
IGF2BP3 Associates with Proliferative Phenotype and Prognostic Features in B-Cell Acute Lymphoblastic Leukemia

 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patient Cohort for Tissue Microarray and Immunohistochemistry
2.2. Image Analysis
2.3. Microarray and RNA-Sequencing Data Sets
2.4. Statistical Analysis
3. Results
3.1. IGF2BP3 Protein Is Widely Expressed in Pediatric B-ALL
3.2. Expression of Ki-67 Is Associated with High IGF2BP3 Protein Expression
3.3. Expression of IGF2BP3 in Hematological Malignancies and B-ALL
3.4. Proliferative “Metagene” Signature in B-ALL
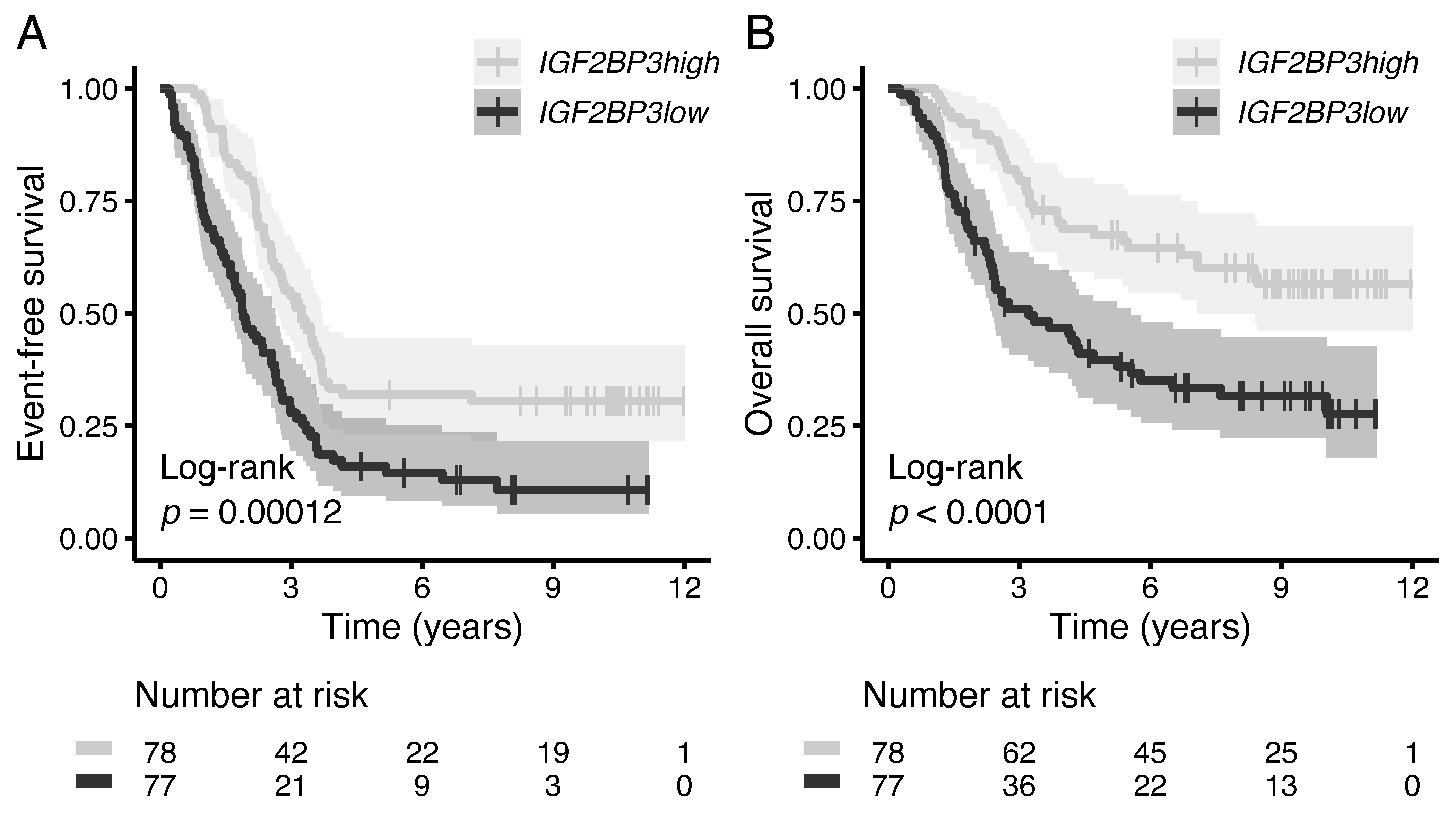
3.5. High IGF2BP3 mRNA Associates with Favorable Survival in High-Risk B-ALL
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Inaba, H.; Greaves, M.; Mullighan, C.G. Acute lymphoblastic leukaemia. Lancet 2013, 381, 1943–1955. [Google Scholar] [CrossRef]
- Hunger, S.P.; Mullighan, C.G. Acute Lymphoblastic Leukemia in Children. N. Engl. J. Med. 2015, 373, 1541–1552. [Google Scholar] [CrossRef] [PubMed]
- Malard, F.; Mohty, M. Acute lymphoblastic leukaemia. Lancet 2020, 395, 1146–1162. [Google Scholar] [CrossRef]
- Mueller-Pillasch, F.; Pohl, B.; Wilda, M.; Lacher, U.; Beil, M.; Wallrapp, C.; Hameister, H.; Knöchel, W.; Adler, G.; Gress, T.M. Expression of the highly conserved RNA binding protein KOC in embryogenesis. Mech. Dev. 1999, 88, 95–99. [Google Scholar] [CrossRef]
- Monk, D.; Bentley, L.; Beechey, C.; Hitchins, M.; Peters, J.; Preece, M.A.; Stanier, P.; Moore, G.E. Characterisation of the growth regulating gene IMP3, a candidate for Silver-Russell syndrome. J. Med. Genet. 2002, 39, 575–581. [Google Scholar] [CrossRef]
- Nielsen, J.; Christiansen, J.; Lykke-Andersen, J.; Johnsen, A.H.; Wewer, U.M.; Nielsen, F.C. A Family of Insulin-Like Growth Factor II mRNA-Binding Proteins Represses Translation in Late Development. Mol. Cell. Biol. 1999, 19, 1262–1270. [Google Scholar] [CrossRef] [PubMed]
- Mancarella, C.; Scotlandi, K. IGF2BP3 From Physiology to Cancer: Novel Discoveries, Unsolved Issues, and Future Perspectives. Front. Cell Dev. Biol. 2020, 7, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Burdelski, C.; Jakani-Karimi, N.; Jacobsen, F.; Möller-Koop, C.; Minner, S.; Simon, R.; Sauter, G.; Steurer, S.; Clauditz, T.S.; Wilczak, W. IMP3 overexpression occurs in various important cancer types and is linked to aggressive tumor features: A tissue microarray study on 8,877 human cancers and normal tissues. Oncol. Rep. 2018, 39, 3–12. [Google Scholar] [CrossRef]
- Chen, L.; Xie, Y.; Li, X.; Gu, L.; Gao, Y.; Tang, L.; Chen, J.; Zhang, X. Prognostic value of high IMP3 expression in solid tumors: A meta-analysis. Onco. Targets. Ther. 2017, 10, 2849–2863. [Google Scholar] [CrossRef] [PubMed]
- King, R.L.; Pasha, T.; Roullet, M.R.; Zhang, P.J.; Bagg, A. IMP-3 is differentially expressed in normal and neoplastic lymphoid tissue. Hum. Pathol. 2009, 40, 1699–1705. [Google Scholar] [CrossRef] [PubMed]
- Findeis-Hosey, J.J.; Xu, H. The use of insulin like-growth factor II messenger RNA binding protein-3 in diagnostic pathology. Hum. Pathol. 2011, 42, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Wei, Q.; Ge, J.; Jian, W.; Liu, J.; Zhong, L.; Fu, B.; Zhao, T. IMP3 as a supplemental diagnostic marker for Hodgkin lymphoma. Hum. Pathol. 2013, 44, 2167–2172. [Google Scholar] [CrossRef] [PubMed]
- Sennekamp, J.; Seelig, H.P. Anti-cytoplasmic autoantibodies in Hodgkin’s lymphoma. Clin. Lab. 2016, 62, 1579–1584. [Google Scholar] [CrossRef] [PubMed]
- Masoud, R.; Ibrahiem, A.; Tantawy, D.; Eldosoky, I. The complementary role of insulin-like growth factor II mRNA-binding protein 3 (IMP3) in diagnosis of Hodgkin’s lymphoma. Ann. Diagn. Pathol. 2019, 42, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, E.M.; Be, S.; Navarro, A.; Trapp, V.; Campo, E.; Ott, G.; Rosenwald, A. Increased tumor cell proliferation in mantle cell lymphoma is associated with elevated insulin-like growth factor 2 mRNA-binding protein 3 expression. Mod. Pathol. 2012, 25, 1227–1235. [Google Scholar] [CrossRef] [PubMed]
- Liao, B.; Hu, Y.; Herrick, D.J.; Brewer, G. The RNA-binding protein IMP-3 is a translational activator of insulin-like growth factor II leader-3 mRNA during proliferation of human K562 leukemia cells. J. Biol. Chem. 2005, 280, 18517–18524. [Google Scholar] [CrossRef] [PubMed]
- Liao, B.; Hu, Y.; Brewer, G. RNA-binding protein insulin-like growth factor mRNA-binding protein 3 (IMP-3) promotes cell survival via insulin-like growth factor II signaling after ionizing radiation. J. Biol. Chem. 2011, 286, 31145–31152. [Google Scholar] [CrossRef] [PubMed]
- Stoskus, M.; Gineikiene, E.; Valceckiene, V.; Valatkaite, B.; Pileckyte, R.; Griskevicius, L. Identification of characteristic IGF2BP expression patterns in distinct B-ALL entities. Blood Cells Mol. Dis. 2011, 46, 321–326. [Google Scholar] [CrossRef]
- Natkunam, Y.; Vainer, G.; Chen, J.; Zhao, S.; Marinelli, R.J.; Hammer, A.S.; Hamilton-Dutoit, S.; Pikarsky, E.; Amir, G.; Levy, R.; et al. Expression of the RNA-binding protein VICKZ in normal hematopoietic tissues and neoplasms. Haematologica 2007, 92, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Palanichamy, J.K.; Tran, T.M.; Howard, J.M.; Contreras, J.R.; Fernando, T.R.; Sterne-Weiler, T.; Katzman, S.; Toloue, M.; Yan, W.; Basso, G.; et al. RNA-binding protein IGF2BP3 targeting of oncogenic transcripts promotes hematopoietic progenitor proliferation. J. Clin. Investig. 2016, 126, 1495–1511. [Google Scholar] [CrossRef] [PubMed]
- Grönroos, T.; Mäkinen, A.; Laukkanen, S.; Mehtonen, J.; Nikkilä, A.; Oksa, L.; Rounioja, S.; Marincevic-Zuniga, Y.; Nordlund, J.; Pohjolainen, V.; et al. Clinicopathological features and prognostic value of SOX11 in childhood acute lymphoblastic leukemia. Sci. Rep. 2020, 10, 2043. [Google Scholar] [CrossRef] [PubMed]
- Mäkinen, A.; Nikkilä, A.; Mehtonen, J.; Teppo, S.; Oksa, L.; Nordlund, J.; Rounioja, S.; Pohjolainen, V.; Laukkanen, S.; Heinäniemi, M.; et al. Expression of BCL6 in paediatric B-cell acute lymphoblastic leukaemia and association with prognosis. Pathology 2021, in press. [Google Scholar]
- Bankhead, P.; Loughrey, M.B.; Fernández, J.A.; Dombrowski, Y.; McArt, D.G.; Dunne, P.D.; McQuaid, S.; Gray, R.T.; Murray, L.J.; Coleman, H.G.; et al. QuPath: Open source software for digital pathology image analysis. Sci. Rep. 2017, 7, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Pölönen, P.; Mehtonen, J.; Lin, J.; Liuksiala, T.; Häyrynen, S.; Teppo, S.; Mäkinen, A.; Kumar, A.; Malani, D.; Pohjolainen, V.; et al. HEMap: An interactive online resource for characterizing molecular phenotypes across hematologic malignancies. Cancer Res. 2019, 79, 2466–2479. [Google Scholar] [CrossRef] [PubMed]
- Mehtonen, J.; Pölönen, P.; Häyrynen, S.; Dufva, O.; Lin, J.; Liuksiala, T.; Granberg, K.; Lohi, O.; Hautamäki, V.; Nykter, M.; et al. Data-driven characterization of molecular phenotypes across heterogeneous sample collections. Nucleic Acids Res. 2019, 47, 76. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.; Churchman, M.L.; Roberts, K.G.; Moore, I.; Zhou, X.; Nakitandwe, J.; Hagiwara, K.; Pelletier, S.; Gingras, S.; Berns, H.; et al. PAX5-driven subtypes of B-progenitor acute lymphoblastic leukemia. Nat. Genet. 2019, 51, 296–307. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Easton, J.; Shao, Y.; Maciaszek, J.; Wang, Z.; Wilkinson, M.R.; McCastlain, K.; Edmonson, M.; Pounds, S.B.; Shi, L.; et al. The genomic landscape of pediatric and young adult T-lineage acute lymphoblastic leukemia. Nat. Genet. 2017, 49, 1211–1218. [Google Scholar] [CrossRef] [PubMed]
- Roberts, K.G.; Li, Y.; Payne-Turner, D.; Harvey, R.C.; Yang, Y.-L.; Pei, D.; McCastlain, K.; Ding, L.; Lu, C.; Song, G.; et al. Targetable Kinase-Activating Lesions in Ph-like Acute Lymphoblastic Leukemia. N. Engl. J. Med. 2014, 371, 1005–1015. [Google Scholar] [CrossRef]
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef] [PubMed]
- Swerdlow, S.H.; Campo, E.; Harris, N.L.; Jaffe, E.S.; Pileri, S.A.; Stein, H.; Thiele, J. WHO Classification of Tumours of Haematopoietic and Lymphoid Tissues, 4th ed.; International Agency for Research on Cancer (IARC): Lyon, France, 2017; ISBN 9789283244943.
- Geng, H.; Hurtz, C.; Lenz, K.B.; Chen, Z.; Baumjohann, D.; Thompson, S.; Goloviznina, N.A.; Chen, W.Y.; Huan, J.; LaTocha, D.; et al. Self-Enforcing Feedback Activation between BCL6 and Pre-B Cell Receptor Signaling Defines a Distinct Subtype of Acute Lymphoblastic Leukemia. Cancer Cell 2015, 27, 409–425. [Google Scholar] [CrossRef]
- Whitfield, M.L.; George, L.K.; Grant, G.D.; Perou, C.M. Common markers of proliferation. Nat. Rev. Cancer 2006, 6, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Giuliano, C.J.; Lin, A.; Smith, J.C.; Palladino, A.C.; Sheltzer, J.M. MELK expression correlates with tumor mitotic activity but is not required for cancer growth. eLife 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- Chan, L.N.; Murakami, M.A.; Robinson, M.E.; Caeser, R.; Sadras, T.; Lee, J.; Cosgun, K.N.; Kume, K.; Khairnar, V.; Xiao, G.; et al. Signalling input from divergent pathways subverts B cell transformation. Nature 2020, 583, 845–851. [Google Scholar] [CrossRef] [PubMed]
- Lederer, M.; Bley, N.; Schleifer, C.; Hüttelmaier, S. The role of the oncofetal IGF2 mRNA-binding protein 3 (IGF2BP3) in cancer. Semin. Cancer Biol. 2014, 29, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Tschirdewahn, S.; Panic, A.; Püllen, L.; Harke, N.N.; Hadaschik, B.; Riesz, P.; Horváth, A.; Szalontai, J.; Nyirády, P.; Baba, H.A.; et al. Circulating and tissue IMP3 levels are correlated with poor survival in renal cell carcinoma. Int. J. Cancer 2019, 145, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Mehtonen, J.; Teppo, S.; Lahnalampi, M.; Kokko, A.; Kaukonen, R.; Oksa, L.; Bouvy-Liivrand, M.; Malyukova, A.; Mäkinen, A.; Laukkanen, S.; et al. Single cell characterization of B-lymphoid differentiation and leukemic cell states during chemotherapy in ETV6-RUNX1-positive pediatric leukemia identifies drug-targetable transcription factor activities. Genome Med. 2020, 12, 99. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Parameter | Median (IQR) |
|---|---|
| Age (years) | 4.3 (2.7–9.7) |
| WBC (x 10E9/l) | 6.3 (2.7–29.2) |
| MRD (%), EOI | 0.01 (0.00–0.14) |
| n (%) | |
| CNS disease | 5 (6.0) |
| Total | 83 |
| WHO Subtype | |
| Other | 32 (38.6) |
| BCR-ABL1 | 1 (1.2) |
| KMT2A-re | 4 (4.8) |
| ETV6-RUNX1 | 20 (24.1) |
| Hyperdiploid | 22 (26.5) |
| Hypodiploid | 1 (1.2) |
| TCF3-PBX1 | 3 (3.6) |
| Event-Free Survival | |||||||
|---|---|---|---|---|---|---|---|
| Univariate | Multivariate | ||||||
| HR | 95% CI | p | HR | 95% CI | p | ||
| IGF2BP3 mRNA | ≤median | 1 | 1 | ||||
| >median | 0.49 | 0.34–0.71 | <0.001 | 0.46 | 0.31–0.68 | <0.001 | |
| Age | 1.01 | 0.97–1.05 | 0.69 | 0.97 | 0.93–1.01 | 0.19 | |
| MRD at the EOI | 1.02 | 0.98–1.07 | 0.39 | 1.01 | 0.97–1.06 | 0.58 | |
| Overall survival | |||||||
| Univariate | Multivariate | ||||||
| HR | 95% CI | p | HR | 95% CI | p | ||
| IGF2BP3 mRNA | ≤median | 1 | 1 | ||||
| >median | 0.44 | 0.28–0.68 | <0.001 | 0.5 | 0.31–0.81 | 0.006 | |
| Age | 1.06 | 1.01–1.11 | 0.01 | 1.03 | 0.98–1.08 | 0.24 | |
| WBC | 1 | 1.00–1.00 | 0.58 | 1 | 0.99–1.00 | 0.68 | |
| MRD at the EOI | 1.02 | 0.97–1.08 | 0.47 | 0.99 | 0.94–1.05 | 0.79 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mäkinen, A.; Nikkilä, A.; Haapaniemi, T.; Oksa, L.; Mehtonen, J.; Vänskä, M.; Heinäniemi, M.; Paavonen, T.; Lohi, O. IGF2BP3 Associates with Proliferative Phenotype and Prognostic Features in B-Cell Acute Lymphoblastic Leukemia. Cancers 2021, 13, 1505. https://doi.org/10.3390/cancers13071505
Mäkinen A, Nikkilä A, Haapaniemi T, Oksa L, Mehtonen J, Vänskä M, Heinäniemi M, Paavonen T, Lohi O. IGF2BP3 Associates with Proliferative Phenotype and Prognostic Features in B-Cell Acute Lymphoblastic Leukemia. Cancers. 2021; 13(7):1505. https://doi.org/10.3390/cancers13071505
Chicago/Turabian StyleMäkinen, Artturi, Atte Nikkilä, Teppo Haapaniemi, Laura Oksa, Juha Mehtonen, Matti Vänskä, Merja Heinäniemi, Timo Paavonen, and Olli Lohi. 2021. "IGF2BP3 Associates with Proliferative Phenotype and Prognostic Features in B-Cell Acute Lymphoblastic Leukemia" Cancers 13, no. 7: 1505. https://doi.org/10.3390/cancers13071505