
Brain Functional Connectivity in Low- and High-Grade Gliomas: Differences in Network Dynamics Associated with Tumor Grade and Location

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
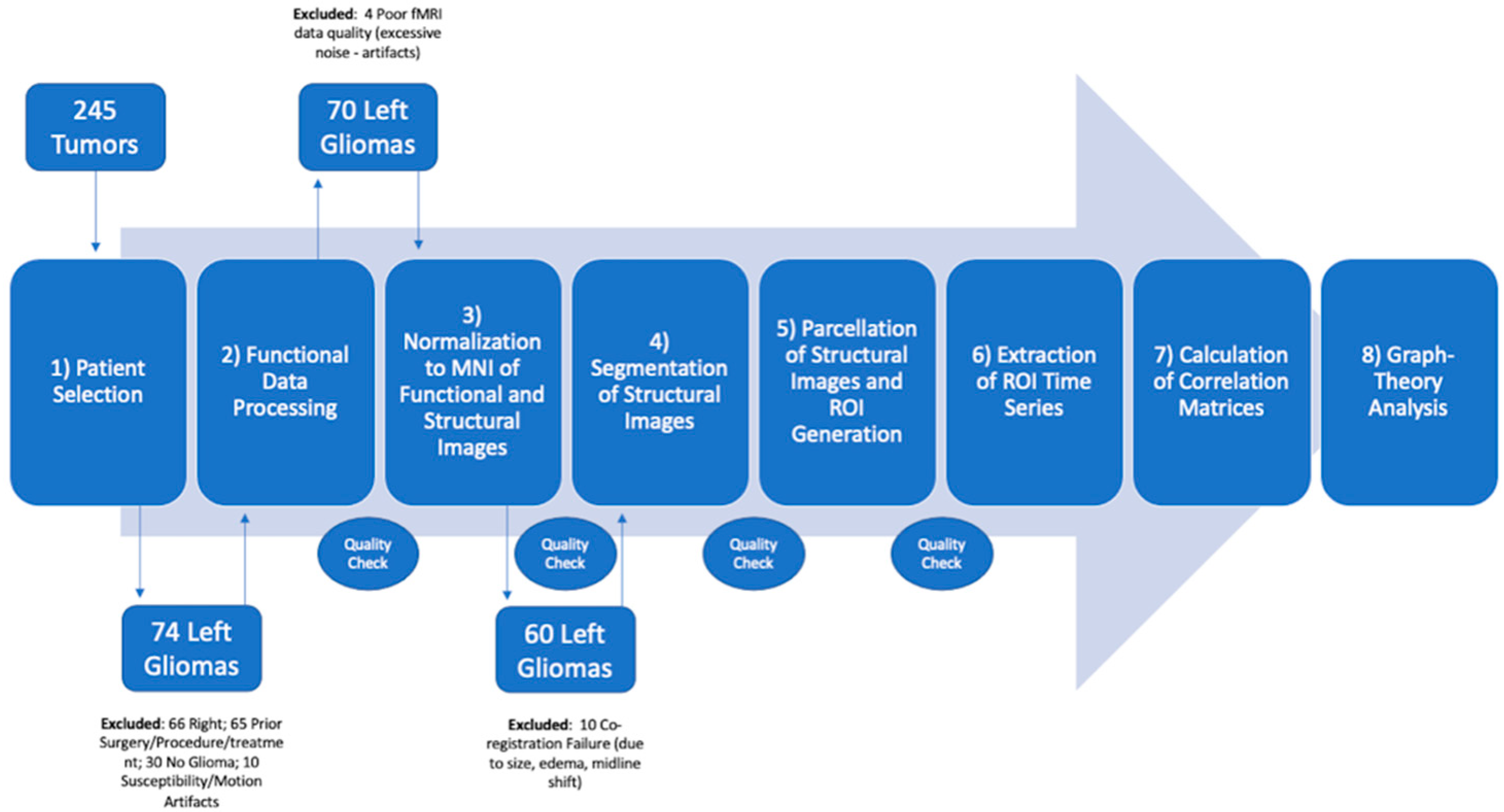
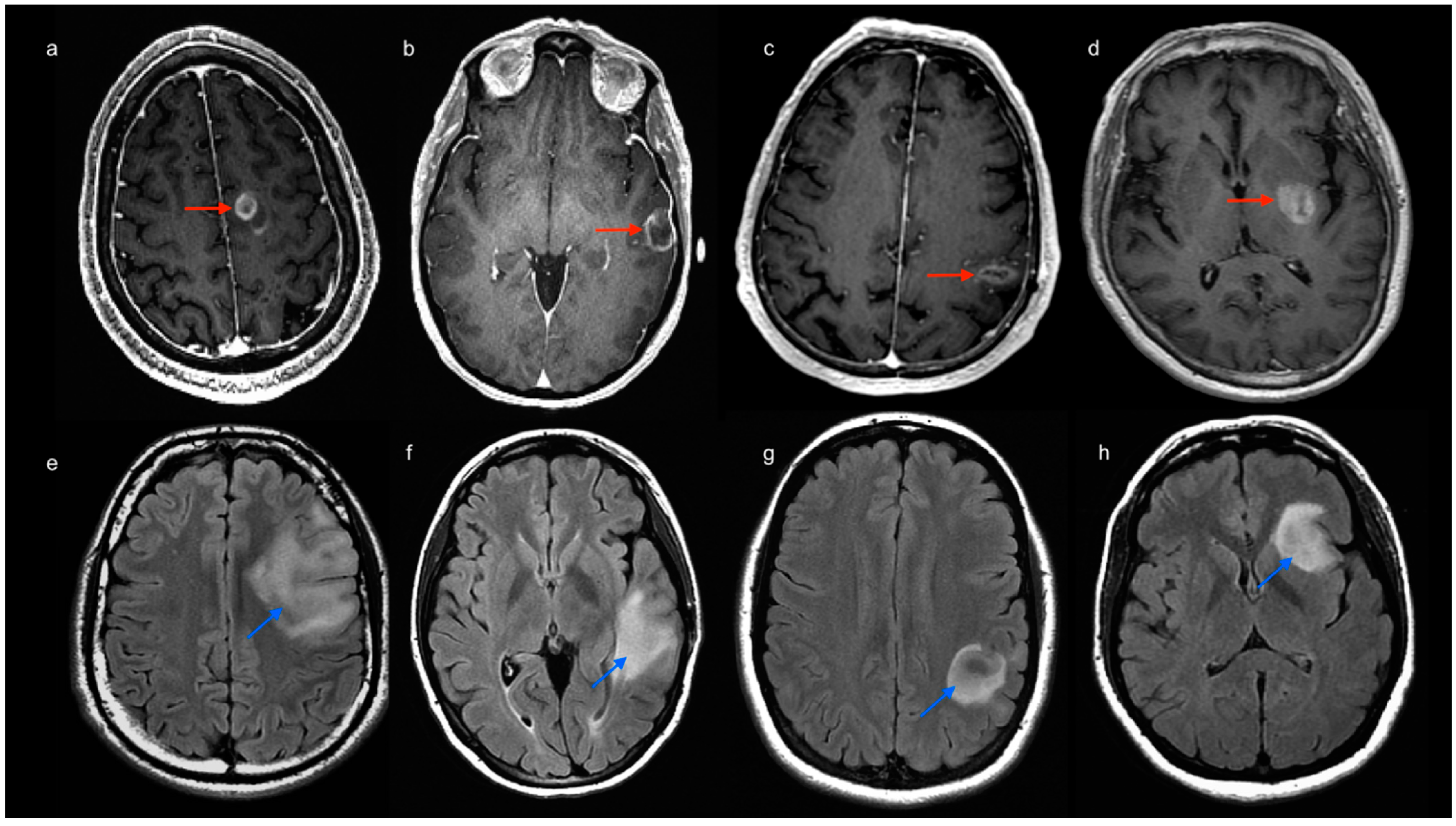
2.1. Patients
2.2. MRI Acquisition
2.3. Functional Connectivity Analysis
2.4. Statistical Analyses
3. Results
3.1. Patients
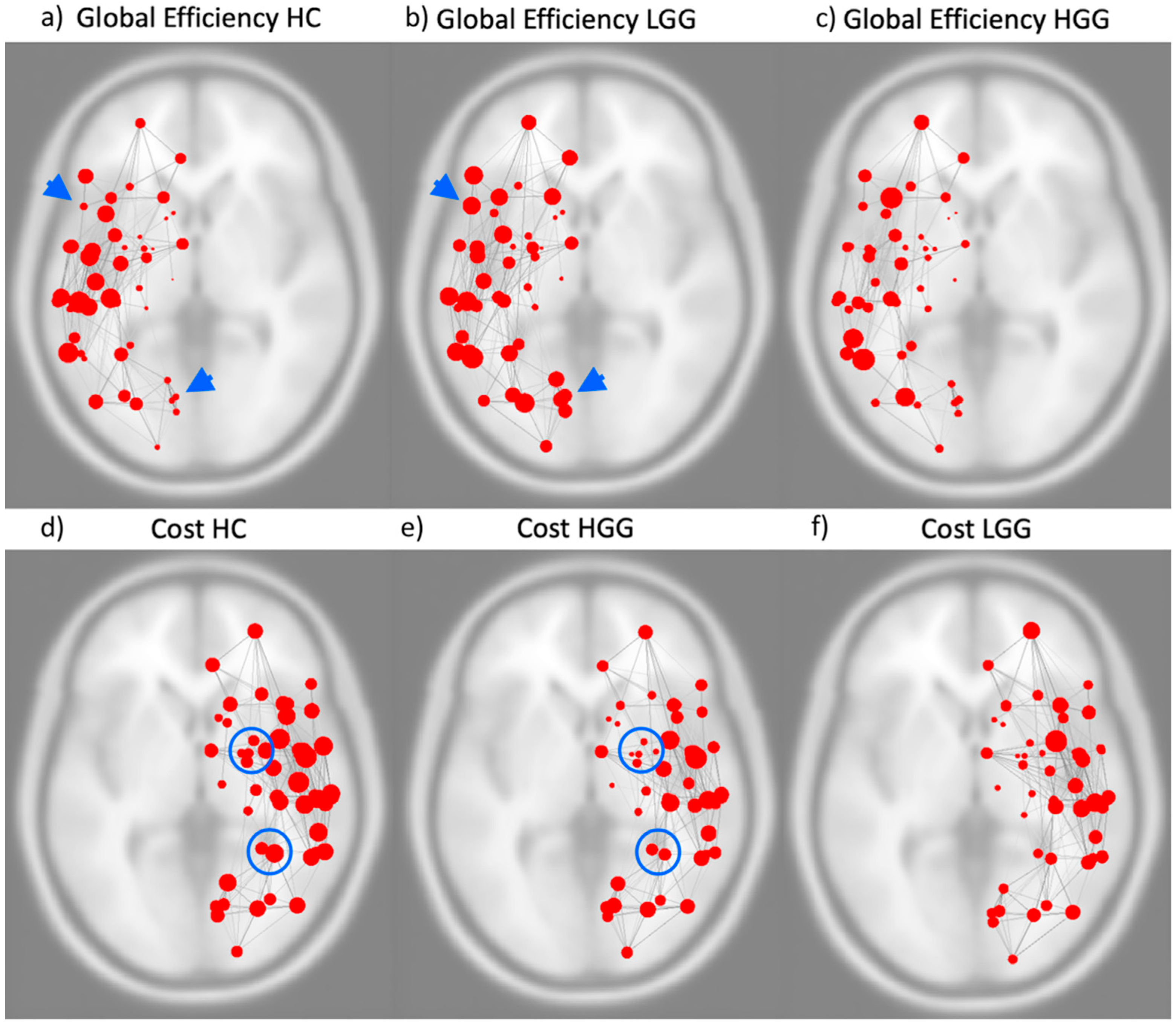
3.2. Graph Theory Analysis: Whole-Brain and Hemispheric Network Analysis
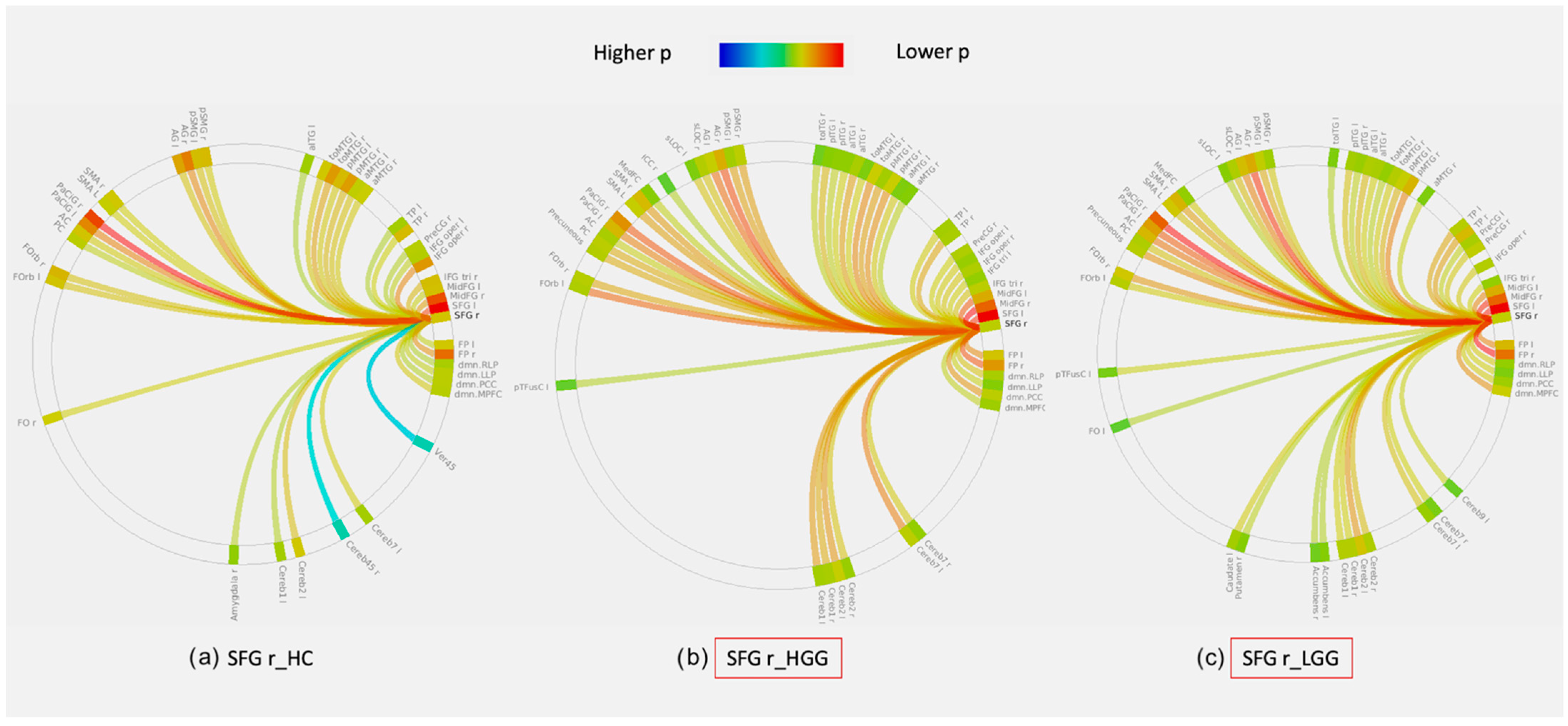
3.3. Graph Theory Analysis: Lobar Networks Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pasquini, L.; Di Napoli, A.; Rossi-Espagnet, M.C.; Visconti, E.; Napolitano, A.; Romano, A.; Bozzao, A.; Peck, K.K.; Holodny, A.I. Understanding language reorganization with neuroimaging: How language adapts to different focal lesions. Insights into clinical applications. Front. Hum. Neurosci. 2022, 16, 747215. [Google Scholar] [CrossRef] [PubMed]
- Cargnelutti, E.; Ius, T.; Skrap, M.; Tomasino, B. What do we know about pre- and postoperative plasticity in patients with glioma? A review of neuroimaging and intraoperative mapping studies. NeuroImage Clin. 2020, 28, 102435. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Rivera, P.A.; Rios-Lago, M.; Sanchez-Casarrubios, S.; Salazar, O.; Yus, M.; González-Hidalgo, M.; Sanz, A.; Avecillas-Chasin, J.; Alvarez-Linera, J.; Pascual-Leone, A.; et al. Cortical plasticity catalyzed by prehabilitation enables extensive resection of brain tumors in eloquent areas. J. Neurosurg. 2017, 126, 1323–1333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karbe, H.; Thiel, A.; Weber-luxenburger, G.; Kessler, J.; Heiss, W. Brain plasticity in poststroke aphasia: What is the contribution of the right hemisphere? pet studies of aphasic stroke patients showed a significant correlation between the neuropsychological deficit and the metabolic impairment of speech-relevant area. Brain Lang. 1998, 230, 215–230. [Google Scholar] [CrossRef]
- Nahmani, M.; Turrigiano, G.G. Adult cortical plasticity following injury: Recapitulation of critical period mechanisms? Neurosci. 2014, 283, 4–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desmurget, M.; Bonnetblanc, F.; Duffau, H. Contrasting acute and slow-growing lesions: A new door to brain plasticity. Brain 2007, 130, 898–914. [Google Scholar] [CrossRef] [Green Version]
- Deverdun, J.; Van Dokkum, L.E.H.; Le Bars, E.; Herbet, G.; Mura, T.; D’agata, B.; Picot, M.C.; Menjot, N.; Molino, F.; Duffau, H.; et al. Language reorganization after resection of low-grade gliomas: An fMRI task based connectivity study. Brain Imaging Behav. 2020, 14, 1779–1791. [Google Scholar] [CrossRef]
- Gratton, C.; Nomura, E.M.; Pérez, F.; D’Esposito, M. Focal brain lesions to critical locations cause widespread disruption of the modular organization of the brain. J. Cogn. Neurosci. 2012, 24, 1275–1285. [Google Scholar] [CrossRef] [Green Version]
- Bullmore, E.; Sporns, O. Complex brain networks: Graph theoretical analysis of structural and functional systems. Nat. Rev. Neurosci. 2009, 10, 186–198. [Google Scholar] [CrossRef]
- Bottino, F.; Lucignani, M.; Pasquini, L.; Mastrogiovanni, M.; Gazzellini, S.; Ritrovato, M.; Longo, D.; Figà-Talamanca, L.; Rossi Espagnet, M.C.; Napolitano, A. Spatial stability of functional networks: A measure to assess the robustness of graph-theoretical metrics to spatial errors related to brain parcellation. Front. Neurosci. 2022, 15, 736524. [Google Scholar] [CrossRef]
- Li, Q.; Pasquini, L.; del Ferraro, G.; Gene, M.; Peck, K.K.; Makse, H.A.; Holodny, A.I. Monolingual and bilingual language networks in healthy subjects using functional MRI and graph theory. Sci. Rep. 2021, 11, 10568. [Google Scholar] [CrossRef] [PubMed]
- Derks, J.; Dirkson, A.R.; de Witt Hamer, P.C.; van Geest, Q.; Hulst, H.E.; Barkhof, F.; Pouwels, P.J.W.; Geurts, J.J.G.; Reijneveld, J.C.; Douw, L. Connectomic profile and clinical phenotype in newly diagnosed glioma patients. NeuroImage Clin. 2017, 14, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Hart, M.G.; Romero-Garcia, R.; Price, S.J.; Suckling, J. Global effects of focal brain tumors on functional complexity and network robustness: A prospective cohort study. Clin. Neurosurg. 2019, 84, 1201–1213. [Google Scholar] [CrossRef] [Green Version]
- Latora, V.; Marchiori, M. Efficient behavior of small-world networks. Phys. Rev. Lett. 2001, 87, 198701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louis, D.N.; Perry, A.; Reifenberger, G.; Von Deimling, A.; Figarella-Branger, D.; Cavenee, W.K.; Ohgaki, H.; Wiestler, O.D.; Kleihues, P.; Ellison, D.W. The 2016 World Health Organization classification of tumors of the central nervous system: A summary. Acta Neuropathol. 2016, 131, 803–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan, E.; Goodglass, H.; Weintraub, S. Boston Naming Test, 2nd ed.; Lea & Febiger: Philadelphia, PA, USA, 1983. [Google Scholar]
- Whitfield-Gabrieli, S.; Nieto-Castanon, A. Conn: A functional connectivity toolbox for correlated and anticorrelated brain networks. Brain Connect. 2012, 2, 125–141. [Google Scholar] [CrossRef] [Green Version]
- Bera, A.K.; Jarque, C.M. Efficient tests for normality, homoscedasticity and serial independence of regression residuals. Monte Carlo evidence. Econ. Lett. 1981, 7, 313–318. [Google Scholar] [CrossRef]
- Yang, D. Standardized MRI assessment of high-grade glioma response: A review of the essential elements and pitfalls of the RANO criteria. Neuro-Oncol. Pract. 2014, 3, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Kristo, G.; Raemaekers, M.; Rutten, G.J.; de Gelder, B.; Ramsey, N.F. Inter-hemispheric language functional reorganization in low-grade glioma patients after tumour surgery. Cortex 2015, 64, 235–248. [Google Scholar] [CrossRef]
- De Abreu, V.H.F.; Peck, K.K.; Petrovich-Brennan, N.M.; Woo, K.M.; Holodny, A.I. Brain tumors: The influence of tumor type and routine MR imaging characteristics at BOLD functional MR imaging in the primary motor gyrus. Radiology 2016, 281, 876–883. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.C.; Feldman, S.C.; Schulder, M.; Kalnin, A.J.; Holodny, A.I.; Zimmerman, A.; Sinensky, R.; Rao, S. The effect of tumour type and distance on activation in the motor cortex. Neuroradiology 2005, 47, 813–819. [Google Scholar] [CrossRef] [PubMed]
- Mallela, A.N.; Peck, K.K.; Petrovich-Brennan, N.M.; Zhang, Z.; Lou, W.; Holodny, A.I. Altered resting-state functional connectivity in the hand motor network in glioma patients. Brain Connect. 2016, 6, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Vollmann-Zwerenz, A.; Leidgens, V.; Feliciello, G.; Klein, C.A.; Hau, P. Tumor cell invasion in glioblastoma. Int. J. Mol. Sci. 2020, 21, 1932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrera, E.; Tononi, G. Diaschisis: Past, present, future. Brain 2014, 137, 2408–2422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duffau, H. Hodotopy, neuroplasticity and diffuse gliomas. Neurochirurgie 2017, 63, 259–265. [Google Scholar] [CrossRef]
- Zhang, N.; Xia, M.; Qiu, T.; Wang, X.; Lin, C.-P.; Guo, Q.; Lu, J.; Wu, Q.; Zhuang, D.; Yu, Z.; et al. Reorganization of cerebro-cerebellar circuit in patients with left hemispheric gliomas involving language network: A combined structural and resting-state functional mri study. Hum. Brain Mapp. 2018, 39, 4802–4819. [Google Scholar] [CrossRef]
- Holodny, A.I.; Schulder, M.; Ybasco, A.; Liu, W.C. Translocation of Broca’s area to the contralateral hemisphere as the result of the growth of a left inferior frontal glioma. J. Comput. Assist. Tomogr. 2002, 26, 941–943. [Google Scholar] [CrossRef]
- Petrovich, N.M.; Holodny, A.I.; Brennan, C.W.; Gutin, P.H. Isolated translocation of Wernicke’s area to the right hemisphere in a 62-year-man with a temporo-parietal glioma. Am. J. Neuroradiol. 2004, 25, 130–133. [Google Scholar]
- Gębska-Kośla, K.; Bryszewski, B.; Jaskólski, D.J.; Fortuniak, J.; Niewodniczy, M.; Stefańczyk, L.; Majos, A. Reorganization of language centers in patients with brain tumors located in eloquent speech areas–A pre-and postoperative preliminary fMRI study. Neurol. I Neurochir. Pol. 2017, 51, 403–410. [Google Scholar] [CrossRef]
- Stensjøen, A.L.; Berntsen, E.M.; Jakola, A.S.; Solheim, O. when did the glioblastoma start growing, and how much time can be gained from surgical resection? A model based on the pattern of glioblastoma growth in vivo. Clin. Neurol. Neurosurg. 2018, 170, 38–42. [Google Scholar] [CrossRef]
- Draganski, B.; Gaser, C.; Kempermann, G.; Kuhn, H.G.; Winkler, J.; Büchel, C.; May, A. Temporal and spatial dynamics of brain structure changes during extensive learning. J. Neurosci. 2006, 26, 6314–6317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; del Ferraro, G.; Pasquini, L.; Peck, K.K.; Makse, H.A.; Holodny, A.I. Core language brain network for fMRI language task used in clinical applications. Netw. Neurosci. 2020, 4, 134–154. [Google Scholar] [CrossRef] [PubMed]
- Friederici, A.D.; Gierhan, S.M.E. The language network. Curr. Opin. Neurobiol. 2013, 23, 250–254. [Google Scholar] [CrossRef] [PubMed]
- Nenning, K.H.; Furtner, J.; Kiesel, B.; Schwartz, E.; Roetzer, T.; Fortelny, N.; Bock, C.; Grisold, A.; Marko, M.; Leutmezer, F.; et al. Distributed changes of the functional connectome in patients with glioblastoma. Sci. Rep. 2020, 10, 18312. [Google Scholar] [CrossRef]
- Keidel, J.L.; Welbourne, S.R.; Lambon Ralph, M.A. Solving the paradox of the equipotential and modular brain: A neurocomputational model of stroke vs. slow-growing glioma. Neuropsychologia 2010, 48, 1716–1724. [Google Scholar] [CrossRef]
- Briganti, C.; Sestieri, C.; Mattei, P.A.; Esposito, R.; Galzio, R.J.; Tartaro, A.; Romani, G.L.; Caulo, M. Reorganization of functional connectivity of the language network in patients with brain gliomas. Am. J. Neuroradiol. 2012, 33, 1983–1990. [Google Scholar] [CrossRef] [Green Version]
- Hadjiabadi, D.H.; Pung, L.; Zhang, J.; Ward, B.D.; Lim, W.T.; Kalavar, M.; Thakor, N.V.; Biswal, B.B.; Pathak, A.P. Brain tumors disrupt the resting-state connectome. NeuroImage Clin. 2018, 18, 279–289. [Google Scholar] [CrossRef]
- Kardan, O.; Reuter-Lorenz, P.A.; Peltier, S.; Churchill, N.W.; Misic, B.; Askren, M.K.; Jung, M.S.; Cimprich, B.; Berman, M.G. Brain connectivity tracks effects of chemotherapy separately from behavioral measures. NeuroImage Clin. 2019, 21, 101654. [Google Scholar] [CrossRef]
- Mitchell, T.J.; Seitzman, B.A.; Ballard, N.; Petersen, S.E.; Shimony, J.S.; Leuthardt, E.C. Human brain functional network organization is disrupted after whole-brain radiation therapy. Brain Connect. 2020, 10, 29–38. [Google Scholar] [CrossRef]
- Bahrami, N.; Seibert, T.M.; Karunamuni, R.; Bartsch, H.; Krishnan, A.P.; Farid, N.; Hattangadi-Gluth, J.A.; McDonald, C.R. altered network topology in patients with primary brain tumors after fractionated radiotherapy. Brain Connect. 2017, 7, 299–308. [Google Scholar] [CrossRef]
- Burke, S.N.; Barnes, C.A. Neural plasticity in the ageing brain. Nat. Rev. Neurosci. 2006, 7, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Liang, X.; Ou, J.; Li, H.; Luo, Y.J.; Tan, L.H. Sex differences in functional brain networks for language. Cereb. Cortex 2020, 30, 1528–1537. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Frontal Tumors | HGG/Controls | p-Value | LGG/Controls | p-Value | HGG/LGG | p-Value |
|---|---|---|---|---|---|---|
| Global Efficiency | SFG r | 0.006 | ||||
| Local Efficiency | SFG r | 0.045 | SFG r | 0.045 | ||
| Betweenness Centrality | IFG tri l | 0.006 | SFG r | 0.025 | IFG tri l | 0.001 |
| SFG l | 0.012 | |||||
| Cost | SFG r | 0.028 | ||||
| Average Path Length | SFG r | 0.003 | SFG r | 0.045 | ||
| SFG l | 0.043 | |||||
| Clustering Coefficient | SFG r | 0.017 | MidFG l | 0.042 | ||
| IFG tri l | 0.016 | |||||
| Degree | SFG r | 0.028 |
| Temporal Tumors | HGG/Controls | p-Value | LGG/Controls | p-Value | HGG/LGG | p-Value |
|---|---|---|---|---|---|---|
| Global Efficiency | pSTG l | 0.010 | aSTG l | 0.027 | ||
| Local Efficiency | pMTG r | 0.044 | pSTG l | 0.003 | ||
| Betweenness Centrality | pSTG r | 0.040 | aMTG l | 0.030 | ||
| Cost | pMTG r | 0.022 | aSTG l | 0.018 | pMTG r | 0.031 |
| toMTG r | 0.032 | pSTG l | 0.008 | |||
| pSTG l | 0.032 | aMTG l | 0.017 | |||
| aITG l | 0.038 | |||||
| Clustering Coefficient | aITG r | 0.017 | ||||
| pSTG l | 0.018 | |||||
| Degree | pMTG r | 0.022 | aSTG l | 0.018 | pMTG r | 0.031 |
| toMTG r | 0.032 | pSTG l | 0.008 | |||
| pSTG l | 0.032 | aMTG l | 0.017 | |||
| aITG l | 0.038 |
| Parietal Tumors | HGG/Controls | p-Value | LGG/Controls | p-Value |
| Global Efficiency | AG l | 0.0434 | ||
| Local Efficiency | PostCG l | 0.0145 | ||
| Betweenness Centrality | PostCG l | 0.0051 | PostCG l | 0.0236 |
| Average Path Length | AG l | 0.0394 | ||
| Insular Tumors | HGG/Controls | p-Value | LGG/Controls | p-Value |
| Cost | IC l | 0.0161 | ||
| Degree | IC l | 0.0161 |
| SFG r | HGG/HC | LGG/HC | pSTG r | HGG/HC | |
|---|---|---|---|---|---|
| Local Efficiency | Global Efficiency | Betweenness | |||
| Clustering Coefficient | Betweenness | pSTG l | HGG/HC | LGG/HC | |
| Cost | Global Efficiency | Local Efficiency | |||
| Average Path Length | Cost | Cost | |||
| Degree | Degree | Clustering Coefficient | |||
| SFG l | LGG/HC | Degree | |||
| Average Path Length | aSTG l | LGG/HC | |||
| MidFG l | LGG/HC | Global Efficiency | |||
| Clustering Coefficient | Cost | ||||
| IFG tri l | HGG/HC | Degree | |||
| Betweenness | aITG l | LGG/HC | |||
| Clustering Coefficient | Cost | ||||
| pMTG r | HGG/HC | Degree | |||
| Local Efficiency | aITG r | LGG/HC | |||
| Cost | Clustering Coefficient | ||||
| Degree | AG l | HGG/HC | |||
| aMTG l | LGG/HC | Global Efficiency | |||
| Betweenness | Average Path Length | ||||
| Cost | PostCG l | HGG/HC | LGG/HC | ||
| Degree | Local Efficiency | Betweenness | |||
| Betweenness | |||||
| toMTG r | HGG/HC | IC l | LGG/HC | ||
| Cost | Cost | ||||
| Degree | Degree |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pasquini, L.; Jenabi, M.; Yildirim, O.; Silveira, P.; Peck, K.K.; Holodny, A.I. Brain Functional Connectivity in Low- and High-Grade Gliomas: Differences in Network Dynamics Associated with Tumor Grade and Location. Cancers 2022, 14, 3327. https://doi.org/10.3390/cancers14143327
Pasquini L, Jenabi M, Yildirim O, Silveira P, Peck KK, Holodny AI. Brain Functional Connectivity in Low- and High-Grade Gliomas: Differences in Network Dynamics Associated with Tumor Grade and Location. Cancers. 2022; 14(14):3327. https://doi.org/10.3390/cancers14143327
Chicago/Turabian StylePasquini, Luca, Mehrnaz Jenabi, Onur Yildirim, Patrick Silveira, Kyung K. Peck, and Andrei I. Holodny. 2022. "Brain Functional Connectivity in Low- and High-Grade Gliomas: Differences in Network Dynamics Associated with Tumor Grade and Location" Cancers 14, no. 14: 3327. https://doi.org/10.3390/cancers14143327