
The Yin and Yang of Discoidin Domain Receptors (DDRs): Implications in Tumor Growth and Metastasis Development

Abstract
:Simple Summary
Abstract
1. Introduction

2. Roles of DDRs in Cancer Progression
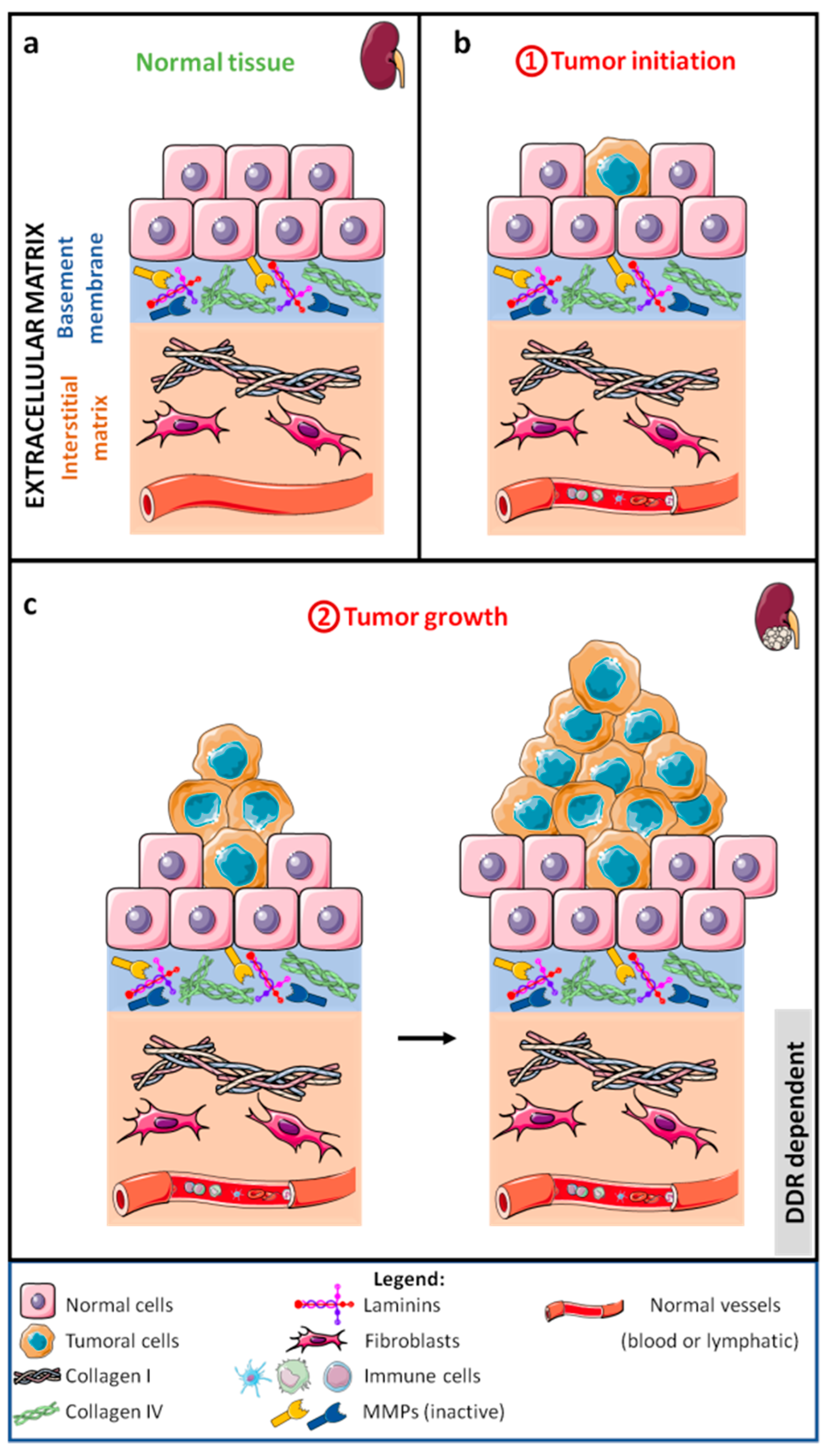
2.1. Tumor Growth
2.1.1. DDRs and Tumor Cells

| Receptor | Cell Survival or Cell Proliferation | Cancers |
|---|---|---|
| DDR1 | increase | -Sarcoma [34] -Colon carcinoma [34] -Breast carcinoma [57,58] -Hodgkin’s lymphoma [52] -Prostate carcinoma [53] -Lung (NSCLC) [54], (adenocarcinoma) [49] -Gastric carcinoma [50] -Pancreatic carcinoma [55,56] |
| decrease | -Breast carcinoma [59,60,71] -Colon carcinoma [61] | |
| DDR2 | increase | -Melanoma [62] -Oral SCC [63] -Gastric carcinoma [64,67] -Hepatocellular carcinoma [65] -Lymphoma [43] -Lung (NSCLC) [66] |
| decrease | -Melanoma [68] -Fibrosarcoma [68,73] |
2.1.2. DDRs and Immune Cells
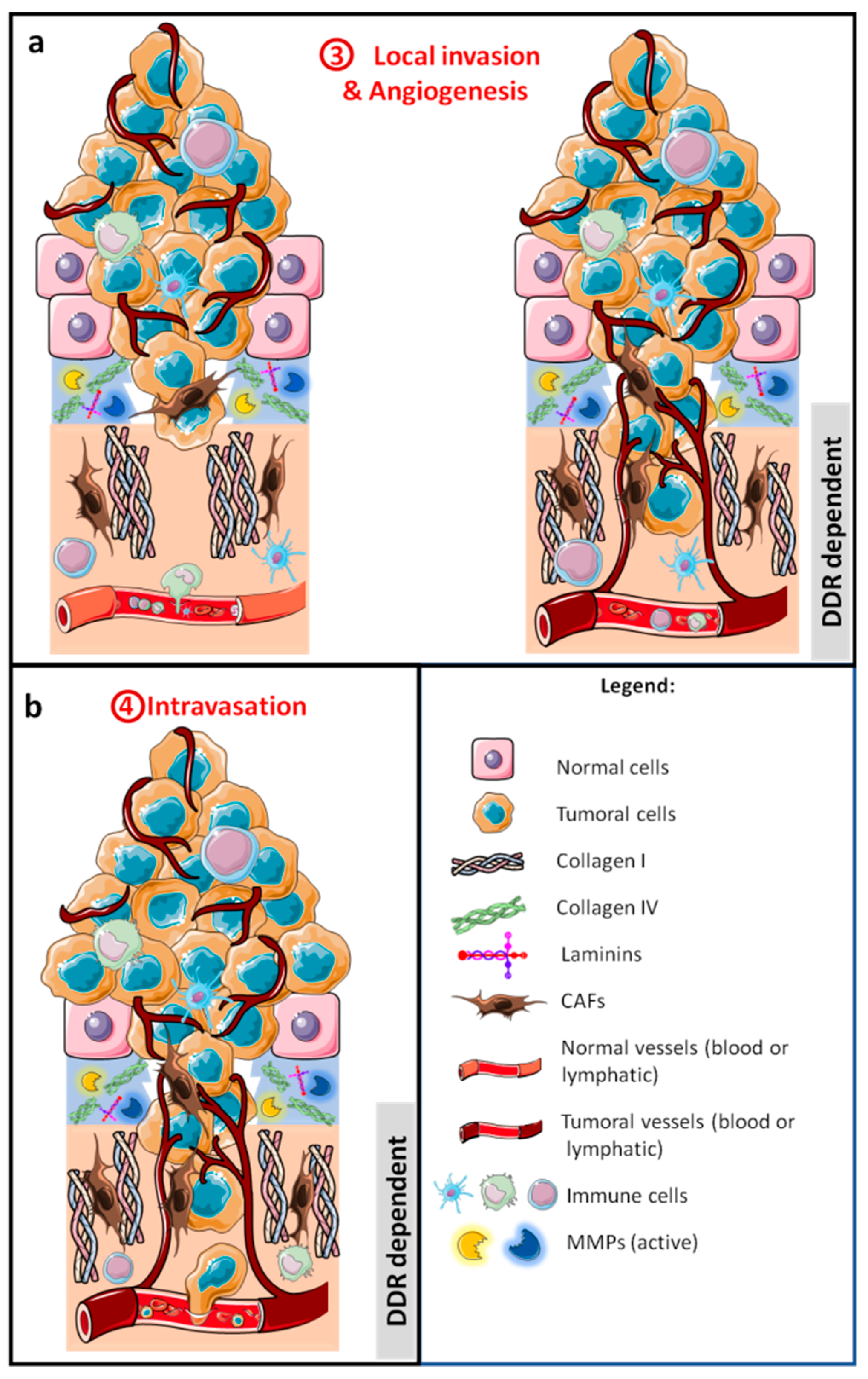
2.2. Local Invasion
2.2.1. Extracellular Matrix Breakdown
2.2.2. Epithelial–Mesenchymal Transition
2.2.3. Migration and Invasion
2.3. Intravasation
2.4. Survival in the Circulation, Extravasation, and Micrometastasis Formation
2.5. Macrometastasis (Colonization)
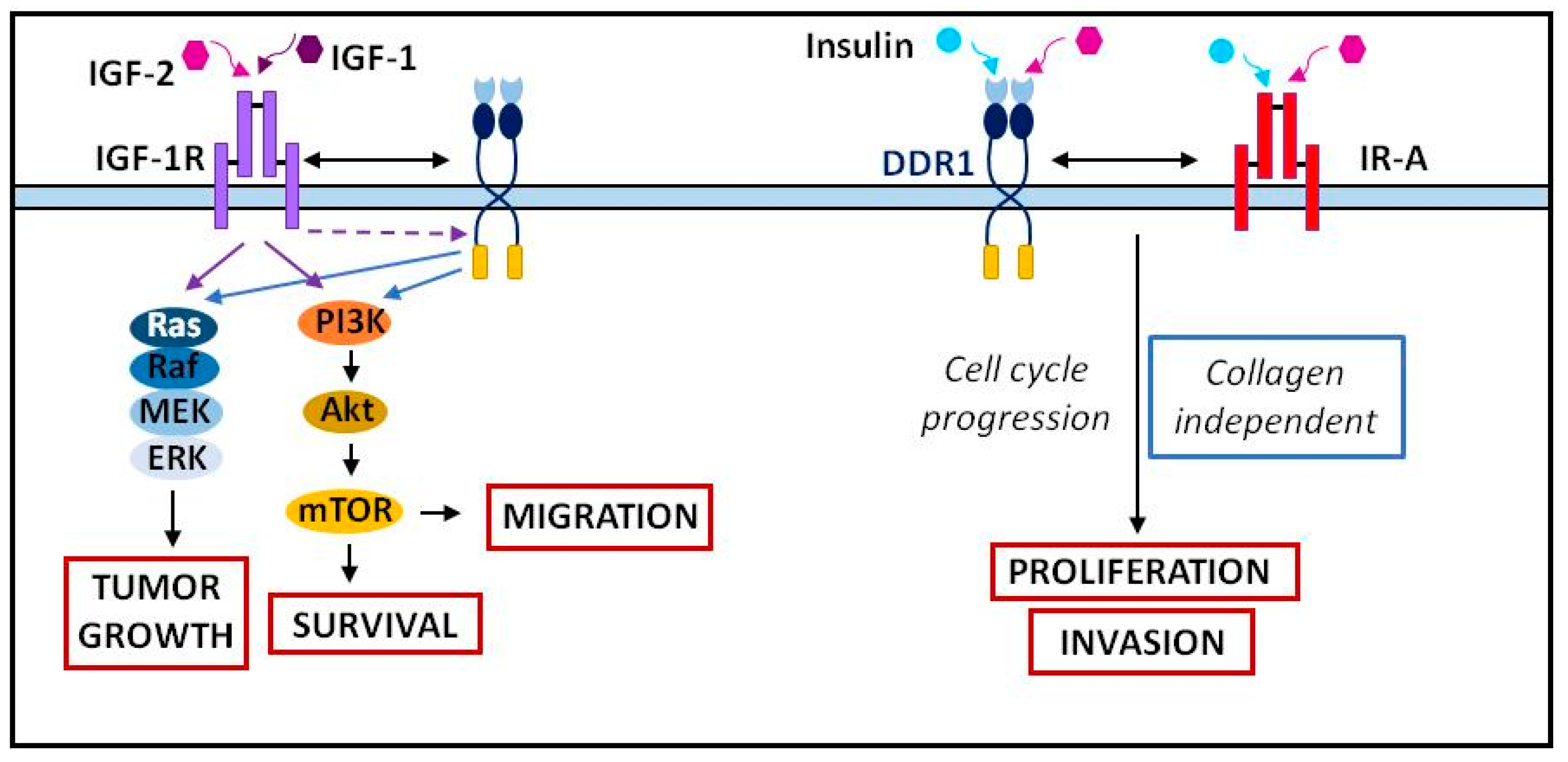
2.6. DDR Association and Crosstalk with Other Membrane Receptors

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3. Cancer Treatment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Auguste, P.; Lemiere, S.; Larrieu-Lahargue, F.; Bikfalvi, A. Molecular Mechanisms of Tumor Vascularization. Crit. Rev. Oncol. Hematol. 2005, 54, 53–61. [Google Scholar] [CrossRef]
- Fidler, I.J. Timeline—The Pathogenesis of Cancer Metastasis: The “seed and Soil” Hypothesis Revisited. Nat. Rev. Cancer 2003, 3, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Valastyan, S.; Weinberg, R.A. Tumor Metastasis: Molecular Insights and Evolving Paradigms. Cell 2011, 147, 275–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinbichler, T.B.; Savic, D.; Dudás, J.; Kvitsaridze, I.; Skvortsov, S.; Riechelmann, H.; Skvortsova, I.-I. Cancer Stem Cells and Their Unique Role in Metastatic Spread. Semin. Cancer Biol. 2020, 60, 148–156. [Google Scholar] [CrossRef]
- Niland, S.; Eble, J.A. Hold on or Cut? Integrin- and MMP-Mediated Cell-Matrix Interactions in the Tumor Microenvironment. Int. J. Mol. Sci. 2021, 22, 238. [Google Scholar] [CrossRef]
- Ricard-Blum, S. The Collagen Family. Cold Spring Harbor Perspect. Biol. 2011, 3, a004978. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Xu, H.; Wang, W.; Li, S.; Li, H.; Li, T.; Zhang, W.; Yu, X.; Liu, L. The Role of Collagen in Cancer: From Bench to Bedside. J. Transl. Med. 2019, 17, 309. [Google Scholar] [CrossRef] [Green Version]
- O’Reilly, M.S.; Boehm, T.; Shing, Y.; Fukai, N.; Vasios, G.; Lane, W.S.; Flynn, E.; Birkhead, J.R.; Olsen, B.R.; Folkman, J. Endostatin: An Endogenous Inhibitor of Angiogenesis and Tumor Growth. Cell 1997, 88, 277–285. [Google Scholar] [CrossRef] [Green Version]
- Ilina, O.; Gritsenko, P.G.; Syga, S.; Lippoldt, J.; La Porta, C.A.M.; Chepizhko, O.; Grosser, S.; Vullings, M.; Bakker, G.-J.; Starruß, J.; et al. Cell–Cell Adhesion and 3D Matrix Confinement Determine Jamming Transitions in Breast Cancer Invasion. Nat. Cell Biol. 2020, 22, 1103–1115. [Google Scholar] [CrossRef]
- Weigelin, B.; Bakker, G.-J.; Friedl, P. Intravital Third Harmonic Generation Microscopy of Collective Melanoma Cell Invasion: Principles of Interface Guidance and Microvesicle Dynamics. IntraVital 2012, 1, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Lorusso, G.; Rueegg, C.; Kuonen, F. Targeting the Extra-Cellular Matrix-Tumor Cell Crosstalk for Anti-Cancer Therapy: Emerging Alternatives to Integrin Inhibitors. Front. Oncol. 2020, 10, 1231. [Google Scholar] [CrossRef]
- Rammal, H.; Saby, C.; Magnien, K.; Van-Gulick, L.; Garnotel, R.; Buache, E.; El Btaouri, H.; Jeannesson, P.; Morjani, H. Corrigendum: Discoidin Domain Receptors: Potential Actors and Targets in Cancer. Front. Pharmacol. 2016, 7, 346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, H.-L.; Valiathan, R.R.; Arkwright, R.; Sohail, A.; Mihai, C.; Kumarasiri, M.; Mahasenan, K.V.; Mobashery, S.; Huang, P.; Agarwal, G.; et al. Discoidin Domain Receptors: Unique Receptor Tyrosine Kinases in Collagen-Mediated Signaling. J. Biol. Chem. 2013, 288, 7430–7437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, T.; Mignatti, P.; Abramson, S.B.; Attur, M. Periostin Interaction with Discoidin Domain Receptor-1 (DDR1) Promotes Cartilage Degeneration. PLoS ONE 2020, 15, e0231501. [Google Scholar] [CrossRef] [PubMed]
- Corcoran, D.S.; Juskaite, V.; Xu, Y.; Gorlitz, F.; Alexandrov, Y.; Dunsby, C.; French, P.M.W.; Leitinger, B. DDR1 Autophosphorylation Is a Result of Aggregation into Dense Clusters. Sci. Rep. 2019, 9, 17104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juskaite, V.; Corcoran, D.S.; Leitinger, B. Collagen Induces Activation of DDR1 through Lateral Dimer Association and Phosphorylation between Dimers. eLife 2017, 6, e25716. [Google Scholar] [CrossRef] [PubMed]
- Leitinger, B. Discoidin Domain Receptor Functions in Physiological and Pathological Conditions. In International Review of Cell and Molecular Biology; Jeon, K.W., Ed.; Elsevier Academic Press Inc.: San Diego, CA, USA, 2014; Volume 310, pp. 39–87. ISBN 978-0-12-800180-6. [Google Scholar]
- Yeung, D.A.; Shanker, N.; Sohail, A.; Weiss, B.A.; Wang, C.; Wellmerling, J.; Das, S.; Ganju, R.K.; Miller, J.L.C.; Herr, A.B.; et al. Clustering, Spatial Distribution, and Phosphorylation of Discoidin Domain Receptors 1 and 2 in Response to Soluble Collagen I. J. Mol. Biol. 2019, 431, 368–390. [Google Scholar] [CrossRef]
- Agarwal, G.; Smith, A.W.; Jones, B. Discoidin Domain Receptors: Micro Insights into Macro Assemblies. Biochim. Biophys. Acta-Mol. Cell Res. 2019, 1866, 118496. [Google Scholar] [CrossRef]
- Yang, K.; Kim, J.H.; Kim, H.J.; Park, I.S.; Kim, I.Y.; Yang, B.S. Tyrosine 740 Phosphorylation of Discoidin Domain Receptor 2 by Src Stimulates Intramolecular Autophosphorylation and Shc Signaling Complex Formation. J. Biol. Chem. 2005, 280, 39058–39066. [Google Scholar] [CrossRef] [Green Version]
- Croissant, C.; Tuariihionoa, A.; Bacou, M.; Souleyreau, W.; Sala, M.; Henriet, E.; Bikfalvi, A.; Saltel, F.; Auguste, P. DDR1 and DDR2 Physical Interaction Leads to Signaling Interconnection but with Possible Distinct Functions. Cell Adhes. Migr. 2018, 12, 324–334. [Google Scholar] [CrossRef] [Green Version]
- Hilton, H.N.; Stanford, P.M.; Harris, J.; Oakes, S.R.; Kaplan, W.; Daly, R.J.; Ormandy, C.J. KIBRA Interacts with Discoidin Domain Receptor 1 to Modulate Collagen-Induced Signalling. Biochim. Biophys. Acta Mol. Cell Res. 2008, 1783, 383–393. [Google Scholar] [CrossRef] [Green Version]
- Wasinski, B.; Sohail, A.; Bonfil, R.D.; Kim, S.; Saliganan, A.; Polin, L.; Bouhamdan, M.; Kim, H.-R.C.; Prunotto, M.; Fridman, R. Discoidin Domain Receptors, DDR1b and DDR2, Promote Tumour Growth within Collagen but DDR1b Suppresses Experimental Lung Metastasis in HT1080 Xenografts. Sci. Rep. 2020, 10, 2309. [Google Scholar] [CrossRef] [PubMed]
- Juin, A.; Di Martino, J.; Leitinger, B.; Henriet, E.; Gary, A.-S.; Paysan, L.; Bomo, J.; Baffet, G.; Gauthier-Rouvière, C.; Rosenbaum, J.; et al. Discoidin Domain Receptor 1 Controls Linear Invadosome Formation via a Cdc42-Tuba Pathway. J. Cell Biol. 2014, 207, 517–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hidalgo-Carcedo, C.; Hooper, S.; Chaudhry, S.I.; Williamson, P.; Harrington, K.; Leitinger, B.; Sahai, E. Collective Cell Migration Requires Suppression of Actomyosin at Cell–Cell Contacts Mediated by DDR1 and the Cell Polarity Regulators Par3 and Par6. Nat. Cell Biol. 2011, 13, 49–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, H.; Chakraborty, G.; Zhang, Z.; Akalay, I.; Gadiya, M.; Gao, Y.; Sinha, S.; Hu, J.; Jiang, C.; Akram, M.; et al. Multi-Organ Site Metastatic Reactivation Mediated by Non-Canonical Discoidin Domain Receptor 1 Signaling. Cell 2016, 166, 47–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, E.-H.; Li, H.-S.; Zhao, T.; Fan, J.-D.; Ma, X.; Xiong, L.; Li, W.-J.; Zhu, L.-L.; Fan, M. Effect of Hypoxia on the Gene Profile of Human Bone Marrow-Derived Mesenchymal Stem Cells. Sheng Li Xue Bao Acta Physiol. Sin. 2007, 59, 227–232. [Google Scholar]
- Chen, S.-C.; Wang, B.-W.; Wang, D.L.; Shyu, K.-G. Hypoxia Induces Discoidin Domain Receptor-2 Expression via the P38 Pathway in Vascular Smooth Muscle Cells to Increase Their Migration. Biochem. Biophys. Res. Commun. 2008, 374, 662–667. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhang, Z.; Xue, J.; Guo, X.; Liang, S.; Liu, A. Effect of Hypoxia on DDR1 Expression in Pituitary Adenomas. Med. Sci. Monit. 2015, 21, 2433–2438. [Google Scholar] [CrossRef] [Green Version]
- Ren, T.; Zhang, W.; Liu, X.; Zhao, H.; Zhang, J.; Zhang, J.; Li, X.; Zhang, Y.; Bu, X.; Shi, M.; et al. Discoidin Domain Receptor 2 (DDR2) Promotes Breast Cancer Cell Metastasis and the Mechanism Implicates Epithelial-Mesenchymal Transition Programme under Hypoxia. J. Pathol. 2014, 234, 526–537. [Google Scholar] [CrossRef]
- Koh, M.; Woo, Y.; Valiathan, R.R.; Jung, H.Y.; Park, S.Y.; Kim, Y.N.; Kim, H.-R.C.; Fridman, R.; Moon, A. Discoidin Domain Receptor 1 Is a Novel Transcriptional Target of ZEB1 in Breast Epithelial Cells Undergoing H-Ras-Induced Epithelial to Mesenchymal Transition. Int. J. Cancer 2015, 136, E508–E520. [Google Scholar] [CrossRef] [Green Version]
- Grither, W.R.; Divine, L.M.; Meller, E.H.; Wilke, D.J.; Desai, R.A.; Loza, A.J.; Zhao, P.; Lohrey, A.; Longmore, G.D.; Fuh, K.C. TWIST1 Induces Expression of Discoidin Domain Receptor 2 to Promote Ovarian Cancer Metastasis. Oncogene 2018, 37, 1714–1729. [Google Scholar] [CrossRef]
- Wang, Z.; Sun, X.; Bao, Y.; Mo, J.; Du, H.; Hu, J.; Zhang, X. E2F1 Silencing Inhibits Migration and Invasion of Osteosarcoma Cells via Regulating DDR1 Expression. Int. J. Oncol. 2017, 51, 1639–1650. [Google Scholar] [CrossRef] [Green Version]
- Ongusaha, P.P.; Kim, J.; Fang, L.; Wong, T.W.; Yancopoulos, G.D.; Aaronson, S.A.; Lee, S.W. P53 Induction and Activation of DDR1 Kinase Counteract P53-Mediated Apoptosis and Influence P53 Regulation through a Positive Feedback Loop. EMBO J. 2003, 22, 1289–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.; Jin, H.; Wu, X.; Jian, Z.; Zou, X.; Huang, J.; Guan, R.; Wei, X. The Cross-Talk between DDR1 and STAT3 Promotes the Development of Hepatocellular Carcinoma. Aging 2020, 12, 14391–14405. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.-L.; Chou, C.-H.; Hsieh, S.-C.; Hwa, S.-Y.; Lee, M.-T.; Wang, F.-F. Transcriptional Upregulation of DDR2 by ATF4 Facilitates Osteoblastic Differentiation through P38 MAPK-Mediated Runx2 Activation. J. Bone Miner. Res. 2010, 25, 2489–2503. [Google Scholar] [CrossRef] [PubMed]
- George, M.; Vijayakumar, A.; Dhanesh, S.B.; James, J.; Shivakumar, K. Molecular Basis and Functional Significance of Angiotensin II-Induced Increase in Discoidin Domain Receptor 2 Gene Expression in Cardiac Fibroblasts. J. Mol. Cell. Cardiol. 2016, 90, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.; Ehsanipour, A.; Hsu, J.J.; Lu, J.; Pedego, T.; Wu, A.; Walthers, C.M.; Demer, L.L.; Seidlits, S.K.; Tintut, Y. Inflammation Drives Retraction, Stiffening, and Nodule Formation via Cytoskeletal Machinery in a Three-Dimensional Culture Model of Aortic Stenosis. Am. J. Pathol. 2016, 186, 2378–2389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ezzoukhry, Z.; Henriet, E.; Piquet, L.; Boyé, K.; Bioulac-Sage, P.; Balabaud, C.; Couchy, G.; Zucman-Rossi, J.; Moreau, V.; Saltel, F. TGF-Β1 Promotes Linear Invadosome Formation in Hepatocellular Carcinoma Cells, through DDR1 up-Regulation and Collagen I Cross-Linking. Eur. J. Cell Biol. 2016, 95, 503–512. [Google Scholar] [CrossRef]
- Ruiz, P.A.; Jarai, G. Collagen I Induces Discoidin Domain Receptor (DDR) 1 Expression through DDR2 and a JAK2-ERK1/2-Mediated Mechanism in Primary Human Lung Fibroblasts. J. Biol. Chem. 2011, 286, 12912–12923. [Google Scholar] [CrossRef] [Green Version]
- Krazinski, B.E.; Kiewisz, J.; Sliwinska-Jewsiewicka, A.; Kowalczyk, A.E.; Grzegrzolka, J.; Godlewski, J.; Kwiatkowski, P.; Dziegiel, P.; Kmiec, Z. Altered Expression of DDR1 in Clear Cell Renal Cell Carcinoma Correlates With MiR-199a/b-5p and Patients’ Outcome. Cancer Genom. Proteom. 2019, 16, 179–193. [Google Scholar] [CrossRef] [Green Version]
- Chou, S.-T.; Peng, H.-Y.; Mo, K.-C.; Hsu, Y.-M.; Wu, G.-H.; Hsiao, J.-R.; Lin, S.-F.; Wang, H.-D.; Shiah, S.-G. MicroRNA-486-3p Functions as a Tumor Suppressor in Oral Cancer by Targeting DDR1. J. Exp. Clin. Cancer Res. 2019, 38, 281. [Google Scholar] [CrossRef]
- Deng, L.; Liu, G.; Zheng, C.; Zhang, L.; Kang, Y.; Yang, F. Circ-LAMP1 Promotes T-Cell Lymphoblastic Lymphoma Progression via Acting as a CeRNA for MiR-615-5p to Regulate DDR2 Expression. Gene 2019, 701, 146–151. [Google Scholar] [CrossRef]
- Ramasamy, R.; Ridgeway, A.; Lipshultz, L.I.; Lamb, D.J. Integrative DNA Methylation and Gene Expression Analysis Identifies Discoidin Domain Receptor 1 Association with Idiopathic Nonobstructive Azoospermia. Fertil. Steril. 2014, 102, 968–973.e3. [Google Scholar] [CrossRef] [Green Version]
- Chung, V.Y.; Tan, T.Z.; Huang, R.-L.; Lai, H.-C.; Huang, R.Y.-J. Loss of Discoidin Domain Receptor 1 (DDR1) via CpG Methylation during EMT in Epithelial Ovarian Cancer. Gene 2017, 635, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Nelson, H.H.; Marsit, C.J.; Christensen, B.C.; Houseman, E.A.; Kontic, M.; Wiemels, J.L.; Karagas, M.R.; Wrensch, M.R.; Zheng, S.; Wiencke, J.K.; et al. Key Epigenetic Changes Associated with Lung Cancer Development. Epigenetics 2012, 7, 559–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, W.F. Ligand-Induced Shedding of Discoidin Domain Receptor 1. FEBS Lett. 2002, 514, 175–180. [Google Scholar] [CrossRef] [Green Version]
- Shitomi, Y.; Thogersen, I.B.; Ito, N.; Leitinger, B.; Enghild, J.J.; Itoh, Y. ADAM10 Controls Collagen Signaling and Cell Migration on Collagen by Shedding the Ectodomain of Discoidin Domain Receptor 1 (DDR1). Mol. Biol. Cell 2015, 26, 659–673. [Google Scholar] [CrossRef] [PubMed]
- Ambrogio, C.; Gómez-López, G.; Falcone, M.; Vidal, A.; Nadal, E.; Crosetto, N.; Blasco, R.B.; Fernández-Marcos, P.J.; Sánchez-Céspedes, M.; Ren, X.; et al. Combined Inhibition of DDR1 and Notch Signaling Is a Therapeutic Strategy for KRAS-Driven Lung Adenocarcinoma. Nat. Med. 2016, 22, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Hur, H.; Ham, I.-H.; Lee, D.; Jin, H.; Aguilera, K.Y.; Oh, H.J.; Han, S.-U.; Kwon, J.E.; Kim, Y.-B.; Ding, K.; et al. Discoidin Domain Receptor 1 Activity Drives an Aggressive Phenotype in Gastric Carcinoma. BMC Cancer 2017, 17, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azemikhah, M.; Ashtiani, H.A.; Aghaei, M.; Rastegar, H. Evaluation of Discoidin Domain Receptor-2 (DDR2) Expression Level in Normal, Benign, and Malignant Human Prostate Tissues. Res. Pharm. Sci. 2015, 10, 356–363. [Google Scholar] [PubMed]
- Cader, F.Z.; Vockerodt, M.; Bose, S.; Nagy, E.; Brundler, M.-A.; Kearns, P.; Murray, P.G. The EBV Oncogene LMP1 Protects Lymphoma Cells from Cell Death through the Collagen-Mediated Activation of DDR. Blood 2013, 122, 10. [Google Scholar] [CrossRef] [Green Version]
- Shimada, K.; Nakamura, M.; Ishida, E.; Higuchi, T.; Yamamoto, H.; Tsujikawa, K.; Konishi, N. Prostate Cancer Antigen-1 Contributes to Cell Survival and Invasion Though Discoidin Receptor 1 in Human Prostate Cancer. Cancer Sci. 2008, 99, 39–45. [Google Scholar] [CrossRef]
- Villalba, M.; Redin, E.; Exposito, F.; Pajares, M.J.; Sainz, C.; Hervas, D.; Guruceaga, E.; Diaz-Lagares, A.; Cirauqui, C.; Redrado, M.; et al. Identification of a Novel Synthetic Lethal Vulnerability in Non-Small Cell Lung Cancer by Co-Targeting TMPRSS4 and DDR1. Sci. Rep. 2019, 9, 15400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruggeri, J.M.; Franco-Barraza, J.; Sohail, A.; Zhang, Y.; Long, D.; Pasca di Magliano, M.; Cukierman, E.; Fridman, R.; Crawford, H.C. Discoidin Domain Receptor 1 (DDR1) Is Necessary for Tissue Homeostasis in Pancreatic Injury and Pathogenesis of Pancreatic Ductal Adenocarcinoma. Am. J. Pathol. 2020, 190, 1735–1751. [Google Scholar] [CrossRef]
- Rudra-Ganguly, N.; Lowe, C.; Mattie, M.; Chang, M.S.; Satpayev, D.; Verlinsky, A.; An, Z.; Hu, L.; Yang, P.; Challita-Eid, P.; et al. Discoidin Domain Receptor 1 Contributes to Tumorigenesis through Modulation of TGFBI Expression. PLoS ONE 2014, 9, e111515. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Duan, L.; Luo, J.; Zhang, L.; Lu, X.; Zhang, Y.; Zhang, Z.; Tu, Z.; Xu, Y.; Ren, X.; et al. Discovery and Optimization of 3-(2-(Pyrazolo[1,5- a ]Pyrimidin-6-Yl)Ethynyl)Benzamides as Novel Selective and Orally Bioavailable Discoidin Domain Receptor 1 (DDR1) Inhibitors. J. Med. Chem. 2013, 56, 3281–3295. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.; Chen, Y.; Liu, Y.; Lai, Y.; Liu, D. MiR-199b-5p Inhibits Triple Negative Breast Cancer Cell Proliferation, Migration and Invasion by Targeting DDR1. Oncol. Lett. 2018, 16, 4889–4896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assent, D.; Bourgot, I.; Hennuy, B.; Geurts, P.; Noel, A.; Foidart, J.-M.; Maquoi, E. A Membrane-Type-1 Matrix Metalloproteinase (MT1-MMP)—Discoidin Domain Receptor 1 Axis Regulates Collagen-Induced Apoptosis in Breast Cancer Cells. PLoS ONE 2015, 10, e0116006. [Google Scholar] [CrossRef]
- Saby, C.; Collin, G.; Sinane, M.; Buache, E.; Van Gulick, L.; Saltel, F.; Maquoi, E.; Morjani, H. DDR1 and MT1-MMP Expression Levels Are Determinant for Triggering BIK-Mediated Apoptosis by 3D Type I Collagen Matrix in Invasive Basal-Like Breast Carcinoma Cells. Front. Pharmacol. 2019, 10, 462. [Google Scholar] [CrossRef]
- Appert-Collin, A. LRP-1 Promotes Colon Cancer Cell Proliferation in 3D Collagen Matrices by Mediating DDR1 Endocytosis. Front. Cell Dev. Biol. 2020, 8, 15. [Google Scholar]
- Badiola, I.; Villacé, P.; Basaldua, I.; Olaso, E. Downregulation of Discoidin Domain Receptor 2 in A375 Human Melanoma Cells Reduces Its Experimental Liver Metastasis Ability. Oncol. Rep. 2011, 26, 971–978. [Google Scholar]
- Sun, M.; Shen, Z. Knockdown of Long Non-Coding RNA (LncRNA) Colon Cancer-Associated Transcript-1 (CCAT1) Suppresses Oral Squamous Cell Carcinoma Proliferation, Invasion, and Migration by Inhibiting the Discoidin Domain Receptor 2 (DDR2)/ERK/AKT Axis. Med. Sci. Monit. 2020, 26, e920020. [Google Scholar] [CrossRef] [PubMed]
- Kurashige, J.; Hasegawa, T.; Niida, A.; Sugimachi, K.; Deng, N.; Mima, K.; Uchi, R.; Sawada, G.; Takahashi, Y.; Eguchi, H.; et al. Integrated Molecular Profiling of Human Gastric Cancer Identifies DDR2 as a Potential Regulator of Peritoneal Dissemination. Sci. Rep. 2016, 6, 22371. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-W.; Lee, Y.-S.; Kim, J.S.; Lee, S.-K.; Kim, B.H.; Lee, J.A.; Lee, N.O.; Kim, S.H.; Hong, E.K. Downregulation of Discoidin Domain Receptor 2 Decreases Tumor Growth of Hepatocellular Carcinoma. J. Cancer Res. Clin. Oncol. 2015, 141, 1973–1983. [Google Scholar] [CrossRef]
- Kim, D.; You, E.; Jeong, J.; Ko, P.; Kim, J.-W.; Rhee, S. DDR2 Controls the Epithelial-Mesenchymal-Transition-Related Gene Expression via c-Myb Acetylation upon Matrix Stiffening. Sci. Rep. 2017, 7, 6847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.-G.; Xu, L.; Jia, R.-R.; Wu, Q.; Wang, T.; Wei, J.; Ma, J.-L.; Shi, M.; Li, Z.-S. DDR2 Induces Gastric Cancer Cell Activities via Activating MTORC2 Signaling and Is Associated with Clinicopathological Characteristics of Gastric Cancer. Dig. Dis. Sci. 2016, 61, 2272–2283. [Google Scholar] [CrossRef]
- Wall, S.J.; Werner, E.; Werb, Z.; DeClerck, Y.A. Discoidin Domain Receptor 2 Mediates Tumor Cell Cycle Arrest Induced by Fibrillar Collagen. J. Biol. Chem. 2005, 280, 40187–40194. [Google Scholar] [CrossRef] [Green Version]
- Terashima, M.; Togashi, Y.; Sato, K.; Mizuuchi, H.; Sakai, K.; Suda, K.; Nakamura, Y.; Banno, E.; Hayashi, H.; De Velasco, M.A.; et al. Functional Analyses of Mutations in Receptor Tyrosine Kinase Genes in Non–Small Cell Lung Cancer: Double-Edged Sword of DDR2. Clin. Cancer Res. 2016, 22, 3663–3671. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.-S.; Jung, E.A.; An, S.B.; Kim, Y.J.; Oh, D.-Y.; Song, J.-Y.; Um, S.-W.; Han, J.; Choi, Y.-L. Prevalence of Mutations in Discoidin Domain-Containing Receptor Tyrosine Kinase 2 (DDR2) in Squamous Cell Lung Cancers in Korean Patients. Cancer Res. Treat. 2017, 49, 1065–1076. [Google Scholar] [CrossRef] [Green Version]
- Saby, C.; Rammal, H.; Magnien, K.; Buache, E.; Brassart-Pasco, S.; Van-Gulick, L.; Jeannesson, P.; Maquoi, E.; Morjani, H. Age-Related Modifications of Type I Collagen Impair DDR1-Induced Apoptosis in Non-Invasive Breast Carcinoma Cells. Cell Adhes. Migr. 2018, 12, 335–347. [Google Scholar] [CrossRef] [Green Version]
- Jin, H.; Ham, I.-H.; Oh, H.J.; Bae, C.A.; Lee, D.; Kim, Y.-B.; Son, S.-Y.; Chwae, Y.-J.; Han, S.-U.; Brekken, R.A.; et al. Inhibition of Discoidin Domain Receptor 1 Prevents Stroma-Induced Peritoneal Metastasis in Gastric Carcinoma. Mol. Cancer Res. 2018, 16, 1590–1600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saby, C.; Buache, E.; Brassart-Pasco, S.; El Btaouri, H.; Courageot, M.-P.; Van Gulick, L.; Garnotel, R.; Jeannesson, P.; Morjani, H. Type I Collagen Aging Impairs Discoidin Domain Receptor 2-Mediated Tumor Cell Growth Suppression. Oncotarget 2016, 7, 24908–24927. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azreq, M.-A.E.; Kadiri, M.; Boisvert, M.; Pagé, N.; Tessier, P.A.; Aoudjit, F. Discoidin Domain Receptor 1 Promotes Th17 Cell Migration by Activating the RhoA/ROCK/MAPK/ERK Signaling Pathway. Oncotarget 2016, 7, 44975–44990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadiri, M.; El Azreq, M.-A.; Berrazouane, S.; Boisvert, M.; Aoudjit, F. Human Th17 Migration in Three-Dimensional Collagen Involves P38 MAPK: Th17 M IGRATION R EQUIRES P38. J. Cell. Biochem. 2017, 118, 2819–2827. [Google Scholar] [CrossRef]
- Zhong, X.; Zhang, W.; Sun, T. DDR1 Promotes Breast Tumor Growth by Suppressing Antitumor Immunity. Oncol. Rep. 2019, 42, 2844–2854. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-E.; Kang, C.-S.; Guan, X.-Y.; Kim, B.-T.; Kim, S.-H.; Lee, Y.-M.; Moon, W.-S.; Kim, D.-K. Discoidin Domain Receptor 2 Is Involved in the Activation of Bone Marrow-Derived Dendritic Cells Caused by Type I Collagen. Biochem. Biophys. Res. Commun. 2007, 352, 244–250. [Google Scholar] [CrossRef]
- Afonso, P.V.; McCann, C.P.; Kapnick, S.M.; Parent, C.A. Discoidin Domain Receptor 2 Regulates Neutrophil Chemotaxis in 3D Collagen Matrices. Blood 2013, 121, 1644–1650. [Google Scholar] [CrossRef] [Green Version]
- Sidles, S.J.; Xiong, Y.; Young, M.R.I.; LaRue, A.C. High-Fat Diet Alters Immunogenic Properties of Circulating and Adipose Tissue-Associated Myeloid-Derived CD45 + DDR2 + Cells. Mediat. Inflamm. 2019, 2019, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.; Chaudhuri, O. Beyond Proteases: Basement Membrane Mechanics and Cancer Invasion. J. Cell Biol. 2019, 218, 2456–2469. [Google Scholar] [CrossRef] [Green Version]
- Valencia, K.; Ormazabal, C.; Zandueta, C.; Luis-Ravelo, D.; Anton, I.; Pajares, M.J.; Agorreta, J.; Montuenga, L.M.; Martinez-Canarias, S.; Leitinger, B.; et al. Inhibition of Collagen Receptor Discoidin Domain Receptor-1 (DDR1) Reduces Cell Survival, Homing, and Colonization in Lung Cancer Bone Metastasis. Clin. Cancer Res. 2012, 18, 969–980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, D.; Teramoto, A. Enhancement of Pituitary Adenoma Cell Invasion and Adhesion Is Mediated by Discoidin Domain Receptor-1. J. Neurooncol. 2007, 82, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Liu, J.; Jiang, B.; Chen, J.; Fu, Z.; Bai, F.; Jiang, J.; Tang, Z. MiR-199a-5p Loss Up-Regulated DDR1 Aggravated Colorectal Cancer by Activating Epithelial-to-Mesenchymal Transition Related Signaling. Dig. Dis. Sci. 2014, 59, 2163–2172. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Chen, X.; Bai, J.; Liu, Q.; Li, H.; Xie, J.; Jing, H.; Zheng, J. Discoidin Domain Receptor 1 (DDR1), a Promising Biomarker, Induces Epithelial to Mesenchymal Transition in Renal Cancer Cells. Tumor Biol. 2016, 37, 11509–11521. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, Y.; He, S.; Li, M.; Cai, X.; Wang, H.; Xu, L.; Cao, J. TM4SF1 Promotes Metastasis of Pancreatic Cancer via Regulating the Expression of DDR1. Sci. Rep. 2017, 7, 45895. [Google Scholar] [CrossRef]
- Lee, J.-H.; Poudel, B.; Ki, H.-H.; Nepali, S.; Lee, Y.-M.; Shin, J.-S.; Kim, D.-K. Complement C1q Stimulates the Progression of Hepatocellular Tumor through the Activation of Discoidin Domain Receptor 1. Sci. Rep. 2018, 8, 4908. [Google Scholar] [CrossRef]
- Bulla, R.; Tripodo, C.; Rami, D.; Ling, G.S.; Agostinis, C.; Guarnotta, C.; Zorzet, S.; Durigutto, P.; Botto, M.; Tedesco, F. C1q Acts in the Tumour Microenvironment as a Cancer-Promoting Factor Independently of Complement Activation. Nat. Commun. 2016, 7, 10346. [Google Scholar] [CrossRef] [Green Version]
- Xie, B.; Lin, W.; Ye, J.; Wang, X.; Zhang, B.; Xiong, S.; Li, H.; Tan, G. DDR2 Facilitates Hepatocellular Carcinoma Invasion and Metastasis via Activating ERK Signaling and Stabilizing SNAIL1. J. Exp. Clin. Cancer Res. 2015, 34, 101. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Corsa, C.A.; Ponik, S.M.; Prior, J.L.; Piwnica-Worms, D.; Eliceiri, K.W.; Keely, P.J.; Longmore, G.D. The Collagen Receptor Discoidin Domain Receptor 2 Stabilizes SNAIL1 to Facilitate Breast Cancer Metastasis. Nat. Cell Biol. 2013, 15, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Majkowska, I.; Shitomi, Y.; Ito, N.; Gray, N.S.; Itoh, Y. Discoidin Domain Receptor 2 Mediates Collagen-Induced Activation of Membrane-Type 1 Matrix Metalloproteinase in Human Fibroblasts. J. Biol. Chem. 2017, 292, 6633–6643. [Google Scholar] [CrossRef] [Green Version]
- Genot, E.; Gligorijevic, B. Invadosomes in Their Natural Habitat. Eur. J. Cell Biol. 2014, 93, 367–379. [Google Scholar] [CrossRef] [Green Version]
- Hammoud, A.A.; Marais, S.; Allain, N.; Ezzoukhry, Z.; Moreau, V.; Saltel, F. DDR2 Induces Linear Invadosome to Promote Angiogenesis in a Fibrillar Type I Collagen Context. Cell Biol. 2020. [Google Scholar] [CrossRef] [Green Version]
- Conklin, M.W.; Eickhoff, J.C.; Riching, K.M.; Pehlke, C.A.; Eliceiri, K.W.; Provenzano, P.P.; Friedl, A.; Keely, P.J. Aligned Collagen Is a Prognostic Signature for Survival in Human Breast Carcinoma. Am. J. Pathol. 2011, 178, 1221–1232. [Google Scholar] [CrossRef]
- Provenzano, P.P.; Eliceiri, K.W.; Campbell, J.M.; Inman, D.R.; White, J.G.; Keely, P.J. Collagen Reorganization at the Tumor-Stromal Interface Facilitates Local Invasion. BMC Med. 2006, 4, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coelho, N.M.; Arora, P.D.; van Putten, S.; Boo, S.; Petrovic, P.; Lin, A.X.; Hinz, B.; McCulloch, C.A. Discoidin Domain Receptor 1 Mediates Myosin-Dependent Collagen Contraction. Cell Rep. 2017, 18, 1774–1790. [Google Scholar] [CrossRef] [Green Version]
- Bayer, S.V.; Grither, W.R.; Brenot, A.; Hwang, P.Y.; Barcus, C.E.; Ernst, M.; Pence, P.; Walter, C.; Pathak, A.; Longmore, G.D. DDR2 Controls Breast Tumor Stiffness and Metastasis by Regulating Integrin Mediated Mechanotransduction in CAFs. eLife 2019, 8, e45508. [Google Scholar] [CrossRef] [PubMed]
- Slocum, E.; Craig, A.; Villanueva, A.; Germain, D. Parity Predisposes Breasts to the Oncogenic Action of PAPP-A and Activation of the Collagen Receptor DDR2. Breast Cancer Res. 2019, 21, 56. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, M.E.; Martin, E.E.; Anwar, T.; Arellano-Garcia, C.; Medhora, N.; Lama, A.; Chen, Y.-C.; Tanager, K.S.; Yoon, E.; Kidwell, K.M.; et al. Mesenchymal Stem Cell-Induced DDR2 Mediates Stromal-Breast Cancer Interactions and Metastasis Growth. Cell Rep. 2017, 18, 1215–1228. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Weinberg, R.A. Epithelial-to-Mesenchymal Transition in Cancer: Complexity and Opportunities. Front. Med. 2018, 12, 361–373. [Google Scholar] [CrossRef] [Green Version]
- Azizi, R.; Salemi, Z.; Fallahian, F.; Aghaei, M. Inhibition of Didscoidin Domain Receptor 1 Reduces Epithelial–Mesenchymal Transition and Induce Cell-cycle Arrest and Apoptosis in Prostate Cancer Cell Lines. J. Cell. Physiol. 2019, 234, 19539–19552. [Google Scholar] [CrossRef]
- Xie, R.; Wang, X.; Qi, G.; Wu, Z.; Wei, R.; Li, P.; Zhang, D. DDR1 Enhances Invasion and Metastasis of Gastric Cancer via Epithelial-Mesenchymal Transition. Tumor Biol. 2016, 37, 12049–12059. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Jiang, M.; Guo, S.; Cai, P.; Wang, W.; Li, Y. Promotion of MiR-221-5p on the Sensitivity of Gastric Cancer Cells to Cisplatin and Its Effects on Cell Proliferation and Apoptosis by Regulating DDR1. OTT 2020, 13, 2333–2345. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Lee, S.; Lee, E.; Lim, H.; Shin, J.; Jung, J.; Kim, S.; Moon, A. Sex-biased Differences in the Correlation between Epithelial-to-mesenchymal Transition-associated Genes in Cancer Cell Lines. Oncol. Lett. 2019. [Google Scholar] [CrossRef] [PubMed]
- Takai, K.; Drain, A.P.; Lawson, D.A.; Littlepage, L.E.; Karpuj, M.; Kessenbrock, K.; Le, A.; Inoue, K.; Weaver, V.M.; Werb, Z. Discoidin Domain Receptor 1 (DDR1) Ablation Promotes Tissue Fibrosis and Hypoxia to Induce Aggressive Basal-like Breast Cancers. Genes Dev. 2018, 32, 244–257. [Google Scholar] [CrossRef] [PubMed]
- Maeyama, M.; Koga, H.; Selvendiran, K.; Yanagimoto, C.; Hanada, S.; Taniguchi, E.; Kawaguchi, T.; Harada, M.; Ueno, T.; Sata, M. Switching in Discoid Domain Receptor Expressions in SLUG-Induced Epithelial-Mesenchymal Transition. Cancer 2008, 113, 2823–2831. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Xie, W.; Zhao, M.; Cheng, G.; Wu, M. DDR2 Facilitates Papillary Thyroid Carcinoma Epithelial Mesenchymal Transition by Activating ERK2/Snail1 Pathway. Oncol. Lett. 2017, 14, 8114–8121. [Google Scholar] [CrossRef] [Green Version]
- Friedl, P.; Wolf, K. Plasticity of Cell Migration: A Multiscale Tuning Model. J. Cell. Biol. 2010, 188, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Dejmek, J.; Dib, K.; Jonsson, M.; Andersson, T. Wnt-5a and G-Protein Signaling Are Required for Collagen-Induced DDR1 Receptor Activation and Normal Mammary Cell Adhesion. Int. J. Cancer 2003, 103, 344–351. [Google Scholar] [CrossRef]
- Romayor, I.; Badiola, I.; Olaso, E. Inhibition of DDR1 Reduces Invasive Features of Human A375 Melanoma, HT29 Colon Carcinoma and SK-HEP Hepatoma Cells. Cell Adhes. Migr. 2020, 14, 69–81. [Google Scholar] [CrossRef] [Green Version]
- Yan, Z.; Jin, S.; Wei, Z.; Huilian, H.; Zhanhai, Y.; Yue, T.; Juan, L.; Jing, L.; Libo, Y.; Xu, L. Discoidin Domain Receptor 2 Facilitates Prostate Cancer Bone Metastasis via Regulating Parathyroid Hormone-Related Protein. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2014, 1842, 1350–1363. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-L.; Tsai, W.-H.; Ko, Y.-C.; Lai, T.-Y.; Cheng, A.-J.; Shiah, S.-G.; Hsiao, J.-R.; Chang, J.-Y.; Lin, S.-F. Discoidin Domain Receptor-1 (DDR1) Is Involved in Angiolymphatic Invasion in Oral Cancer. Cancers 2020, 12, 841. [Google Scholar] [CrossRef] [Green Version]
- Castro-Sanchez, L.; Soto-Guzman, A.; Navarro-Tito, N.; Martinez-Orozco, R.; Salazar, E.P. Native Type IV Collagen Induces Cell Migration through a CD9 and DDR1-Dependent Pathway in MDA-MB-231 Breast Cancer Cells. Eur. J. Cell Biol. 2010, 89, 843–852. [Google Scholar] [CrossRef]
- Chow, C.R.; Ebine, K.; Knab, L.M.; Bentrem, D.J.; Kumar, K.; Munshi, H.G. Cancer Cell Invasion in Three-Dimensional Collagen Is Regulated Differentially by Gα 13 Protein and Discoidin Domain Receptor 1-Par3 Protein Signaling. J. Biol. Chem. 2016, 291, 1605–1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poudel, B.; Lee, Y.-M.; Kim, D.-K. DDR2 Inhibition Reduces Migration and Invasion of Murine Metastatic Melanoma Cells by Suppressing MMP2/9 Expression through ERK/NF- B Pathway. Acta Biochim. Biophys. Sin. 2015, 47, 292–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Lu, W.; Zhang, S.; Zhu, C.; Ren, T.; Zhu, T.; Zhao, H.; Liu, Y.; Su, J. Overexpression of DDR2 Contributes to Cell Invasion and Migration in Head and Neck Squamous Cell Carcinoma. Cancer Biol. Ther. 2014, 15, 612–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grither, W.R.; Longmore, G.D. Inhibition of Tumor-Microenvironment Interaction and Tumor Invasion by Small-Molecule Allosteric Inhibitor of DDR2 Extracellular Domain. Proc. Natl. Acad. Sci. USA 2018, 115, E7786–E7794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, C.; Greengard, P.; Nairn, A.C.; Andersson, T.; Vogel, W.F. Phosphorylation of DARPP-32 Regulates Breast Cancer Cell Migration Downstream of the Receptor Tyrosine Kinase DDR1. Exp. Cell Res. 2006, 312, 4011–4018. [Google Scholar] [CrossRef] [PubMed]
- Yuge, R.; Kitadai, Y.; Takigawa, H.; Naito, T.; Oue, N.; Yasui, W.; Tanaka, S.; Chayama, K. Silencing of Discoidin Domain Receptor-1 (DDR1) Concurrently Inhibits Multiple Steps of Metastasis Cascade in Gastric Cancer. Transl. Oncol. 2018, 11, 575–584. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhi, Z.; Wang, L.; Zhao, Y.; Deng, M.; Liu, Y.; Qin, Y.; Tian, M.; Liu, Y.; Shen, T.; et al. NSD2 Circular RNA Promotes Metastasis of Colorectal Cancer by Targeting MiR-199b-5p-mediated DDR1 and JAG1 Signalling. J. Pathol. 2019, 248, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Larrieu-Lahargue, F.; Welm, A.L.; Bouchecareilh, M.; Alitalo, K.; Li, D.Y.; Bikfalvi, A.; Auguste, P. Blocking Fibroblast Growth Factor Receptor Signaling Inhibits Tumor Growth, Lymphangiogenesis, and Metastasis. PLoS ONE 2012, 7, e39540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Q.; Jiang, Y.; Liu, Q.; Yue, J.; Liu, C.; Zhao, X.; Qiao, Y.; Ji, H.; Chen, J.; Ge, G. Minor Type IV Collagen A5 Chain Promotes Cancer Progression through Discoidin Domain Receptor-1. PLoS Genet. 2015, 11, e1005249. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.; Seo, M.; Choi, J.-S.; Joo, C.-K.; Lee, S.K. MiR-199a/b-5p Inhibits Lymphangiogenesis by Targeting Discoidin Domain Receptor 1 in Corneal Injury. Mol. Cells 2018, 41, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Bu, X.; Zhao, H.; Yu, J.; Wang, Y.; Li, D.; Zhu, C.; Zhu, T.; Ren, T.; Liu, X.; et al. A Host Deficiency of Discoidin Domain Receptor 2 (DDR2) Inhibits Both Tumour Angiogenesis and Metastasis: DDR2 Regulates Endothelial Function and Tumour Angiogenesis. J. Pathol. 2014, 232, 436–448. [Google Scholar] [CrossRef]
- Lee, N.O.; Park, J.-W.; Lee, J.A.; Shim, J.H.; Kong, S.-Y.; Kim, K.T.; Lee, Y.-S. Dual Action of a Selective Cyclooxygenase-2 Inhibitor on Vascular Endothelial Growth Factor Expression in Human Hepatocellular Carcinoma Cells: Novel Involvement of Discoidin Domain Receptor 2. J. Cancer Res. Clin. Oncol. 2012, 138, 73–84. [Google Scholar] [CrossRef]
- Romayor, I.; Badiola, I.; Benedicto, A.; Marquez, J.; Herrero, A.; Arteta, B.; Olaso, E. Silencing of Sinusoidal DDR1 Reduces Murine Liver Metastasis by Colon Carcinoma. Sci. Rep. 2020, 10, 18398. [Google Scholar] [CrossRef] [PubMed]
- Badiola, I.; Olaso, E.; Crende, O.; Friedman, S.L.; Vidal-Vanaclocha, F. Discoidin Domain Receptor 2 Deficiency Predisposes Hepatic Tissue to Colon Carcinoma Metastasis. Gut 2012, 61, 1465–1472. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, J.; Fushida, S.; Harada, S.; Makino, I.; Nakamura, K.; Oyama, K.; Fujita, H.; Ninomiya, I.; Fujimura, T.; Kayahara, M.; et al. Type IV Collagen Levels Are Elevated in the Serum of Patients with Peritoneal Dissemination of Gastric Cancer. Oncol. Lett. 2010, 1, 989–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, G.; Chen, J.; Zhang, G.; Wang, S.; Kawasaki, K.; Zhu, J.; Zhang, Y.; Nagata, K.; Li, Z.; Zhou, B.P.; et al. Hsp47 Promotes Cancer Metastasis by Enhancing Collagen-Dependent Cancer Cell-Platelet Interaction. Proc. Natl. Acad. Sci. USA 2020, 117, 3748–3758. [Google Scholar] [CrossRef] [Green Version]
- Ye, L.; Pu, C.; Tang, J.; Wang, Y.; Wang, C.; Qiu, Z.; Xiang, T.; Zhang, Y.; Peng, W. Transmembrane-4 L-Six Family Member-1 (TM4SF1) Promotes Non-Small Cell Lung Cancer Proliferation, Invasion and Chemo-Resistance through Regulating the DDR1/Akt/ERK-MTOR Axis. Respir. Res. 2019, 20, 106. [Google Scholar] [CrossRef]
- Jia, S.; Agarwal, M.; Yang, J.; Horowitz, J.C.; White, E.S.; Kim, K.K. Discoidin Domain Receptor 2 Signaling Regulates Fibroblast Apoptosis through PDK1/Akt. Am. J. Respir. Cell Mol. Biol. 2018, 59, 295–305. [Google Scholar] [CrossRef]
- Matà, R.; Palladino, C.; Nicolosi, M.L.; Presti, A.R.L.; Malaguarnera, R.; Ragusa, M.; Sciortino, D.; Morrione, A.; Maggiolini, M.; Vella, V.; et al. IGF-I Induces Upregulation of DDR1 Collagen Receptor in Breast Cancer Cells by Suppressing MIR-199a-5p through the PI3K/AKT Pathway. Oncotarget 2016, 7, 7683–7700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vella, V.; Malaguarnera, R.; Nicolosi, M.L.; Palladino, C.; Spoleti, C.; Massimino, M.; Vigneri, P.; Purrello, M.; Ragusa, M.; Morrione, A.; et al. Discoidin Domain Receptor 1 Modulates Insulin Receptor Signaling and Biological Responses in Breast Cancer Cells. Oncotarget 2017, 8, 43248–43270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vella, V.; Nicolosi, M.L.; Cantafio, P.; Massimino, M.; Lappano, R.; Vigneri, P.; Ciuni, R.; Gangemi, P.; Morrione, A.; Malaguarnera, R.; et al. DDR1 Regulates Thyroid Cancer Cell Differentiation via IGF-2/IR-A Autocrine Signaling Loop. Endocr.-Relat. Cancer 2019, 26, 197–214. [Google Scholar] [CrossRef] [PubMed]
- Buraschi, S.; Morcavallo, A.; Neill, T.; Stefanello, M.; Palladino, C.; Xu, S.-Q.; Belfiore, A.; Iozzo, R.V.; Morrione, A. Discoidin Domain Receptor 1 Functionally Interacts with the IGF-I System in Bladder Cancer. Matrix Biol. Plus 2020, 6–7, 100022. [Google Scholar] [CrossRef]
- Avino, S.; Marco, P.D.; Cirillo, F.; Santolla, M.F.; Francesco, E.M.D.; Perri, M.G.; Rigiracciolo, D.; Dolce, V.; Belfiore, A.; Maggiolini, M.; et al. Stimulatory Actions of IGF-I Are Mediated by IGF-IR Cross-Talk with GPER and DDR1 in Mesothelioma and Lung Cancer Cells. Oncotarget 2016, 7, 52710–52728. [Google Scholar] [CrossRef]
- Morcavallo, A.; Gaspari, M.; Pandini, G.; Palummo, A.; Cuda, G.; Larsen, M.R.; Vigneri, R.; Belfiore, A. Research Resource: New and Diverse Substrates for the Insulin Receptor Isoform A Revealed by Quantitative Proteomics After Stimulation With IGF-II or Insulin. Mol. Endocrinol. 2011, 25, 1456–1468. [Google Scholar] [CrossRef] [Green Version]
- Vella, V.; Malaguarnera, R.; Nicolosi, M.L.; Morrione, A.; Belfiore, A. Insulin/IGF Signaling and Discoidin Domain Receptors: An Emerging Functional Connection. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 118522. [Google Scholar] [CrossRef]
- Day, E.; Waters, B.; Spiegel, K.; Alnadaf, T.; Manley, P.W.; Buchdunger, E.; Walker, C.; Jarai, G. Inhibition of Collagen-Induced Discoidin Domain Receptor 1 and 2 Activation by Imatinib, Nilotinib and Dasatinib. Eur. J. Pharmacol. 2008, 599, 44–53. [Google Scholar] [CrossRef]
- Montenegro, R.C.; Howarth, A.; Ceroni, A.; Fedele, V.; Farran, B.; Mesquita, F.P.; Frejno, M.; Berger, B.-T.; Heinzlmeir, S.; Sailem, H.Z.; et al. Identification of Molecular Targets for the Targeted Treatment of Gastric Cancer Using Dasatinib. Oncotarget 2020, 11, 535–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- von Maessenhausen, A.; Sanders, C.; Braegelmann, J.; Konantz, M.; Queisser, A.; Vogel, W.; Kristiansen, G.; Duensing, S.; Schroeck, A.; Bootz, F.; et al. Targeting DDR2 in Head and Neck Squamous Cell Carcinoma with Dasatinib. Int. J. Cancer 2016, 139, 2359–2369. [Google Scholar] [CrossRef] [Green Version]
- Gajdosik, Z. Sitravatinib Multitargeted Tyrosine Kinase Inhibitor Treatment of Solid Tumors. Drug Future 2018, 43, 181–187. [Google Scholar] [CrossRef]
- Aguilera, K.Y.; Huang, H.; Du, W.; Hagopian, M.M.; Wang, Z.; Hinz, S.; Hwang, T.H.; Wang, H.; Fleming, J.B.; Castrillon, D.H.; et al. Inhibition of Discoidin Domain Receptor 1 Reduces Collagen-Mediated Tumorigenicity in Pancreatic Ductal Adenocarcinoma. Mol. Cancer Ther. 2017, 16, 2473–2485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nokin, M.-J.; Darbo, E.; Travert, C.; Drogat, B.; Lacouture, A.; San José, S.; Cabrera, N.; Turcq, B.; Prouzet-Mauleon, V.; Falcone, M.; et al. Inhibition of DDR1 Enhances in Vivo Chemosensitivity in KRAS-Mutant Lung Adenocarcinoma. JCI Insight 2020, 5, e137869. [Google Scholar] [CrossRef] [PubMed]
- Elkamhawy, A.; Park, J.-E.; Cho, N.-C.; Sim, T.; Pae, A.N.; Roh, E.J. Discovery of a Broad Spectrum Antiproliferative Agent with Selectivity for DDR1 Kinase: Cell Line-Based Assay, Kinase Panel, Molecular Docking, and Toxicity Studies. J. Enzyme. Inhib. Med. Chem. 2016, 31, 158–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mo, C.; Zhang, Z.; Li, Y.; Huang, M.; Zou, J.; Luo, J.; Tu, Z.-C.; Xu, Y.; Ren, X.; Ding, K.; et al. Design and Optimization of 3′-(Imidazo[1,2-a]Pyrazin-3-Yl)-[1,1′-Biphenyl]-3-Carboxamides as Selective DDR1 Inhibitors. ACS Med. Chem. Lett. 2020, 11, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-G.; Tan, L.; Weisberg, E.L.; Liu, F.; Canning, P.; Choi, H.G.; Ezell, S.A.; Wu, H.; Zhao, Z.; Wang, J.; et al. Discovery of a Potent and Selective DDR1 Receptor Tyrosine Kinase Inhibitor. ACS Chem. Biol. 2013, 8, 2145–2150. [Google Scholar] [CrossRef] [Green Version]
- Richter, H.; Satz, A.L.; Bedoucha, M.; Buettelmann, B.; Petersen, A.C.; Harmeier, A.; Hermosilla, R.; Hochstrasser, R.; Burger, D.; Gsell, B.; et al. DNA-Encoded Library-Derived DDR1 Inhibitor Prevents Fibrosis and Renal Function Loss in a Genetic Mouse Model of Alport Syndrome. ACS Chem. Biol. 2019, 14, 37–49. [Google Scholar] [CrossRef]
- Siddiqui, K.; Kim, G.W.; Lee, D.H.; Shin, H.R.; Yang, E.G.; Lee, N.T.; Yang, B.-S. Actinomycin D Identified as an Inhibitor of Discoidin Domain Receptor 2 Interaction with Collagen through an Insect Cell Based Screening of a Drug Compound Library. Biol. Pharm. Bull. 2009, 32, 136–141. [Google Scholar] [CrossRef] [Green Version]
- Terai, H.; Tan, L.; Beauchamp, E.M.; Hatcher, J.M.; Liu, Q.; Meyerson, M.; Gray, N.S.; Hammerman, P.S. Characterization of DDR2 Inhibitors for the Treatment of DDR2 Mutated Nonsmall Cell Lung Cancer. ACS Chem. Biol. 2015, 10. [Google Scholar] [CrossRef]
- Baltes, F.; Caspers, J.; Henze, S.; Schlesinger, M.; Bendas, G. Targeting Discoidin Domain Receptor 1 (DDR1) Signaling and Its Crosstalk with Β1-Integrin Emerges as a Key Factor for Breast Cancer Chemosensitization upon Collagen Type 1 Binding. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef]
- Vehlow, A.; Cordes, N. DDR1 (Discoidin Domain Receptor Tyrosine Kinase 1) Drives Glioblastoma Therapy Resistance by Modulating Autophagy. Autophagy 2019, 15, 1487–1488. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Zhao, F.; Hui, L.; Li, X.; Zhang, D.; Lin, W.; Chen, Z.; Ning, Y. Suppressing MiR-199a-3p by Promoter Methylation Contributes to Tumor Aggressiveness and Cisplatin Resistance of Ovarian Cancer through Promoting DDR1 Expression. J. Ovarian Res. 2017, 10, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, S.L.; Tan, M.L.; Hollows, R.J.; Robinson, M.; Ibrahim, M.; Margielewska, S.; Parkinson, E.K.; Ramanathan, A.; Zain, R.B.; Mehanna, H.; et al. Collagen Induces a More Proliferative, Migratory and Chemoresistant Phenotype in Head and Neck Cancer via DDR1. Cancers 2019, 11, 1766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chappell, W.H.; Candido, S.; Abrams, S.L.; Akula, S.M.; Steelman, L.S.; Martelli, A.M.; Ratti, S.; Cocco, L.; Cervello, M.; Montalto, G.; et al. Influences of TP53 and the Anti-Aging DDR1 Receptor in Controlling Raf/MEK/ERK and PI3K/Akt Expression and Chemotherapeutic Drug Sensitivity in Prostate Cancer Cell Lines. Aging 2020, 12, 10194–10210. [Google Scholar] [CrossRef]
- Tu, M.M.; Lee, F.Y.F.; Jones, R.T.; Kimball, A.K.; Saravia, E.; Graziano, R.F.; Coleman, B.; Menard, K.; Yan, J.; Michaud, E.; et al. Targeting DDR2 Enhances Tumor Response to Anti–PD-1 Immunotherapy. Sci. Adv. 2019, 5, eaav2437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beauchamp, E.M.; Woods, B.A.; Dulak, A.M.; Tan, L.; Xu, C.; Gray, N.S.; Bass, A.J.; Wong, K.-K.; Meyerson, M.; Hammerman, P.S. Acquired Resistance to Dasatinib in Lung Cancer Cell Lines Conferred by DDR2 Gatekeeper Mutation and NF1 Loss. Mol. Cancer Ther. 2014, 13, 475–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.; Svoboda, R.A.; Lazenby, A.J.; Saowapa, J.; Chaika, N.; Ding, K.; Wheelock, M.J.; Johnson, K.R. Up-Regulation of N-Cadherin by Collagen I-Activated Discoidin Domain Receptor 1 in Pancreatic Cancer Requires the Adaptor Molecule Shc1. J. Biol. Chem. 2016, 291, 23208–23223. [Google Scholar] [CrossRef] [Green Version]
- Ram, R.; Lorente, G.; Nikolich, K.; Urfer, R.; Foehr, E.; Nagavarapu, U. Discoidin Domain Receptor-1a (DDR1a) Promotes Glioma Cell Invasion and Adhesion in Association with Matrix Metalloproteinase-2. J. Neurooncol. 2006, 76, 239–248. [Google Scholar] [CrossRef]
- Yang, S.H.; Baek, H.A.; Lee, H.J.; Park, H.S.; Jang, K.Y.; Kang, M.J.; Lee, D.G.; Lee, Y.C.; Moon, W.S.; Chung, M.J. Discoidin Domain Receptor 1 Is Associated with Poor Prognosis of Non-Small Cell Lung Carcinomas. Oncol. Rep. 2010, 24, 311–319. [Google Scholar] [CrossRef] [Green Version]



| Receptor | MMP Activation/Invadosome Formation | Cancers |
|---|---|---|
| DDR1 | Increase | -Lung carcinoma [82] -Pituitary adenoma [83] -Colorectal carcinoma [84] -Renal carcinoma [85] -Pancreatic [86] -Liver [87] -Breast carcinoma [24] |
| Decrease | ||
| DDR2 | Increase | -Ovarian [32] -Melanoma [62]-Hepatocellular carcinoma [89] -Breast [90] |
| Decrease |
| Receptor | EMT | Cancers |
|---|---|---|
| DDR1 | Increase | -Prostate [101] -Gastric [102,103] -Hepatocellular carcinoma [35] -Colorectal [84] -Renal carcinoma [85] |
| Decrease | -Breast carcinoma [31,104,105] -Ovarian [45,104] -Liver (female, [104]) | |
| DDR2 | Increase | -Breast [30,90,98] -Ovarian [32] -Papillary thyroid carcinoma [107] -Hepatocellular carcinoma [89] -Gastric [67] |
| Decrease |
| Receptor | Migration/Invasion | Cancers |
|---|---|---|
| DDR1 | Increase | -Squamous cell carcinoma [25,112] -Hepatocellular carcinoma [35] -Pancreatic [86] -Breast [113] -Renal carcinoma [85] -Gastric [103] -Prostate [53] -Colorectal [120] -Epidermoid carcinoma [48] |
| Decrease | -Breast [118] -Pancreatic [114] | |
| DDR2 | Increase | -Gastric [64] -Hepatocellular carcinoma [65] -Melanoma [115] -Squamous cell carcinoma [63,116] -Breast [99,117] -Ovarian [32] |
| Decrease |
| Inhibitor | Targeted Kinases | In Vitro Assay (Cancer) | Preclinical (Cancer) | Clinical (DDR Targeted) |
|---|---|---|---|---|
| Imatinib | BCR-ABL, DDR1, DDR2 | |||
| Nilotinib | ABL1, Kit, PDGFRA, PDGFRB, CSF1R, DDR1, DDR2 | DDR1: -Breast (decreased apoptosis and DDR1 phosphorylation) -Colon (increased proliferation, decreased DDR1 phosphorylation). | -NCT02029001: Malignant solid neoplasms. Mutated DDR1, mutated DDR2. Phase 2 (recruiting). | |
| Dasatinib | BCR-ABL, SRC kinase family, DDR1, DDR2 | DDR1: -Lung (decreased proliferation). -Gastric (decreased proliferation, migration, invasion). DDR2: -Head and neck (decreased proliferation, migration, invasion) | DDR2: -Head and neck (zebrafish): decreased migration | -NCT01491633: Squamous cell lung cancer. Mutated DDR2. Phase 2 (not evaluable, toxicity). -NCT04439305: lymphoma, myeloma, solid neoplasm. Mutated DDR2. Phase 2: withdrawn. -NCT01514864: NSCLC. Mutated DDR2. Phase 2: not completed, disease progression. |
| Sitravatinib (MGCD156) | MET, AXL, VEGFR1, VEGFR2, VEGFR3, PDGFRA, PDGFRB, TRKA, TRKB, DDR2 | -NCT02219711: NSCLC, renal cell carcinoma. Mutated DDR2. Phase 1 (recruiting). |
| Inhibitor | Specificity | In Vitro Assay (Cancer) | Preclinical (Cancer) |
|---|---|---|---|
| 7rh | DDR1 > DDR2 | -Pancreatic (decreased DDR1 phosphorylation, signaling, colony formation, migration) -Gastric (decreased proliferation) -Lung (increased apoptosis) | -Pancreatic (mice): decreased tumor development and proliferation, increased apoptosis and survival (in association with chemotherapy). -Gastric (mice): decreased peritoneal tumor nodules. -Lung (mice): In association with chemotherapy, decreased tumor development and increased apoptosis |
| 7rj | DDR1 > DDR2 | ||
| KST9046 | DDR1 > ARAF, FLT3, LIMK1, LMK2 | -Decreased proliferation (leukemia, lung, colon, brain, melanoma, renal, ovarian, prostate and breast) and DDR1 phosphorylation | |
| 8v | DDR1 > DDR2, BCL-ABL, c-KIT | -Lung: decreased colony formation, proliferation, migration, invasion and DDR1 phosphorylation | |
| DDR1-IN1 | DDR1 > DDR2 | -Decreased DDR1 phosphorylation, cell proliferation (colon, breast, lung, uterus, liver). -Prostate: decreased proliferation, cell viability, colony formation, EMT and migration. Increased apoptosis. | |
| DDR1-IN2 | DDR1 > DDR2 | Decreased DDR1 phosphorylation, cell proliferation (colon, breast, lung, uterus, liver) | |
| 2.45 | DDR1 > DDR2 | Decreased DDR1 phosphorylation | |
| Compound 1 | DDR2 | Decreased DDR2 phosphorylation and lung cell proliferation. | |
| WRG-28 | DDR2 | Breast: decreased migration and invasion. | Decreased breast metastasis to lung |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Majo, S.; Auguste, P. The Yin and Yang of Discoidin Domain Receptors (DDRs): Implications in Tumor Growth and Metastasis Development. Cancers 2021, 13, 1725. https://doi.org/10.3390/cancers13071725
Majo S, Auguste P. The Yin and Yang of Discoidin Domain Receptors (DDRs): Implications in Tumor Growth and Metastasis Development. Cancers. 2021; 13(7):1725. https://doi.org/10.3390/cancers13071725
Chicago/Turabian StyleMajo, Sandra, and Patrick Auguste. 2021. "The Yin and Yang of Discoidin Domain Receptors (DDRs): Implications in Tumor Growth and Metastasis Development" Cancers 13, no. 7: 1725. https://doi.org/10.3390/cancers13071725