
The Japanese Wild-Derived Inbred Mouse Strain, MSM/Ms in Cancer Research



Abstract
:Simple Summary
Abstract
1. Introduction
2. Radiation-Induced Thymic Lymphomas
2.1. Identification of Tumor Resistance Loci
2.2. Genetic Analysis of the LOH Region
3. Chemically-Induced Skin Tumors
3.1. Identification of Tumor Resistance Loci
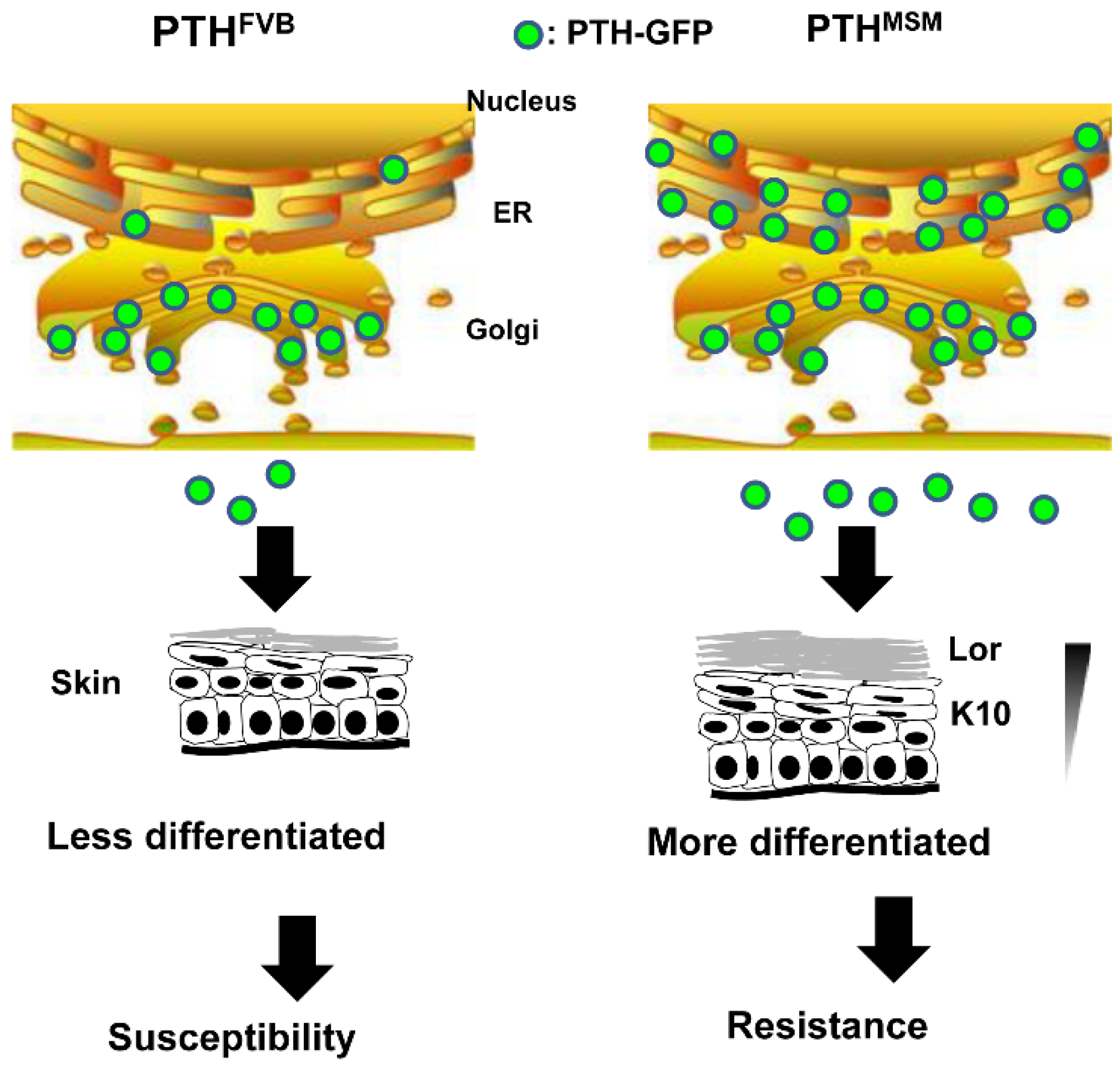
3.2. Tumor Resistance Loci on Chromosome 7
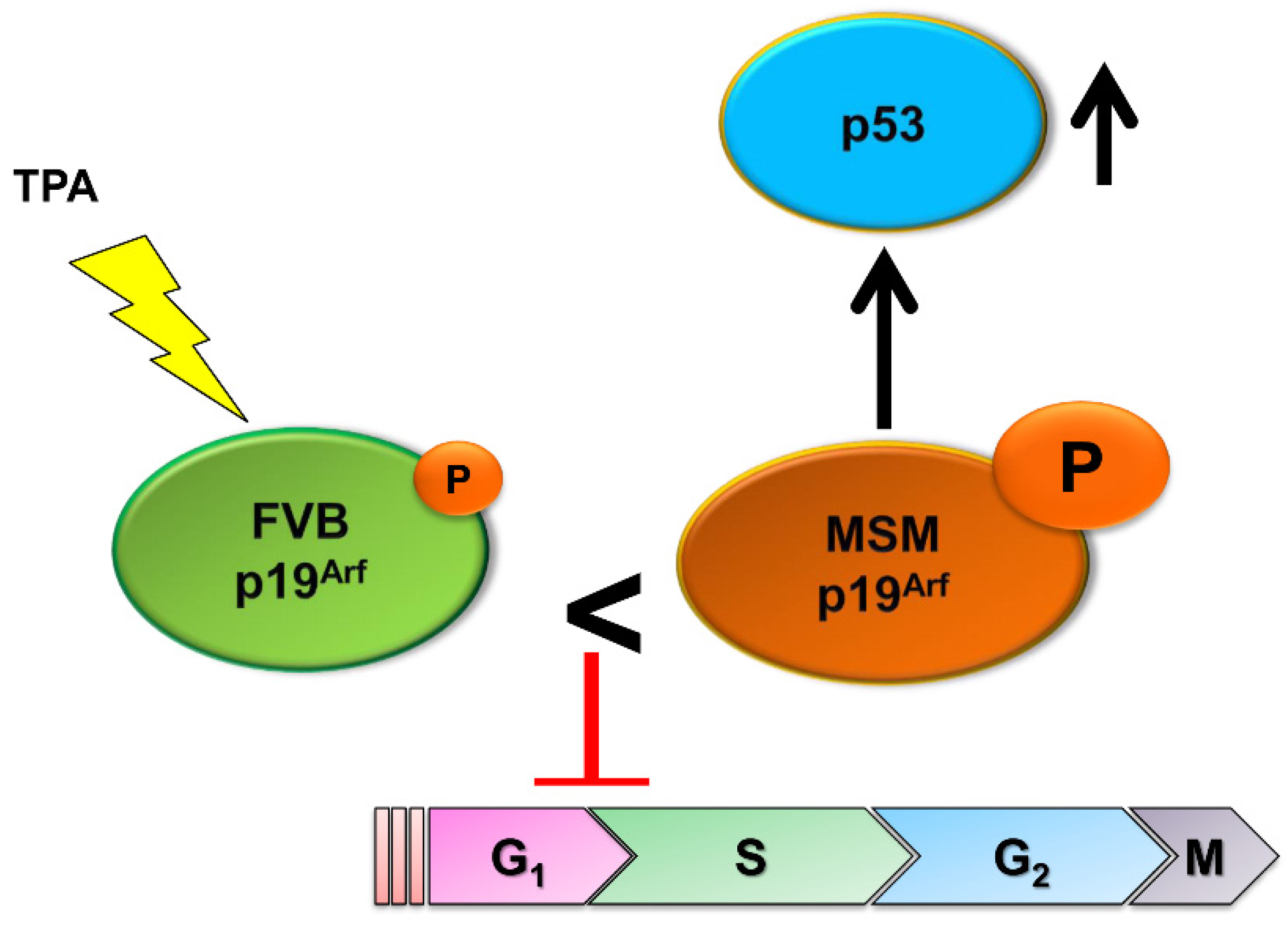
3.3. Tumor Resistance Loci on Chromosome 4
4. Closing Remarks
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stadler, Z.K.; Thom, P.; Robson, M.E.; Weitzel, J.N.; Kauff, N.D.; Hurley, K.E.; Devlin, V.; Gold, B.; Klein, R.J.; Offit, K. Genome-wide association studies of cancer. J. Clin. Oncol. 2010, 28, 4255–4267. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Losada, J.; Castellanos-Martín, A.; Mao, J.H. Cancer evolution and individual susceptibility. Integr. Biol. 2011, 3, 316–328. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, W.F.; Lander, E.S.; Smith, J.S.; Moser, A.R.; Gould, K.A.; Luongo, C.; Borenstein, N.; Dove, W. Genetic identification of Mom-1, a major modifier locus affecting Min-induced intestinal neoplasia in the mouse. Cell 1993, 75, 631–639. [Google Scholar] [CrossRef]
- Fijneman, R.J.; de Vries, S.S.; Jansen, R.C.; Demant, P. Complex interactions of new quantitative trait loci, Sluc1, Sluc2, Sluc3, and Sluc4, that influence the susceptibility to lung cancer in the mouse. Nat. Genet. 1996, 14, 465–467. [Google Scholar] [CrossRef] [PubMed]
- Ruivenkamp, C.A.L.; van Wezel, T.; Zanon, C.; Stassen, A.P.M.; Vlcek, C.; Csikós, T.; Klous, A.M.; Tripodis, N.; Perrakis, A.; Boerrigter, L.; et al. Ptprj is a candidate for the mouse colon-cancer susceptibility locus Scc1 and is frequently deleted in human cancers. Nat. genet. 2002, 31, 295–300. [Google Scholar] [CrossRef]
- Demant, P. Cancer susceptibility in the mouse: Genetics, biology and implications for human cancer. Nat. Rev. Genet. 2003, 4, 721–734. [Google Scholar] [CrossRef]
- Dragani, T.A. 10 years of mouse cancer modifier loci: Human relevance. Cancer Res. 2003, 63, 3011–3018. [Google Scholar]
- Mao, J.-H.; Balmain, A. Genomic approaches to identification of tumour-susceptibility genes using mouse models. Curr. Opin. Genet. Dev. 2003, 13, 14–19. [Google Scholar] [CrossRef]
- Nagase, H.; Bryson, S.; Cordell, H.; Kemp, C.J.; Fee, F.; Balmain, A. Distinct genetic loci control development of benign and malignant skin tumours in mice. Nat. Genet. 1995, 10, 424–429. [Google Scholar] [CrossRef]
- To, M.D.; Perez-Losada, J.; Mao, J.-H.; Hsu, J.; Jacks, T.; Balmain, A. A functional switch from lung cancer resistance to susceptibility at the Pas1 locus in Kras2LA2 mice. Nat. Genet. 2006, 38, 926–930. [Google Scholar] [CrossRef] [Green Version]
- Santos, J.; Montagutelli, X.; Acevedo, A.; López, P.; Vaquero, C.; Fernández, M.; Arnau, M.-R.; Szatanik, M.; Salido, E.; Guénet, J.-L.; et al. A new locus for resistance to gamma-radiation-induced thymic lymphoma identified using inter-specific consomic and inter-specific recombinant congenic strains of mice. Oncogene 2002, 21, 6680–6683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ewart-Toland, A.; Briassouli, P.; de Koning, J.P.; Mao, J.-H.; Yuan, J.; Chan, F.; MacCarthy-Morrogh, L.; Ponder, B.A.J.; Nagase, H.; Burn, J.; et al. Identification of Stk6/STK15 as a candidate low-penetrance tumor-susceptibility gene in mouse and human. Nat. Genet. 2003, 34, 403–412. [Google Scholar] [CrossRef]
- Peissel, B.; Zaffaroni, D.; Zanesi, N.; Zedda, I.; Manenti, G.; Rebessi, S.; Pazzaglia, S.; Doria, G.; Covelli, V.; Dragani, T.A.; et al. Linkage disequilibrium and haplotype mapping of a skin cancer susceptibility locus in outbred mice. Mamm. Genome 2000, 11, 979–981. [Google Scholar] [CrossRef]
- Wakabayashi, Y.; Mao, J.-H.; Brown, K.; Girardi, M.; Balmain, A. Promotion of Hras-induced squamous carcinomas by a polymorphic variant of the Patched gene in FVB mouse. Nature 2007, 445, 761–765. [Google Scholar] [CrossRef] [PubMed]
- Angel, J.M.; Beltrán, L.; Minda, K.; Rupp, T.; DiGiovanni, J. Association of a murine chromosome 9 locus (Psl1) with susceptibility to mouse skin tumor promotion by 12-O-tetradecanoylphorbol-13-acetate. Mol. Carcinog. 1997, 20, 162–167. [Google Scholar] [CrossRef]
- Angel, J.M.; Caballero, M.; DiGiovanni, J. Identification of novel genetic loci contributing to 12-O-tetradecanoylphorbol-13-acetate skin tumor promotion susceptibility in DBA/2 and C57BL/6 mice. Cancer Res. 2003, 63, 2747–2751. [Google Scholar]
- Fujiwara, K.; Igarashi, J.; Irahara, N.; Kimura, M.; Nagase, H. New chemically induced skin tumour susceptibility loci identified in a mouse backcross between FVB and dominant resistant PWK. BMC Genet. 2007, 8, 39. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara, K.; Wie, B.; Elliott, R.; Nagase, H. New outbred colony derived from Mus musculus castaneus to identify skin tumor susceptibility loci. Mol. Carcinog. 2010, 49, 653–661. [Google Scholar] [CrossRef] [Green Version]
- Moriwaki, K.; Shiroishi, T.; Yonekawa, H. Genetics in Wild Mice; Japan Scientific Societies Press: Tokyo, Japan; Karger: Basel, Switzerland, 1994. [Google Scholar]
- Yonekawa, H.; Moriwaki, K.; Gotoh, O.; Miyashita, N.; Matsushima, Y.; Shi, L.M.; Cho, W.S.; Zhen, X.L.; Tagashira, Y. Hybrid origin of Japanese mice ‘‘Mus musculus molossinus’’: Evidence from restriction analysis of mitochondrial DNA. Mol. Biol. Evol. 1988, 5, 63–78. [Google Scholar]
- Takada, T.; Ebata, T.; Noguchi, H.; Keane, T.M.; Adams, D.J.; Narita, T.; Shin-I., T.; Fujisawa, H.; Toyoda, A.; Abe, K.; et al. The ancestor of extant Japanese fancy mice contributed to the mosaic genomes of classical inbred strains. Genome Res. 1988, 23, 1329–1338. [Google Scholar] [CrossRef] [Green Version]
- Moriwaki, K.; Miyashita, N.; Mita, A.; Gotoh, H.; Tsuchiya, K.; Kato, H.; Mekada, K.; Noro, C.; Oota, S.; Yoshiki, A.; et al. Unique inbred strain MSM/Ms established from the Japanese wild mouse. Exp. Anim. 2009, 58, 123–134. [Google Scholar] [CrossRef] [Green Version]
- Kikkawa, Y.; Miura, I.; Takahama, S.; Wakana, S.; Yamazaki, Y.; Moriwaki, K.; Shiroishi, T.; Yonekawa, H. Microsatellite database for MSM/Ms and JF1/Ms, molossinus-derived inbred strains. Mamm. Genome 2001, 12, 750–752. [Google Scholar] [CrossRef] [PubMed]
- Abe, K.; Noguchi, H.; Tagawa, K.; Yuzuriha, M.; Toyoda, A.; Kojima, T.; Ezawa, K.; Saitou, N.; Hattori, M.; Sakaki, Y.; et al. Contribution of Asian mouse subspecies Mus musculus molossinus to genomic constitution of strain C57BL/6J, as defined by BAC-end sequence-SNP analysis. Genome Res. 2004, 14, 2439–2447. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, M.; Ohno, T.; Tsuchiya, T.; Horio, F. Characterization of diabetes-related traits in MSM and JF1 mice on high-fat diet. J. Nutr. Biochem. 2004, 15, 614–621. [Google Scholar] [CrossRef]
- Koide, T.; Moriwaki, K.; Ikeda, K.; Niki, H.; Shiroishi, T. Multiphenotype behavioral characterization of inbred strains derived from wild stocks of Mus musculus. Mamm. Genome 2000, 11, 664–670. [Google Scholar] [CrossRef]
- Nemoto, M.; Morita, Y.; Mishima, Y.; Takahashi, S.; Nomura, T.; Ushiki, T.; Shiroishi, T.; Kikkawa, Y.; Yonekawa, H.; Kominami, R. Ahl3, a third locus on mouse chromosome 17 affecting age-related hearing loss. Biochem. Biophys. Res. Commun. 2004, 324, 1283–1288. [Google Scholar] [CrossRef]
- Miyasaka, Y.; Suzuki, S.; Ohshiba, Y.; Watanabe, K.; Sagara, Y.; Yasuda, S.-P.; Matsuoka, K.; Shitara, H.; Yonekawa, H.; Kominami, R.; et al. Compound heterozygosity of the functionally null Cdh23(v-ngt) and hypomorphic Cdh23(ahl) alleles leads to early-onset progressive hearing loss in mice. Exp. Anim. 2013, 62, 333–346. [Google Scholar] [CrossRef] [Green Version]
- Nakanishi, M.; Tazawa, H.; Tsuchiya, N.; Sugimura, T.; Tanaka, T.; Nakagama, H. Mouse strain differences in inflammatory responses of colonic mucosa induced by dextran sulfate sodium cause differential susceptibility to PhIP-induced large bowel carcinogenesis. Cancer Sci. 2007, 98, 1157–1163. [Google Scholar] [CrossRef]
- Okamoto, M.; Yonekawa, H. Intestinal tumorigenesis in Min mice is enhanced by X-irradiation in an age-dependent manner. J. Radiat. Res. 2005, 46, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Pataer, A.; Kamoto, T.; Lu, L.M.; Yamada, Y.; Hiai, H. Two dominant host resistance genes to pre-B lymphoma in wild-derived inbred mouse strain MSM/Ms. Cancer Res. 1996, 56, 3716–3720. [Google Scholar]
- Okumoto, M.; Mori, N.; Miyashita, N.; Moriwaki, K.; Imai, S.; Haga, S.; Hiroishi, S.; Takamori, Y.; Esaki, K. Radiation-induced lymphomas in MSM, (BALB/cHeA x MSM) F1 and (BALB/cHeA x STS/A) F1 hybrid mice. Exp. Anim. 1995, 44, 43–48. [Google Scholar] [CrossRef] [Green Version]
- Saito, Y.; Ochiai, Y.; Kodama, Y.; Tamura, Y.; Togashi, T.; Kosugi-Okano, H.; Miyazawa, T.; Wakabayashi, Y.; Hatakeyama, K.; Wakana, S.; et al. Genetic loci controlling susceptibility to gamma-ray-induced thymic lymphoma. Oncogene 2001, 20, 5243–5247. [Google Scholar] [CrossRef] [Green Version]
- Miyasaka, K.; Ohtake, K.; Nomura, K.; Kanda, H.; Kominami, R.; Miyashita, N.; Kitagawa, T. Frequent loss of heterozygosity on chromosome 4 in diethylnitrosamine-induced C3H/MSM mouse hepatocellular carcinomas in culture. Mol. Carcinog. 1995, 13, 37–43. [Google Scholar] [CrossRef]
- Masui, T.; Tezuka, N.; Nakanishi, H.; Inada, K.-I.; Miyashita, N.; Tatematsu, M. Induction of invasive squamous cell carcinomas in the forestomach of (C3H x MSM)F1, MSM, and C3H mice by N-methyl-N-nitrosourea and mutational analysis of the H-ras and p53 genes. Cancer Lett. 1997, 111, 97–104. [Google Scholar] [CrossRef]
- Okumura, K.; Sato, M.; Saito, M.; Miura, I.; Wakana, S.; Mao, J.H.; Miyasaka, Y.; Kominami, R.; Wakabayashi, Y. Independent genetic control of early and late stages of chemically induced skin tumors in a cross of a Japanese wild-derived inbred mouse strain, MSM/Ms. Carcinogenesis 2012, 33, 2260–2268. [Google Scholar] [CrossRef]
- Araki, K.; Takeda, N.; Yoshiki, A.; Obata, Y.; Nakagata, N.; Shiroishi, T.; Moriwaki, K.; Yamamura, K. Establishment of germline-competent embryonic stem cell lines from the MSM/Ms strain. Mamm. Genome 2009, 20, 14–20. [Google Scholar] [CrossRef]
- Hemann, M.T.; Greider, C.W. Wild-derived inbred mouse strains have short telomeres. Nucleic Acids Res. 2000, 28, 4474–4478. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Hathcock, K.S.; Hande, P.; Lansdorp, P.M.; Seldin, M.F.; Hodes, R.J. Telomere length regulation in mice is linked to a novel chromosome locus. Proc. Natl. Acad. Sci. USA 1998, 95, 8648–8653. [Google Scholar] [CrossRef] [Green Version]
- Ullrich, R.L.; Bowles, N.D.; Satterfield, L.C.; Davis, C.M. Strain-dependent susceptibility to radiation-induce mammary cancer is a result of differences in epithelial cell sensitivity to transformation. Radiat. Res. 1996, 146, 353–355. [Google Scholar] [CrossRef]
- Li, Y.R.; Halliwill, K.D.; Adams, C.J.; Iyer, V.; Riva, L.; Mamunur, R.; Jen, K.Y.; Del Rosario, R.; Fredlund, E.; Hirst, G.; et al. Mutational signatures in tumours induced by high and low energy radiation in Trp53 deficient mice. Nat. Commun. 2020, 11, 394. [Google Scholar]
- Tamura, Y.; Maruyama, M.; Mishima, Y.; Fujisawa, H.; Obata, M.; Kodama, Y.; Yoshikai, Y.; Aoyagi, Y.; Niwa, O.; Schaffner, W.; et al. Predisposition to mouse thymic lymphomas in response to ionizing radiation depends on variant alleles encoding metal-responsive transcription factor-1 (Mtf-1). Oncogene 2005, 24, 399–406. [Google Scholar] [CrossRef] [Green Version]
- Kodama, Y.; Yoshikai, Y.; Tamura, Y.; Wakana, S.; Takagi, R.; Niwa, O.; Kominami, R. The D5Mit7 locus on mouse chromosome 5 provides resistance to gamma-ray-induced but not N-methyl-N-nitrosourea-induced thymic lymphomas. Carcinogenesis 2004, 25, 143–148. [Google Scholar] [CrossRef] [Green Version]
- Ochiai, Y.; Tamura, Y.; Saito, Y.; Matsuki, A.; Wakabayashi, Y.; Aizawa, Y.; Niwa, O.; Kominami, R. Mapping of genetic modifiers of thymic lymphoma development in p53-knockout mice. Oncogene 2003, 22, 1098–1102. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, Y.; Kosugi, S.; Shinbo, T.; Chou, D.; Ohashi, M.; Wakabayashi, Y.; Sakai, K.; Okumoto, M.; Mori, N.; Aizawa, S.; et al. Allelic loss analysis of gamma-ray-induced mouse thymic lymphomas: Two candidate tumor suppressor gene loci on chromosomes 12 and 16. Oncogene 1998, 16, 2747–2754. [Google Scholar] [CrossRef] [Green Version]
- Shinbo, T.; Matsuki, A.; Matsumoto, Y.; Kosugi, S.; Takahashi, Y.; Niwa, O.; Kominami, R. Allelic loss mapping and physical delineation of a region harboring a putative thymic lymphoma suppressor gene on mouse chromosome 12. Oncogene 1999, 18, 4131–4136. [Google Scholar] [CrossRef] [Green Version]
- Wakabayashi, Y.; Inoue, J.; Takahashi, Y.; Matsuki, A.; Kosugi-Okano, H.; Shinbo, T.; Mishima, Y.; Niwa, O.; Kominami, R. Homozygous deletions and point mutations of the Rit1/Bcl11b gene in gamma-ray induced mouse thymic lymphomas. Biochem. Biophys. Res. Commun. 2003, 301, 598–603. [Google Scholar] [CrossRef]
- Wakabayashi, Y.; Watanabe, H.; Inoue, J.; Takeda, N.; Sakata, J.; Mishima, Y.; Hitomi, J.; Yamamoto, T.; Utsuyama, M.; Niwa, O.; et al. Bcl11b is required for differentiation and survival of alphabeta T lymphocytes. Nat. Immunol. 2003, 4, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Kamimura, K.; Ohi, H.; Kubota, T.; Okazuka, K.; Yoshikai, Y.; Wakabayashi, Y.; Aoyagi, Y.; Mishima, Y.; Kominami, R. Haploinsufficiency of Bcl11b for suppression of lymphomagenesis and thymocyte development. Biochem. Biophys. Res. Commun. 2007, 355, 538–542. [Google Scholar] [CrossRef]
- Okano, H.; Saito, Y.; Miyazawa, T.; Shinbo, T.; Chou, D.; Kosugi, S.; Takahashi, Y.; Odani, S.; Niwa, O.; Kominami, R. Homozygous deletions and point mutations of the Ikaros gene in gamma-ray-induced mouse thymic lymphomas. Oncogene 1999, 18, 6677–6683. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Wakabayashi, Y.; Okano, H.; Saito, Y.; Miyazawa, T.; Kominami, R. Genetic mapping and allelic loss analysis in mouse thymic lymphomas of Helios and Aiolos belonging to the Ikaros gene family. Jpn. J. Cancer Res. 2001, 92, 36–41. [Google Scholar] [CrossRef]
- Abel, E.L.; Angel, J.M.; Kiguchi, K.; DiGiovanni, J. Multi-stage chemical carcinogenesis in mouse skin: Fundamentals and applications. Nat. Protoc. 2009, 4, 1350–1362. [Google Scholar] [CrossRef] [Green Version]
- Okumura, K.; Saito, M.; Isogai, E.; Miura, I.; Wakana, S.; Kominami, R.; Wakabayashi, Y. Congenic mapping and allele-specific alteration analysis of Stmm1 locus conferring resistance to early-stage chemically induced skin papillomas. PLoS ONE 2014, 9, e97201. [Google Scholar] [CrossRef]
- Okumura, K.; Saito, M.; Yoshizawa, Y.; Munakata, H.; Isogai, E.; Miura, I.; Wakana, S.; Yamaguchi, M.; Shitara, H.; Taya, C.; et al. The parathyroid hormone regulates skin tumour susceptibility in mice. Sci. Rep. 2017, 7, 11208. [Google Scholar] [CrossRef] [PubMed]
- Safer, J.D.; Ray, S.; Holick, M.F. A topical parathyroid hormone/parathyroid hormone-related peptide receptor antagonist stimulates hair growth in mice. Endocrinology 2007, 148, 1167–1170. [Google Scholar] [CrossRef] [Green Version]
- Okumura, K.; Saito, M.; Yoshizawa, Y.; Ito, Y.; Isogai, E.; Araki, K.; Wakabayashi, Y. Pak1 maintains epidermal stem cells by regulating Langerhans cells and is required for skin carcinogenesis. Oncogene 2020, 39, 4756–4769. [Google Scholar] [CrossRef] [PubMed]
- Saito, M.; Okumura, K.; Miura, I.; Wakana, S.; Kominami, R.; Wakabayashi, Y. Identification of Stmm3 locus conferring resistance to late-stage chemically induced skin papillomas on mouse chromosome 4 by congenic mapping and allele-specific alteration analysis. Exp. Anim. 2014, 63, 339–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamijo, T.; Zindy, F.; Roussel, M.F.; Quelle, D.E.; Downing, J.R.; Ashmun, R.A.; Grosveld, G.; Sherr, C.J. Tumor suppression at the mouse INK4a locus mediated by the alternative reading frame product p19ARF. Cell 1997, 91, 649–659. [Google Scholar] [CrossRef] [Green Version]
- Kamijo, T.; Weber, J.D.; Zambetti, G.; Zindy, F.; Roussel, M.F.; Sherr, C.J. Functional and physical interactions of the ARF tumor suppressor with p53 and Mdm2. Proc. Natl. Acad. Sci. USA 1998, 95, 8292–8297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, M.; Okumura, K.; Isogai, E.; Araki, K.; Tanikawa, C.; Matsuda, K.; Kamijo, T.; Kominami, R.; Wakabayashi, Y. A Polymorphic Variant in p19Arf Confers Resistance to Chemically Induced Skin Tumors by Activating the p53 Pathway. J. Investig. Derm. 2019, 139, 1459–1469. [Google Scholar] [CrossRef]
- Takada, T.; Shiroishi, T. Complex Quantitative Traits Cracked by the Mouse Inter-Subspecific Consomic Strains. Exp. Anim. 2012, 61, 375–388. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tumor Induction | Tumor Types | References |
|---|---|---|
| Urethan | Lung tumors | [22] |
| PhIP | Intestinal tumors | [29] |
| Min mice | Intestinal tumors | [30] |
| MuLV | B-cell lymphomas | [31] |
| γ-ray | Thymic lymphomas | [32,33] |
| DEN | Hepatocellular carcinomas | [34] |
| MNU | Forestomach tumors | [35] |
| DMBA/TPA | Skin tumors | [36] |
| Tumor Types | FVB/N | BALB/c | C57BL6/J | MSM/Ms |
|---|---|---|---|---|
| Resistance to radiation-induced lymphomas | ++ | ++ | +++ | +++++ |
| Resistance to chemically- induced skin tumors | + | ++ | +++ | +++++ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okumura, K.; Saito, M.; Isogai, E.; Wakabayashi, Y. The Japanese Wild-Derived Inbred Mouse Strain, MSM/Ms in Cancer Research. Cancers 2021, 13, 1026. https://doi.org/10.3390/cancers13051026
Okumura K, Saito M, Isogai E, Wakabayashi Y. The Japanese Wild-Derived Inbred Mouse Strain, MSM/Ms in Cancer Research. Cancers. 2021; 13(5):1026. https://doi.org/10.3390/cancers13051026
Chicago/Turabian StyleOkumura, Kazuhiro, Megumi Saito, Eriko Isogai, and Yuichi Wakabayashi. 2021. "The Japanese Wild-Derived Inbred Mouse Strain, MSM/Ms in Cancer Research" Cancers 13, no. 5: 1026. https://doi.org/10.3390/cancers13051026