu, H.-F.; Ding, Y.; Gao, G.-D. Predictive Value of MicroRNA Let-7a Expression for Efficacy and Prognosis of Radiotherapy in Patients with Lung Cancer Brain Metastasis: A Case–Control Study. Medicine 2018, 97, e12847. [Google Scholar] [CrossRef]
- Wei, C.; Zhang, R.; Cai, Q.; Gao, X.; Tong, F.; Dong, J.; Hu, Y.; Wu, G.; Dong, X. MicroRNA-330-3p Promotes Brain Metastasis and Epithelial-Mesenchymal Transition via GRIA3 in Non-Small Cell Lung Cancer. Aging 2019, 11, 6734–6761. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Li, X.; Zhao, Y.; Liu, W.; Wu, H.; Liu, J.; Mu, X.; Wu, H. Down-Regulated MicroRNA-375 Expression as a Predictive Biomarker in Non-Small Cell Lung Cancer Brain Metastasis and Its Prognostic Significance. Pathol. Res. Pract. 2017, 213, 882–888. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Deng, S.; Li, L.; Liu, T.; Zhang, T.; Li, J.; Yu, Y.; Xu, Y. TGF-Β1-Mediated Exosomal Lnc-MMP2-2 Increases Blood–Brain Barrier Permeability via the MiRNA-1207-5p/EPB41L5 Axis to Promote Non-Small Cell Lung Cancer Brain Metastasis. Cell Death Dis. 2021, 12, 721. [Google Scholar] [CrossRef] [PubMed]
- Mouttet, D.; Laé, M.; Caly, M.; Gentien, D.; Carpentier, S.; Peyro-Saint-Paul, H.; Vincent-Salomon, A.; Rouzier, R.; Sigal-Zafrani, B.; Sastre-Garau, X.; et al. Estrogen-Receptor, Progesterone-Receptor and HER2 Status Determination in Invasive Breast Cancer. Concordance between Immuno-Histochemistry and MapQuant™ Microarray Based Assay. PLoS ONE 2016, 11, e0146474. [Google Scholar] [CrossRef] [Green Version]
- Berman, A.T.; Thukral, A.D.; Hwang, W.-T.; Solin, L.J.; Vapiwala, N. Incidence and Patterns of Distant Metastases for Patients With Early-Stage Breast Cancer After Breast Conservation Treatment. Clin. Breast Cancer 2013, 13, 88–94. [Google Scholar] [CrossRef]
- Saha, A.; Ghosh, S.; Roy, C.; Choudhury, K.; Chakrabarty, B.; Sarkar, R. Demographic and Clinical Profile of Patients with Brain Metastases: A Retrospective Study. Asian J. Neurosurg. 2013, 8, 157. [Google Scholar] [CrossRef] [Green Version]
- Wilhelm, I.; Molnár, J.; Fazakas, C.; Haskó, J.; Krizbai, I. Role of the Blood-Brain Barrier in the Formation of Brain Metastases. Int. J. Mol. Sci. 2013, 14, 1383–1411. [Google Scholar] [CrossRef] [Green Version]
- Kennecke, H.; Yerushalmi, R.; Woods, R.; Cheang, M.C.U.; Voduc, D.; Speers, C.H.; Nielsen, T.O.; Gelmon, K. Metastatic Behavior of Breast Cancer Subtypes. J. Clin. Oncol. 2010, 28, 3271–3277. [Google Scholar] [CrossRef]
- Sereno, M.; Haskó, J.; Molnár, K.; Medina, S.J.; Reisz, Z.; Malhó, R.; Videira, M.; Tiszlavicz, L.; Booth, S.A.; Wilhelm, I.; et al. Downregulation of Circulating MiR 802-5p and MiR 194-5p and Upregulation of Brain MEF2C along Breast Cancer Brain Metastasization. Mol. Oncol. 2020, 14, 520–538. [Google Scholar] [CrossRef] [Green Version]
- Figueira, I.; Godinho-Pereira, J.; Galego, S.; Maia, J.; Haskó, J.; Molnár, K.; Malhó, R.; Costa-Silva, B.; Wilhelm, I.; Krizbai, I.A.; et al. MicroRNAs and Extracellular Vesicles as Distinctive Biomarkers of Precocious and Advanced Stages of Breast Cancer Brain Metastases Development. Int. J. Mol. Sci. 2021, 22, 5214. [Google Scholar] [CrossRef]
- Debeb, B.G.; Lacerda, L.; Anfossi, S.; Diagaradjane, P.; Chu, K.; Bambhroliya, A.; Huo, L.; Wei, C.; Larson, R.A.; Wolfe, A.R.; et al. MiR-141-Mediated Regulation of Brain Metastasis From Breast Cancer. J. Natl. Cancer Inst. 2016, 108, djw026. [Google Scholar] [CrossRef] [Green Version]
- Okuda, H.; Xing, F.; Pandey, P.R.; Sharma, S.; Watabe, M.; Pai, S.K.; Mo, Y.-Y.; Iiizumi-Gairani, M.; Hirota, S.; Liu, Y.; et al. MiR-7 Suppresses Brain Metastasis of Breast Cancer Stem-Like Cells By Modulating KLF4. Cancer Res. 2013, 73, 1434–1444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, S.J.; Seol, H.J.; Park, Y.M.; Kim, K.H.; Gorospe, M.; Nam, D.-H.; Kim, H.H. MicroRNA-146a Suppresses Metastatic Activity in Brain Metastasis. Mol. Cells 2012, 34, 329–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, F.; Sharma, S.; Liu, Y.; Mo, Y.-Y.; Wu, K.; Zhang, Y.-Y.; Pochampally, R.; Martinez, L.A.; Lo, H.-W.; Watabe, K. MiR-509 Suppresses Brain Metastasis of Breast Cancer Cells by Modulating RhoC and TNF-α. Oncogene 2015, 34, 4890–4900. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.-K.; Lin, C.-H.; Kuo, Y.-L.; Ger, L.-P.; Cheng, H.-C.; Yao, Y.-C.; Hsiao, M.; Lu, P.-J. MiR-211 Determines Brain Metastasis Specificity through SOX11/NGN2 Axis in Triple-Negative Breast Cancer. Oncogene 2021, 40, 1737–1751. [Google Scholar] [CrossRef]
- Tominaga, N.; Kosaka, N.; Ono, M.; Katsuda, T.; Yoshioka, Y.; Tamura, K.; Lötvall, J.; Nakagama, H.; Ochiya, T. Brain Metastatic Cancer Cells Release MicroRNA-181c-Containing Extracellular Vesicles Capable of Destructing Blood–Brain Barrier. Nat. Commun. 2015, 6, 6716. [Google Scholar] [CrossRef] [Green Version]
- Xing, F.; Liu, Y.; Wu, S.-Y.; Wu, K.; Sharma, S.; Mo, Y.-Y.; Feng, J.; Sanders, S.; Jin, G.; Singh, R.; et al. Loss of XIST in Breast Cancer Activates MSN-c-Met and Reprograms Microglia via Exosomal MiRNA to Promote Brain Metastasis. Cancer Res. 2018, 78, 4316–4330. [Google Scholar] [CrossRef] [Green Version]
- Fong, M.Y.; Zhou, W.; Liu, L.; Alontaga, A.Y.; Chandra, M.; Ashby, J.; Chow, A.; O’Connor, S.T.F.; Li, S.; Chin, A.R.; et al. Breast-Cancer-Secreted MiR-122 Reprograms Glucose Metabolism in Premetastatic Niche to Promote Metastasis. Nat. Cell Biol. 2015, 17, 183–194. [Google Scholar] [CrossRef] [Green Version]
- Abbas, O.; Miller, D.D.; Bhawan, J. Cutaneous Malignant Melanoma: Update on Diagnostic and Prognostic Biomarkers. Am. J. Dermatopathol. 2014, 36, 363–379. [Google Scholar] [CrossRef] [PubMed]
- Knoll, S.; Fürst, K.; Kowtharapu, B.; Schmitz, U.; Marquardt, S.; Wolkenhauer, O.; Martin, H.; Pützer, B.M. E2F1 Induces MiR-224/452 Expression to Drive EMT through TXNIP Downregulation. EMBO Rep. 2014, 15, 1315–1329. [Google Scholar] [CrossRef] [Green Version]
- Rang, Z.; Yang, G.; Wang, Y.; Cui, F. MiR-542-3p Suppresses Invasion and Metastasis by Targeting the Proto-Oncogene Serine/Threonine Protein Kinase, PIM1, in Melanoma. Biochem. Biophys. Res. Commun. 2016, 474, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Mikkelsen, L.H.; Andersen, M.K.; Andreasen, S.; Larsen, A.-C.; Tan, Q.; Toft, P.B.; Wadt, K.; Heegaard, S. Global MicroRNA Profiling of Metastatic Conjunctival Melanoma. Melanoma Res. 2019, 29, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Bustos, M.A.; Tran, K.D.; Rahimzadeh, N.; Gross, R.; Lin, S.Y.; Shoji, Y.; Murakami, T.; Boley, C.L.; Tran, L.T.; Cole, H.; et al. Integrated Assessment of Circulating Cell-Free MicroRNA Signatures in Plasma of Patients with Melanoma Brain Metastasis. Cancers 2020, 12, 1692. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhao, H.; Yang, J.; Ma, Y.; Liu, Z.; Li, C.; Wang, T.; Yan, Z.; Du, N. MiR-150-5p Regulates Melanoma Proliferation, Invasion and Metastasis via SIX1-Mediated Warburg Effect. Biochem. Biophys. Res. Commun. 2019, 515, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Hanniford, D.; Zhong, J.; Koetz, L.; Gaziel-Sovran, A.; Lackaye, D.J.; Shang, S.; Pavlick, A.; Shapiro, R.; Berman, R.; Darvishian, F.; et al. A MiRNA-Based Signature Detected in Primary Melanoma Tissue Predicts Development of Brain Metastasis. Clin. Cancer Res. 2015, 21, 4903–4912. [Google Scholar] [CrossRef] [Green Version]
- Tan, W.-S.; Ho, K.-S.; Eu, K.-W. Brain Metastases in Colorectal Cancers. World J. Surg. 2009, 33, 817–821. [Google Scholar] [CrossRef]
- Li, Z.; Gu, X.; Fang, Y.; Xiang, J.; Chen, Z. MicroRNA Expression Profiles in Human Colorectal Cancers with Brain Metastases. Oncol. Lett. 2012, 3, 346–350. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-J.; Kim, S.-J.; Seo, H.-H.; Shin, S.-P.; Kim, D.; Park, C.-S.; Kim, K.-T.; Kim, Y.-H.; Jeong, J.-S.; Kim, I.-H. Over-Expression of MiR-145 Enhances the Effectiveness of HSVtk Gene Therapy for Malignant Glioma. Cancer Lett. 2012, 320, 72–80. [Google Scholar] [CrossRef]
- Wang, J.; Li, B.; Wang, C.; Luo, Y.; Zhao, M.; Chen, P. Long Noncoding RNA FOXD2-AS1 Promotes Glioma Cell Cycle Progression and Proliferation through the FOXD2-AS1/MiR-31/CDK1 Pathway. J. Cell. Biochem. 2019, 120, 19784–19795. [Google Scholar] [CrossRef]
- Kim, C.W.; Oh, E.-T.; Kim, J.M.; Park, J.-S.; Lee, D.H.; Lee, J.-S.; Kim, K.K.; Park, H.J. Corrigendum to “Hypoxia-Induced MicroRNA-590-5p Promotes Colorectal Cancer Progression by Modulating Matrix Metalloproteinase Activity” [Cancer Lett. 416 (2018) 31–41]. Cancer Lett. 2019, 455, 73. [Google Scholar] [CrossRef]
- Ljungberg, B.; Albiges, L.; Abu-Ghanem, Y.; Bensalah, K.; Dabestani, S.; Fernández-Pello, S.; Giles, R.H.; Hofmann, F.; Hora, M.; Kuczyk, M.A.; et al. European Association of Urology Guidelines on Renal Cell Carcinoma: The 2019 Update. Eur. Urol. 2019, 75, 799–810. [Google Scholar] [CrossRef] [PubMed]
- Jonasch, E.; Gao, J.; Rathmell, W.K. Renal Cell Carcinoma. BMJ 2014, 349, g4797. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Qi, L.; Zhang, K.; Wang, F. MicroRNA-10a Suppresses Cell Metastasis by Targeting BDNF and Predicted Patients Survival in Renal Cell Carcinoma. J. BUON Off. J. Balk. Union Oncol. 2021, 26, 250–258. [Google Scholar]
- Heinzelmann, J.; Unrein, A.; Wickmann, U.; Baumgart, S.; Stapf, M.; Szendroi, A.; Grimm, M.-O.; Gajda, M.R.; Wunderlich, H.; Junker, K. MicroRNAs with Prognostic Potential for Metastasis in Clear Cell Renal Cell Carcinoma: A Comparison of Primary Tumors and Distant Metastases. Ann. Surg. Oncol. 2014, 21, 1046–1054. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Li, H.; Zhang, Y. Downregulation of MicroRNA-206 Suppresses Clear Cell Renal Carcinoma Proliferation and Invasion by Targeting Vascular Endothelial Growth Factor A. Oncol. Rep. 2016, 35, 1778–1786. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Rao, Y.; Zhang, G.; Kong, X. MicroRNA-384 Inhibits the Growth and Invasion of Renal Cell Carcinoma Cells by Targeting Astrocyte Elevated Gene 1. Oncol. Res. Featur. Preclin. Clin. Cancer Ther. 2018, 26, 457–466. [Google Scholar] [CrossRef]
- Dong, J.-S.; Wu, B.; Zha, Z.-L. MicroRNA-588 Regulates Migration Capacity and Invasiveness of Renal Cancer Cells by Targeting EIF5A2. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 10248–10256. [Google Scholar] [CrossRef]


{kind=link}
| microRNA | Primary Tumor | Dysregulation | Role in Metastases | Origin | References |
|---|---|---|---|---|---|
| miR-21 | LC, NSCLC | Up | Promotion of cell migration, invasion, proliferation, and angiogenesis | Cancer cells | [118,119,120] |
| miR-768-3p | LC | Dow | Brain colonization via K-Ras expression enhancement | Cancer cells | [121] |
| miR-193b | LC | Down | Inhibition of cell migration and invasion | Cancer cells | [122] |
| miR-215-3p | LC | Down | Not specified | Cancer cells | [123] |
| miR-217 | LC, NSCLC | Down | Inhibition of cell viability and migration | Cancer cells | [124] |
| miR-145 | LC | Down | Inhibition of cell migration | Cancer cells | [125,126] |
| miR-95-3p | LC | Down | Inhibition of cell proliferation and invasiveness via targeting Cyclin D1 | Cancer Cells | [127] |
| miR-378 | LC, NSCLC | Up | Promotion of cell migration, invasiveness and vasculogeny mimicry | Cancer cells | [128] |
| miR-328 | LC, NSCLC | Up | Promotion of primary tumor cells migration | Cancer cells | [129] |
| miR-143-3p | LC | Up | Promotion of cell migration, invasiveness, and BBB passage | Cancer cells | [130] |
| let-7a | LC | Down | Inhibition of cell proliferation | Cancer cells | [131] |
| miR-330-3p | LC, NSCLC | Up | EMT promotion | Cancer cells | [132] |
| miR-375 | LC, NSCLC | Down | Not examined | Cancer cells | [133] |
| miR-1207-5p | LC | Down | Promoting BBB permeability | Brain microvascular endothelial cells | [134] |
| miR-802-5p | BC, TNBC | Down | Not known | Not known | [140,141] |
| miR-194-5p | BC, TNBC | Down | Supposedly promotion of passage through BBB | Cancer cells, BBB cells | [140,141] |
| miR-92a-1-5p | BC, TNBC | Up | Not examined | Cancer cells, BBB cells | [141] |
| miR-205-5p | BC, TNBC | Up | Not examined | Cancer cells | [141] |
| miR-181a-1-3p | BC, TNBC | Up | Not examined | Cancer cells, BBB cells | [141] |
| miR-141 | BC | Up | Not examined | Cancer cells | [142] |
| miR-7 | BC | Down | Inhibition of CSC self-renrewal | CSC | [143] |
| miR-146a | BC | Down | Inhibition of cell migration and invasion | Cancer cells | [144] |
| miR-509 | BC | Down | Inhibition of extravasation, BBB disruption | Upregulated in metastasizing cells | [145] |
| miR-211 | BC, TNBC | Up | Promotion of passage through BBB | Upregulated in metastasizing cells | [146] |
| miR-181c | BC | Up | BBB disruption, Extravasation | Cancer cells (secreted via exosoms) | [147] |
| miR-503 | BC | Up | Microenvironment modulation | Cancer cells (secreted via exosoms) | [148] |
| miR-19a | BC | Up | Promotion of BM growth, proliferation and inhibition of apoptosis | Astrocytes | [15] |
| miR-122 | BC | Up | Microenvironment modulation | Cancer cells (secreted via exosoms) | [149] |
| miR-224-5p/miR-452 cluster | Melanoma | Up | EMT promotion | Cancer cells | [151] |
| miR-542-3p | Melanoma | Down | EMT Inhibition | Cancer cells | [152] |
| miR-671-5p, miR-4664-3p, miR-4665-3p, miR-5694, miR-6741-3p, and miR-6796-3p | Melanoma | Up | Not Examined | Not stated | [154] |
| miR-150-5p | Melanoma | Down | Inhibition of cell proliferation, migration and invasivness | Cancer cells | [155,156] |
| miR-199a, miR-133a, miR-145, miR-143, miR-10b, miR-1, miR-199a-5p, miR-145-3p, miR-125b, miR-133b, miR-22, miR-126-5p, miR-146a, miR-28-5p, miR-576-5p, miR-199b-5p, HS_287 | CRC | Up | Not examined | Cancer cells | [158] |
| miR-31, HS_170 | CRC | Down | Not examined | Cancer cells | [158] |
| miR-590-5p | CRC | Up | Cell mobility promotion | Cancer cells | [161] |
| miR-10b, miR-30c | RCC | Down | Not examined | Cancer cells | [165] |
| miR-206 | RCC | Down | Inhibition of cell migration and invasiveness | Cancer cells | [166] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siegl, F.; Vecera, M.; Roskova, I.; Smrcka, M.; Jancalek, R.; Kazda, T.; Slaby, O.; Sana, J. The Significance of MicroRNAs in the Molecular Pathology of Brain Metastases. Cancers 2022, 14, 3386. https://doi.org/10.3390/cancers14143386
Siegl F, Vecera M, Roskova I, Smrcka M, Jancalek R, Kazda T, Slaby O, Sana J. The Significance of MicroRNAs in the Molecular Pathology of Brain Metastases. Cancers. 2022; 14(14):3386. https://doi.org/10.3390/cancers14143386
Chicago/Turabian StyleSiegl, Frantisek, Marek Vecera, Ivana Roskova, Martin Smrcka, Radim Jancalek, Tomas Kazda, Ondrej Slaby, and Jiri Sana. 2022. "The Significance of MicroRNAs in the Molecular Pathology of Brain Metastases" Cancers 14, no. 14: 3386. https://doi.org/10.3390/cancers14143386