
Reduction of LPAR1 Expression in Neuroblastoma Promotes Tumor Cell Migration

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Microarray Data Collection and Preprocessing
2.2. Functional and Pathway Enrichment Analysis of Neuroblastoma-Specific DEGs
2.3. PPI Networks
2.4. The PPI Networks and Module Selection
2.5. Cell Lines
2.6. Transient Transfection of siRNA
2.7. PCR and Real-Time PCR
2.8. Cell Proliferation Assay
2.9. Wound-Healing Assay
2.10. Cell Migration Assay
2.11. Statistical Analysis
3. Results
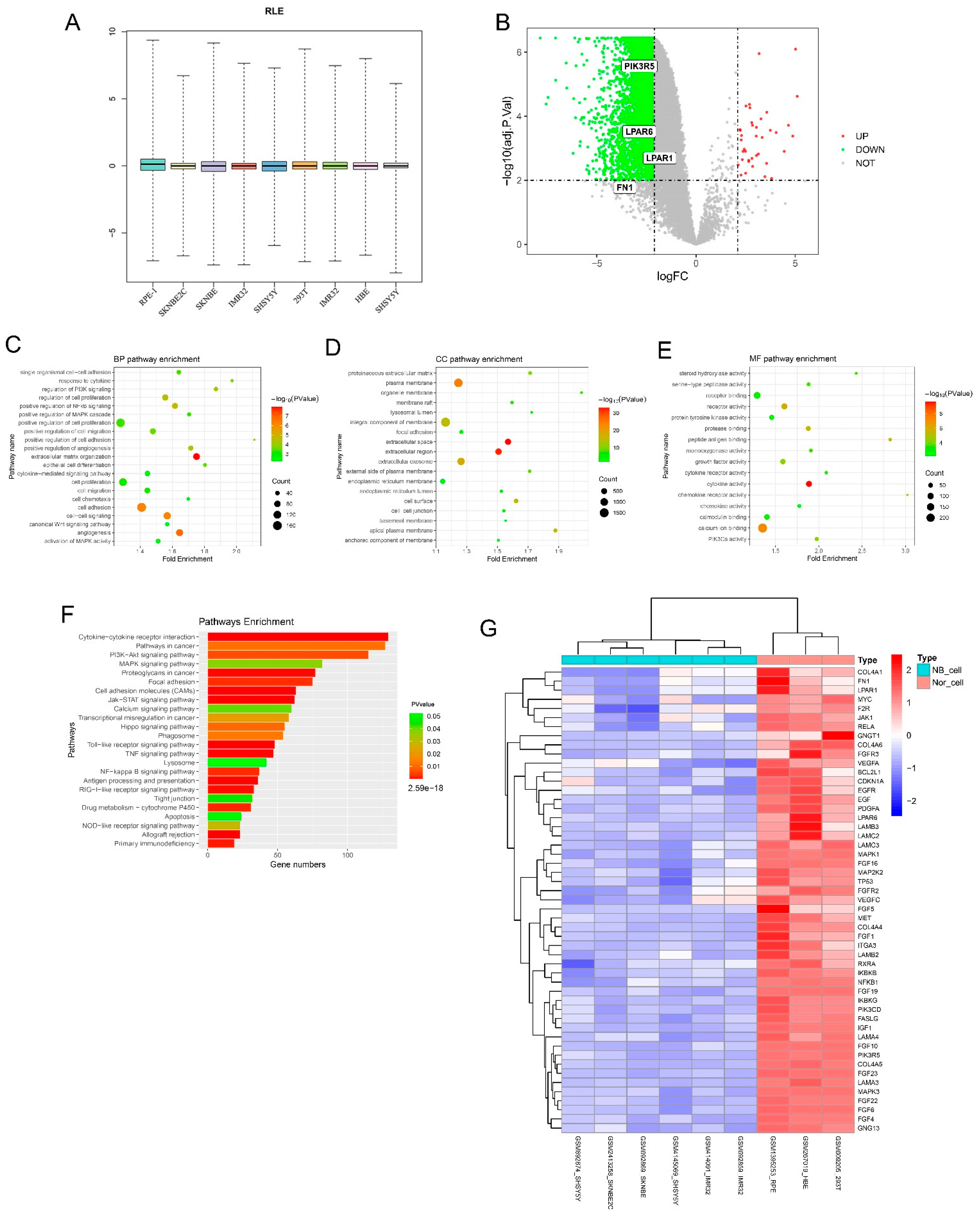
3.1. Identification of DEGs Using mRNA Microarray Data Analysis and GO/KEGG Enrichment Analysis
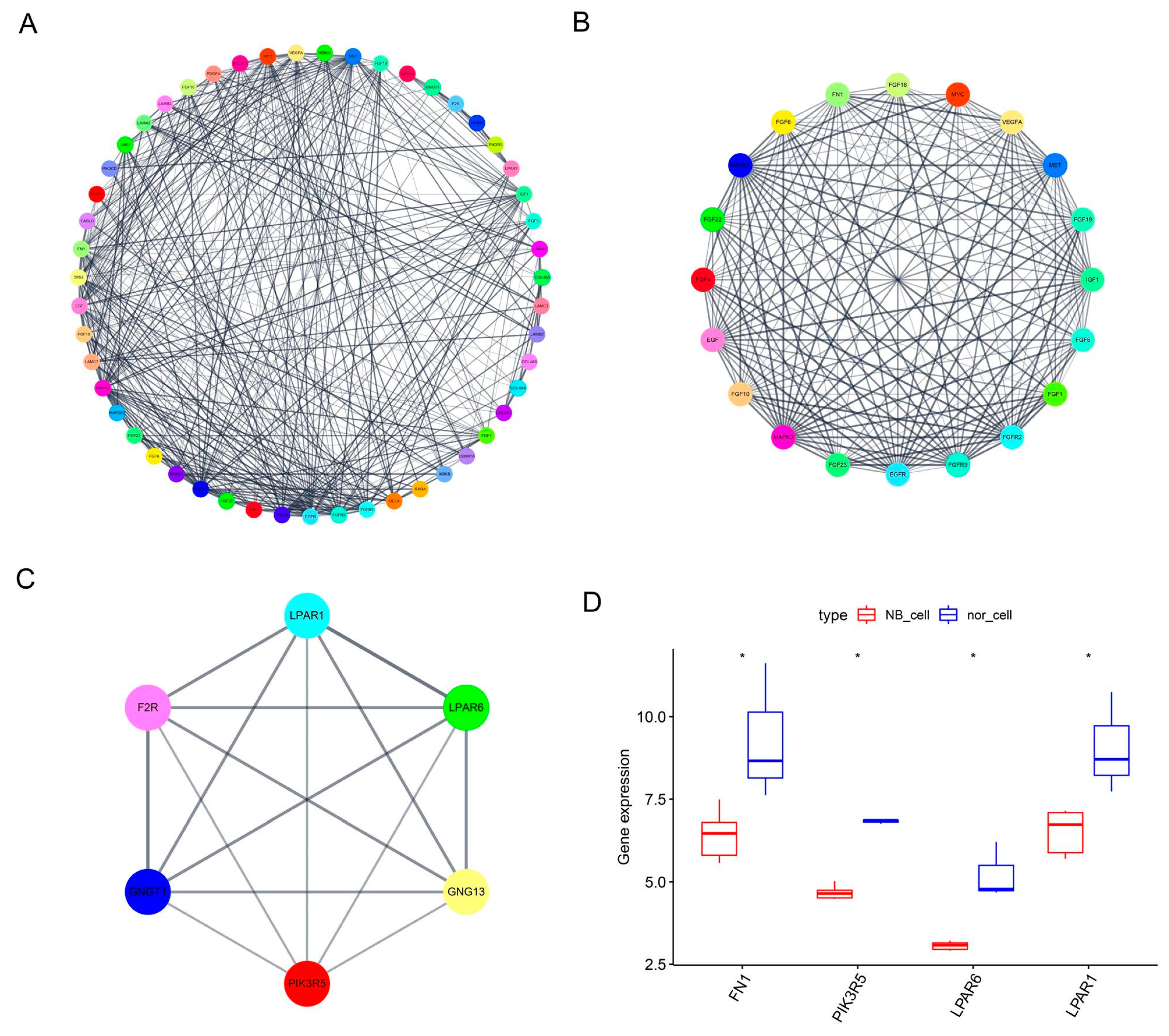
3.2. PPI Network Construction, Module Analysis and Hub Gene Determination
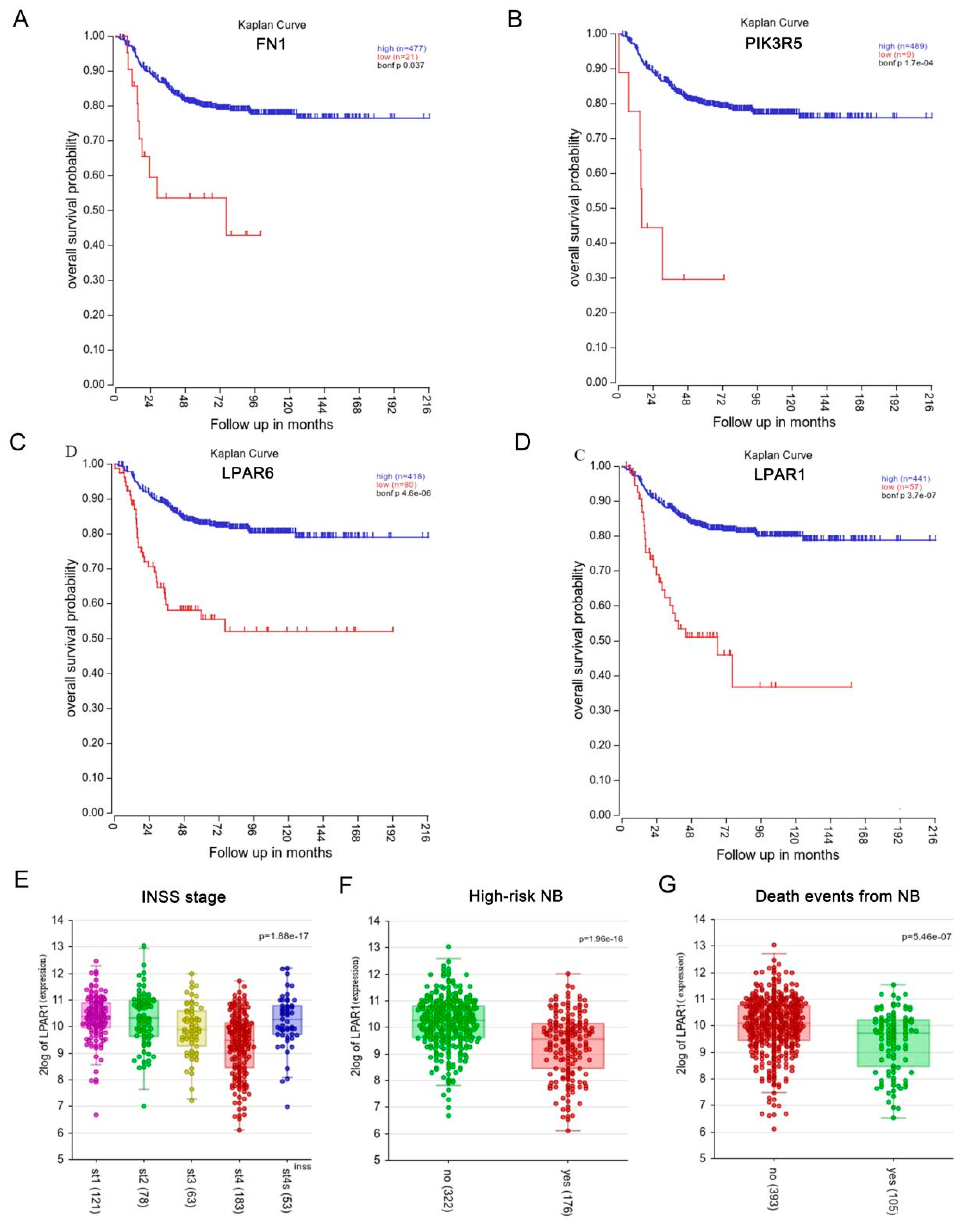
3.3. Hub Gene Expression and Survival Analysis
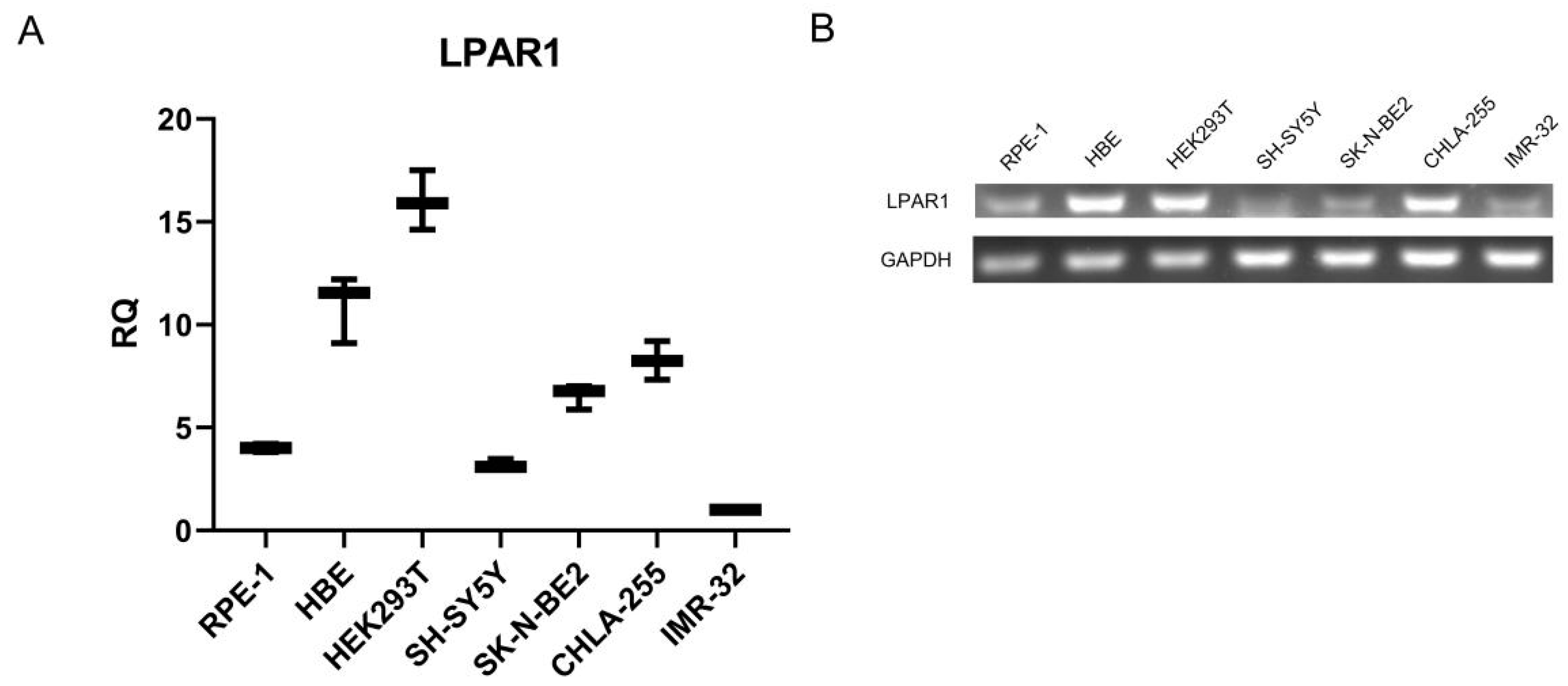
3.4. NB Cells Showed Low Expression Level of LPAR1 Compared to Non-Malignant Cell Lines
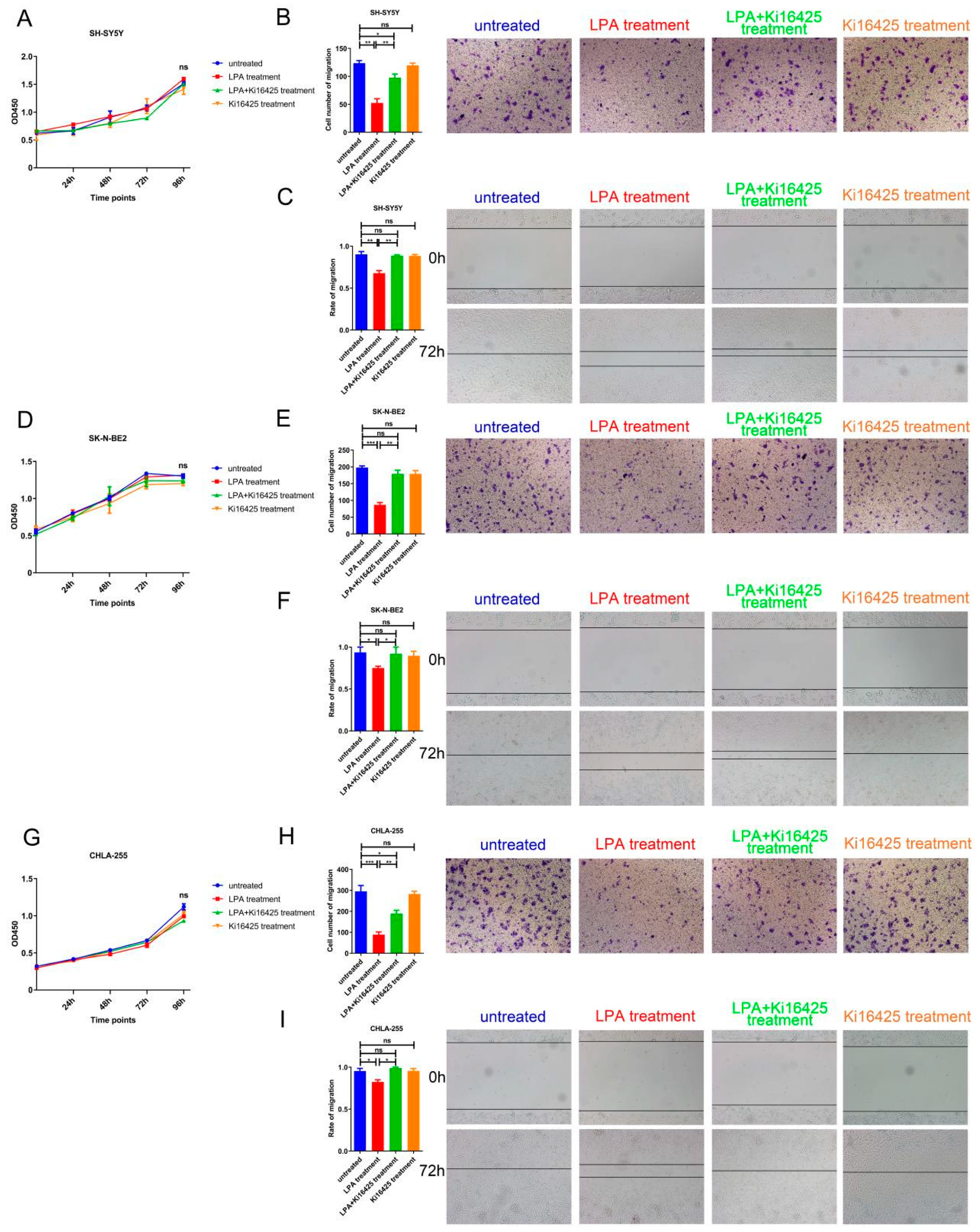
3.5. LPA Suppressed the Migration of NB Cells via LPAR1
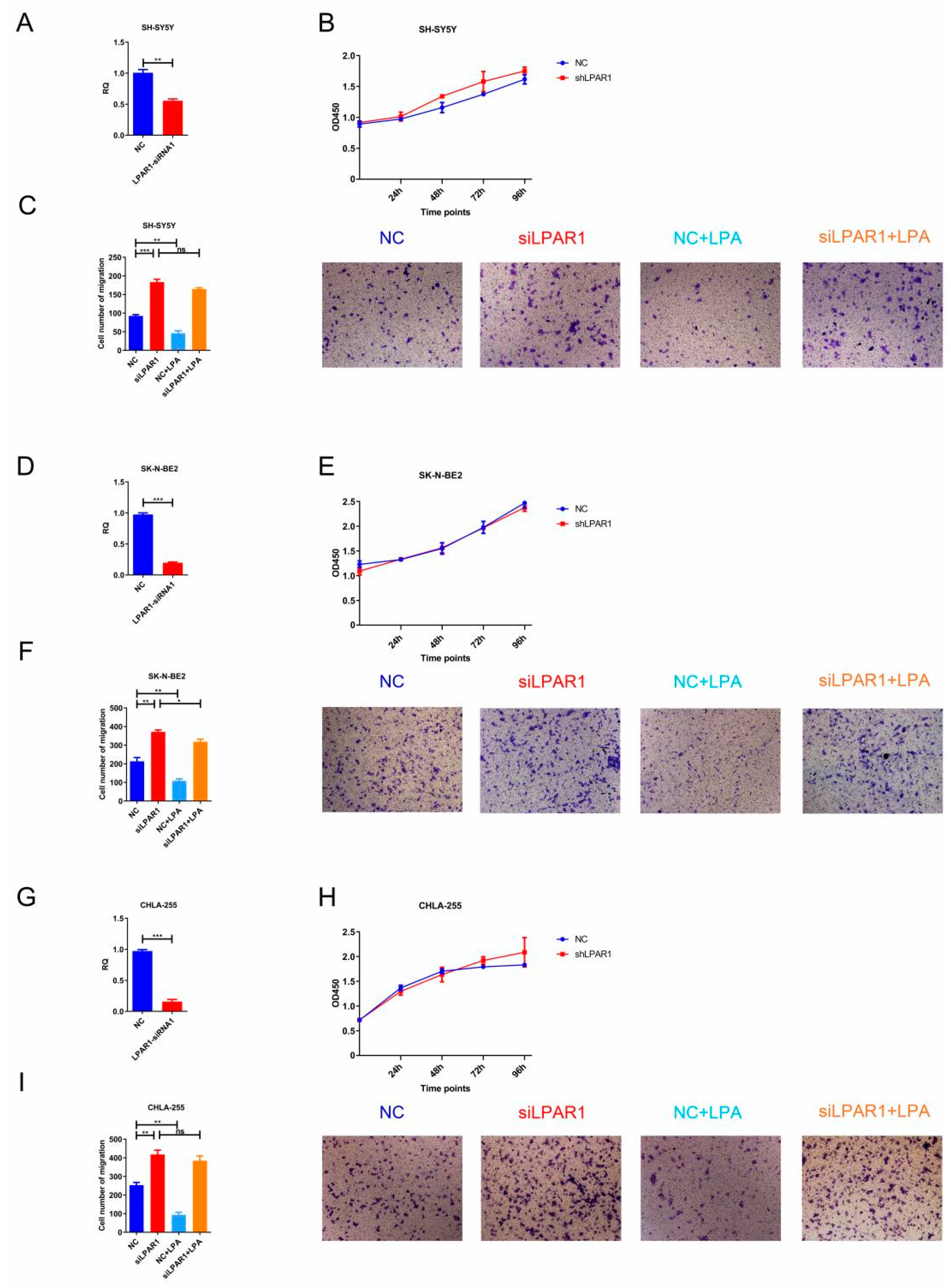
3.6. Knockdown of LPAR1 Promoted the Migration of NB Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Newman, E.A.; Abdessalam, S.; Aldrink, J.H.; Austin, M.; Heaton, T.E.; Bruny, J.; Ehrlich, P.; Dasgupta, R.; Baertschiger, R.M.; Lautz, T.B.; et al. Update on neuroblastoma. J. Pediatr. Surg. 2019, 54, 383–389. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, K.I.; Treis, D.; Johnsen, J.I. Neuroblastoma Heterogeneity, Plasticity, and Emerging Therapies. Curr. Oncol. Rep. 2022. [Google Scholar] [CrossRef]
- Toro-Dominguez, D.; Martorell-Marugan, J.; Lopez-Dominguez, R.; Garcia-Moreno, A.; Gonzalez-Rumayor, V.; Alarcon-Riquelme, M.E.; Carmona-Saez, P. ImaGEO: Integrative gene expression meta-analysis from GEO database. Bioinformatics 2019, 35, 880–882. [Google Scholar] [CrossRef] [PubMed]
- Barrett, T.; Troup, D.B.; Wilhite, S.E.; Ledoux, P.; Rudnev, D.; Evangelista, C.; Kim, I.F.; Soboleva, A.; Tomashevsky, M.; Marshall, K.A.; et al. NCBI GEO: Archive for high-throughput functional genomic data. Nucleic Acids Res. 2009, 37, D885–D890. [Google Scholar] [CrossRef]
- Lin, Y.H.; Lin, Y.C.; Chen, C.C. Lysophosphatidic Acid Receptor Antagonists and Cancer: The Current Trends, Clinical Implications, and Trials. Cells 2021, 10, 1629. [Google Scholar] [CrossRef] [PubMed]
- Harma, V.; Knuuttila, M.; Virtanen, J.; Mirtti, T.; Kohonen, P.; Kovanen, P.; Happonen, A.; Kaewphan, S.; Ahonen, I.; Kallioniemi, O.; et al. Lysophosphatidic acid and sphingosine-1-phosphate promote morphogenesis and block invasion of prostate cancer cells in three-dimensional organotypic models. Oncogene 2012, 31, 2075–2089. [Google Scholar] [CrossRef] [Green Version]
- Geraldo, L.; Spohr, T.; Amaral, R.; Fonseca, A.; Garcia, C.; Mendes, F.A.; Freitas, C.; Dossantos, M.F.; Lima, F. Role of lysophosphatidic acid and its receptors in health and disease: Novel therapeutic strategies. Signal Transduct. Target Ther. 2021, 6, 45. [Google Scholar] [CrossRef]
- Taniguchi, R.; Inoue, A.; Sayama, M.; Uwamizu, A.; Yamashita, K.; Hirata, K.; Yoshida, M.; Tanaka, Y.; Kato, H.E.; Nakada-Nakura, Y.; et al. Structural insights into ligand recognition by the lysophosphatidic acid receptor LPA6. Nature 2017, 548, 356–360. [Google Scholar] [CrossRef]
- Choi, J.W.; Herr, D.R.; Noguchi, K.; Yung, Y.C.; Lee, C.W.; Mutoh, T.; Lin, M.E.; Teo, S.T.; Park, K.E.; Mosley, A.N.; et al. LPA receptors: Subtypes and biological actions. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 157–186. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Jiang, D.; Yang, S.; Zhang, X.; Wang, J.; Liu, Y.; Sun, Y.; Lu, Y.; Yang, K. LPAR1, Correlated With Immune Infiltrates, Is a Potential Prognostic Biomarker in Prostate Cancer. Front. Oncol. 2020, 10, 846. [Google Scholar] [CrossRef]
- Ma, X.; Feng, J.; Lu, M.; Tang, W.; Han, J.; Luo, X.; Zhao, Q.; Yang, L. Microrna-501-5p promotes cell proliferation and migration in gastric cancer by downregulating LPAR1. J. Cell. Biochem. 2020, 121, 1911–1922. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Yoshikawa, K.; Tanabe, E.; Kitayoshi, M.; Fukui, R.; Fukushima, N.; Tsujiuchi, T. Opposite roles of LPA1 and LPA3 on cell motile and invasive activities of pancreatic cancer cells. Tumour. Biol. 2012, 33, 1739–1744. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, M.; Okabe, K.; Kato, K.; Okumura, M.; Fukui, R.; Fukushima, N.; Tsujiuchi, T. Differential function of lysophosphatidic acid receptors in cell proliferation and migration of neuroblastoma cells. Cancer Lett. 2012, 316, 91–96. [Google Scholar] [CrossRef]
- Hirane, M.; Araki, M.; Dong, Y.; Honoki, K.; Fukushima, N.; Tsujiuchi, T. Inhibitory effects of LPA1 on cell motile activities stimulated by hydrogen peroxide and 2,3-dimethoxy-1,4-naphthoquinone in fibroblast 3T3 cells. Biochem. Biophys. Res. Commun. 2013, 441, 47–52. [Google Scholar] [CrossRef]
- Irizarry, R.A.; Hobbs, B.; Collin, F.; Beazer-Barclay, Y.D.; Antonellis, K.J.; Scherf, U.; Speed, T.P. Exploration, normalization, and summaries of high density oligonucleotide array probe level data. Biostatistics 2003, 4, 249–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gautier, L.; Cope, L.; Bolstad, B.M.; Irizarry, R.A. Affy—analysis of Affymetrix GeneChip data at the probe level. Bioinformatics 2004, 20, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Ding, R.; Qu, Y.; Wu, C.H.; Vijay-Shanker, K. Automatic gene annotation using GO terms from cellular component domain. BMC Med. Inform. Decis. Mak. 2018, 18 (Suppl. S5), 119. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Dennis, G.J.; Sherman, B.T.; Hosack, D.A.; Yang, J.; Gao, W.; Lane, H.C.; Lempicki, R.A. DAVID: Database for Annotation, Visualization, and Integrated Discovery. Genome Biol. 2003, 4, 3. [Google Scholar] [CrossRef]
- Feng, H.; Gu, Z.Y.; Li, Q.; Liu, Q.H.; Yang, X.Y.; Zhang, J.J. Identification of significant genes with poor prognosis in ovarian cancer via bioinformatical analysis. J. Ovarian Res. 2019, 12, 35. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.; Kitagawa, T.; Baron, B.; Abiko, Y.; Chiba, I.; Kuramitsu, Y. ITGA2, LAMB3, and LAMC2 may be the potential therapeutic targets in pancreatic ductal adenocarcinoma: An integrated bioinformatics analysis. Sci. Rep. 2021, 11, 10563. [Google Scholar] [CrossRef]
- Zhang, H.; Chai, W.; Yang, W.; Han, W.; Mou, W.; Xi, Y.; Chen, X.; Wang, H.; Wang, W.; Qin, H.; et al. The increased IL-17-producing gammadeltaT cells promote tumor cell proliferation and migration in neuroblastoma. Clin. Immunol. 2020, 211, 108343. [Google Scholar] [CrossRef] [PubMed]
- Noorolyai, S.; Shajari, N.; Baghbani, E.; Sadreddini, S.; Baradaran, B. The relation between PI3K/AKT signalling pathway and cancer. Gene 2019, 698, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Rich, J.T.; Neely, J.G.; Paniello, R.C.; Voelker, C.C.; Nussenbaum, B.; Wang, E.W. A practical guide to understanding Kaplan-Meier curves. Otolaryngol. Head Neck. Surg. 2010, 143, 331–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Li, M.; Yin, N.; Zhang, J. The Expression Regulation and Biological Function of Autotaxin. Cells 2021, 10, 939. [Google Scholar] [CrossRef]
- Zuckerman, V.; Sokolov, E.; Swet, J.H.; Ahrens, W.A.; Showlater, V.; Iannitti, D.A.; Mckillop, I.H. Expression and function of lysophosphatidic acid receptors (LPARs) 1 and 3 in human hepatic cancer progenitor cells. Oncotarget 2016, 7, 2951–2967. [Google Scholar] [CrossRef] [Green Version]
- Takagi, S.; Sasaki, Y.; Koike, S.; Takemoto, A.; Seto, Y.; Haraguchi, M.; Ukaji, T.; Kawaguchi, T.; Sugawara, M.; Saito, M.; et al. Platelet-derived lysophosphatidic acid mediated LPAR1 activation as a therapeutic target for osteosarcoma metastasis. Oncogene 2021, 40, 5548–5558. [Google Scholar] [CrossRef]
- Ha, J.H.; Ward, J.D.; Radhakrishnan, R.; Jayaraman, M.; Song, Y.S.; Dhanasekaran, D.N. Lysophosphatidic acid stimulates epithelial to mesenchymal transition marker Slug/Snail2 in ovarian cancer cells via Galphai2, Src, and HIF1alpha signaling nexus. Oncotarget 2016, 7, 37664–37679. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Rebecca, V.W.; Kossenkov, A.V.; Connelly, T.; Liu, Q.; Gutierrez, A.; Xiao, M.; Li, L.; Zhang, G.; Samarkina, A.; et al. Neural Crest-Like Stem Cell Transcriptome Analysis Identifies LPAR1 in Melanoma Progression and Therapy Resistance. Cancer Res. 2021, 81, 5230–5241. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.S.; Johansson, P.; Chen, L.; Song, Y.K.; Tolman, C.; Li, S.; Hurd, L.; Patidar, R.; Wen, X.; Badgett, T.C.; et al. Massively parallel sequencing reveals an accumulation of de novo mutations and an activating mutation of LPAR1 in a patient with metastatic neuroblastoma. PLoS ONE 2013, 8, e77731. [Google Scholar] [CrossRef] [PubMed]
- Ishii, S.; Tsujiuchi, T.; Fukushima, N. Functional characterization of lysophosphatidic acid receptor 1 mutants identified in rat cancer tissues. Biochem. Biophys. Res. Commun. 2017, 486, 767–773. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Fukui, R.; Okabe, K.; Tanabe, E.; Kitayoshi, M.; Fukushima, N.; Tsujiuchi, T. Constitutively active lysophosphatidic acid receptor-1 enhances the induction of matrix metalloproteinase-2. Biochem. Biophys. Res. Commun. 2012, 417, 790–793. [Google Scholar] [CrossRef] [PubMed]
- Kitayoshi, M.; Kato, K.; Tanabe, E.; Yoshikawa, K.; Fukui, R.; Fukushima, N.; Tsujiuchi, T. Enhancement of endothelial cell migration by constitutively active LPA(1)-expressing tumor cells. Biochem. Biophys. Res. Commun. 2012, 422, 339–343. [Google Scholar] [CrossRef]
- Tan, X.; Gong, W.; Chen, B.; Gong, B.; Hua, Z.; Zhang, S.; Chen, Y.; Li, Q.; Li, Z. Downregulation of fibronectin 1 attenuates ATRA-induced inhibition of cell migration and invasion in neuroblastoma cells. Mol. Cell. Biochem. 2021, 476, 3601–3612. [Google Scholar] [CrossRef]
- Liu, W.; Jiang, D.; Gong, F.; Huang, Y.; Luo, Y.; Rong, Y.; Wang, J.; Ge, X.; Ji, C.; Fan, J.; et al. MiR-210-5p promotes epithelial-mesenchymal transition by inhibiting PIK3R5 thereby activating oncogenic autophagy in osteosarcoma cells. Cell Death Dis. 2020, 11, 93. [Google Scholar] [CrossRef] [Green Version]
- Boeva, V.; Louis-Brennetot, C.; Peltier, A.; Durand, S.; Pierre-Eugene, C.; Raynal, V.; Etchevers, H.C.; Thomas, S.; Lermine, A.; Daudigeos-Dubus, E.; et al. Heterogeneity of neuroblastoma cell identity defined by transcriptional circuitries. Nat. Genet. 2017, 49, 1408–1413. [Google Scholar] [CrossRef]
- van Groningen, T.; Koster, J.; Valentijn, L.J.; Zwijnenburg, D.A.; Akogul, N.; Hasselt, N.E.; Broekmans, M.; Haneveld, F.; Nowakowska, N.E.; Bras, J.; et al. Neuroblastoma is composed of two super-enhancer-associated differentiation states. Nat. Genet. 2017, 49, 1261–1266. [Google Scholar] [CrossRef]
- Gartlgruber, M.; Sharma, A.K.; Quintero, A.; Dreidax, D.; Jansky, S.; Park, Y.G.; Kreth, S.; Meder, J.; Doncevic, D.; Saary, P.; et al. Super enhancers define regulatory subtypes and cell identity in neuroblastoma. Nat. Cancer 2021, 2, 114–128. [Google Scholar] [CrossRef]
- Lazic, D.; Kromp, F.; Rifatbegovic, F.; Repiscak, P.; Kirr, M.; Mivalt, F.; Halbritter, F.; Bernkopf, M.; Bileck, A.; Ussowicz, M.; et al. Landscape of Bone Marrow Metastasis in Human Neuroblastoma Unraveled by Transcriptomics and Deep Multiplex Imaging. Cancers 2021, 13, 4311. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | GO ID | Term | Gene ID | p-Value |
|---|---|---|---|---|
| BP | GO:0030198 | Extracellular matrix organization | VIT, ITGB4, TNC, F11R, TNF, DAG1 | 1.08 × 10−8 |
| BP | GO:0001525 | Angiogenesis | CIB1, CTGF, LEPR, SYK, EREG, TGFA | 1.22 × 10−7 |
| BP | GO:0007155 | Cell adhesion | TNC, COMP, TNR, FEZ1, CD151, LPP | 2.70 × 10−6 |
| BP | GO:0043123 | Positive regulation of NF-κB signaling | TNF, CCR7, LTF, IRF3, LPAR1, NEK6 | 1.89 × 10−5 |
| BP | GO:0042127 | Regulation of cell proliferation | TES, CXCL3, JAK1, FGR, ACE2, LCK | 1.06 × 10−5 |
| BP | GO:0014066 | Regulation of PI3K signaling | KLB, EGFR, IER3, BTC, NRG4, IRS1 | 5.35 × 10−5 |
| BP | GO:0030335 | Positive regulation of cell migration | ILK, PLAU, FGR, LEF1, CCL7, DAB2 | 3.02 × 10−4 |
| BP | GO:0000187 | Activation of MAPK activity | LPAR1, PLCE1, GRM4, WNT5A, MOS | 0.003828 |
| CC | GO:0005886 | Plasma membrane | SLA2, LIPH, AR, ACE2, FPR3, MYO6 | 2.53 × 10−25 |
| CC | GO:0009986 | Cell surface | LIPG, KRT4, BST2, TF, CALR, SHH | 1.58 × 10−4 |
| CC | GO:0005578 | Proteinaceous extracellular matrix | GLDN, TNR, LOX, PI3, CILP, CALR | 1.66 × 10−10 |
| CC | GO:0031090 | Organelle membrane | FAAH, TFPI, FMO1, FA2H, CYP2S1 | 3.28 × 10−7 |
| CC | GO:0005911 | Cell-cell junction | MLC1, KRT8, DSG2, TLN1, VCL | 9.37 × 10−5 |
| CC | GO:0005925 | Focal adhesion | TNC, PVR, TNS4, EZR, PXN, CALR | 0.002148 |
| CC | GO:0005789 | Endoplasmic reticulum membrane | ALG1, POR, HPD, RCE1, PIGS, PIGZ | 0.008747 |
| MF | GO:0005509 | Calcium ion binding | SYTL2, REG4, AIF1L, EHD1, CALR | 2.32 × 10−6 |
| MF | GO:0004872 | Receptor activity | PVR, THBD, TLR1, LRP1, CALCR | 1.06 × 10−6 |
| MF | GO:0046934 | PIK3Ca activity | KLB, PIK3R5, EGF, BTC, LCK, NRG4 | 7.22 × 10−5 |
| MF | GO:0004896 | Cytokine receptor activity | FLT3, MPL, CSF2RB, OSMR, CD44 | 0.001221 |
| KEGG ID | Term | Gene ID | p-Value |
|---|---|---|---|
| hsa04060 | Cytokine-cytokine receptor interaction | MPL, EDAR, NGFR, LIF, EDA, PRL | 2.59 × 10−18 |
| hsa04514 | Cell adhesion molecules (CAMs) | PVR, SPN, CTLA4, CD8A, SELP | 1.01 × 10−5 |
| hsa04630 | Jak-STAT signaling pathway | OXTR, LEPR, LPAR1, MC2R, PLG | 4.98 × 10−5 |
| hsa04668 | TNF signaling pathway | RELA, JUN, EDN1, JAG1, MLKL | 2.38 × 10−6 |
| hsa04064 | NF-kappa B signaling pathway | PTGS2, RELA, PLAU, SYK, LTBR | 0.002268 |
| hsa04510 | Focal adhesion | MYLK, TNR, VWF, VCL, SRC, SPP1 | 0.002427 |
| hsa04151 | PI3K-Akt signaling pathway | TP53, LPAR1, CHAD, PCK1, PRL | 0.005575 |
| hsa05200 | Pathways in cancer | MITF, TP53, LPAR1, LPAR6, FLT3 | 0.011031 |
| hsa04010 | MAPK signaling pathway | FOS, TP53, RRAS, FLNB, NTRK2 | 0.037591 |
| hsa04020 | Calcium signaling pathway | RYR1, OXTR, PLCE1, ORAI1, ITPR3 | 0.040378 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Pei, M.; Yu, Y.; Wang, X.; Gui, J. Reduction of LPAR1 Expression in Neuroblastoma Promotes Tumor Cell Migration. Cancers 2022, 14, 3346. https://doi.org/10.3390/cancers14143346
Liu X, Pei M, Yu Y, Wang X, Gui J. Reduction of LPAR1 Expression in Neuroblastoma Promotes Tumor Cell Migration. Cancers. 2022; 14(14):3346. https://doi.org/10.3390/cancers14143346
Chicago/Turabian StyleLiu, Xiangjun, Mengmiao Pei, Yongbo Yu, Xiaolin Wang, and Jingang Gui. 2022. "Reduction of LPAR1 Expression in Neuroblastoma Promotes Tumor Cell Migration" Cancers 14, no. 14: 3346. https://doi.org/10.3390/cancers14143346